中國產細紋獅子魚的骨學和肌肉學研究*
2022-04-11 06:44:08亢世華金顯仕
漁業科學進展 2022年2期
亢世華 金顯仕,2 王 俊,3
中國產細紋獅子魚的骨學和肌肉學研究*
亢世華1金顯仕1,2王 俊1,3①
(1. 中國水產科學研究院黃海水產研究所 農業農村部海洋漁業可持續發展重點實驗室 山東省漁業資源與生態環境重點實驗室 山東 青島 266071;2. 青島海洋科學與技術試點國家實驗室海洋漁業科學與食物產出過程功能實驗室 山東 青島 266071;3. 青島海洋科學與技術試點國家實驗室海洋生態與環境科學功能實驗室 山東 青島 266237)
本研究解剖了中國產細紋獅子魚()標本,并詳細描述了該物種的骨骼系統、骨骼肌系統及相關神經,包括圍眶骨、腦顱、頜骨、懸器和鰓蓋骨、舌弓、鰓弓、肩帶、腰帶、中軸骨架和奇鰭支鰭骨、尾部骨架的10個部分骨骼,以及頰肌、腦顱與懸器–鰓蓋骨間肌肉、頭部腹面肌肉、鰓弓肌肉、胸鰭肌肉、腹鰭肌肉、奇鰭相關肌肉、尾鰭肌肉的8個部分骨骼肌。闡明了骨學和肌肉學名詞術語的更新和同物異名等情況,并對舌弓展肌和鰓弓背面牽縮肌等部分進行中文命名。講解了腹鰭吸盤的特殊構造及其運動機制,以及頰肌的分化和功能。
骨學;肌肉學;細紋獅子魚;形態學;魚類解剖學
細紋獅子魚()隸屬于輻鰭魚綱(Osteichthyes)、鲉形目(Scorpaeniformes)、杜父魚亞目(Cottoidei)、獅子魚科(Liparidae)、獅子魚屬(),是冷溫性底層海洋魚類,分布于太平洋西北部,在中國分布于渤海、黃海和東海(Nakabo, 2002; 金鑫波, 2006)。20世紀80年代起,細紋獅子魚成為黃海魚類群落的優勢種(Jin, 1996),在生態系統中占據非常重要的地位(張波等, 2011)。國內外對細紋獅子魚的研究主要圍繞漁業資源與生態方面進行(單秀娟等, 2014; Myoung, 2016),而以骨骼和肌肉為主的解剖學研究卻少之又少,僅在獅子魚科比較骨學研究中略有提及(Kido, 1988; Chernova, 2008)。
對魚類骨骼肌肉進行精準描述,能為系統發育、進化、形態功能學、動物醫學等研究奠定基礎。但國內對魚類骨骼和肌肉的研究并不充足,近些年只有幾篇涉及局部骨骼(如腦顱、咽顱、胸鰭等)的論文發表(如陳詠霞等, 2014; 楊太有等, 2014)。書籍教材中的解剖學知識也十分有限,有些術語和翻譯未及時更新,內容不夠清晰詳實。本研究對黃海產細紋獅子魚標本進行解剖,詳細描述細紋獅子魚的骨骼、肌肉及相關神經的形態、位置和構造,并將相關解剖學的中英文術語名詞進行了簡要敘述。
1 材料與方法
實驗解剖采用中國水產科學研究院黃海水產研究所館藏的細紋獅子魚標本:YSFIRII-000287 (TL 185.5 mm, 35°N, 123°E)和YSFIRII-000475 (TL 200.4 mm, 33°N, 124°E)。標本為2018年10月搭載該所“北斗”號科學調查船,于黃海中南部海域采用拖網捕獲的。標本經過福爾馬林溶液固定和酒精保存液浸泡,進行了形態學檢查和測量,確保物種鑒定的準確性。先用阿利新藍和茜素紅對整個標本進行軟骨和硬骨染色,再在體式顯微鏡下解剖、觀察并記錄特征,繪制成骨學和肌肉學圖片,并描述其形態構造,闡述相關結構功能。
中文版骨學(Osteology)術語主要參照孟慶聞等(1994)和水柏年等(2019),肌肉學(Myology)術語參照秉志等(1958)和孟慶聞等(1994)。現行英文版骨學術語主要參照Kido (1988)和Kawai (2017),肌肉學術語主要參照Winterbottom(1974)和Endo等(2017)。舊版英文術語主要參照Tominaga (1968)和秉志等(1958)。神經解剖學(Neuroanatomy)術語主要參照秉志等(1959)和Freihofer (1978)。
2 結果
2.1 骨學
2.1.1 圍眶骨(circumorbital bones)(圖1) 魚頭部每側有4塊眶下骨(infraorbital, IO, 曾用英文名suborbital和infrasphenoid),前3塊位于眼的下方,第4塊位于眼的后方。它們構成的管狀結構內通眶下管(infraorbital canal),管道下方開口為上頜部黏液孔。
第1眶下骨(又名淚骨lachrymal)前內側關節面與腭骨前端、上頜骨頭部后側相接觸,內側有一個關節面與側篩骨的側上關節頭相接觸。第2眶下骨緊密連接第1和第3眶下骨。第3眶下骨前部為管狀,后部為細長棒狀,稱為眶下骨架(suborbital stay, SS),橫跨魚的頰部,末端接觸前鰓蓋骨中后部。第4眶下骨為長管狀,體積最小,游離于其他骨頭。

圖1 圍眶骨外側圖
IO 1~4:第1~4眶下骨;SS:眶下骨架
IO 1~4: First to fourth infraorbitals; SS:Suborbital stay
2.1.2 腦顱(neurocranium)(圖2) 腦顱區骨骼包括左右成對的側篩骨、額骨、頂骨、蝶耳骨、翼蝶骨、前耳骨、翼耳骨、上耳骨、后耳骨和側枕骨,以及不成對的前犁骨、中篩骨、上枕骨、基枕骨和副蝶骨。無第6眶下骨(又名膜蝶耳骨dermosphenotic),基蝶骨(basisphenoid)和鼻骨在此描述。

圖2 腦顱部的背面圖(上)、側面圖(中)和腹面圖(下)
BO:基枕骨;E:中篩骨;EC:中篩骨軟骨;EO:側枕骨;EPO:上耳骨;FON:嗅神經孔;FRO:額骨;IC:后耳骨;LET:側篩骨;NA:鼻骨;PA:頂骨;PAS:副蝶骨;PRO:前耳骨;PTO:翼耳骨;PTS:翼蝶骨;PV:前犁骨;SO:上枕骨;SPO:蝶耳骨
BO: Basioccipital; E: Ethmoid; EC: Ethmoid cartilage; EO: Exoccipital; EPO: Epiotic; FON:Foramen for olfactory nerve; FRO: Frontal; IC: Intercalar; LET: Lateral ethmoid; NA: Nasal; PA: Parietal; PAS: Parasphenoid; PRO: Prootic; PTO: Pterotic; PTS: Pterosphenoid; PV: Prevomer; SO: Supraoccipital; SPO: Sphenotic
鼻骨(nasal, NA)近長方形片狀,具不完全管狀結構,位于頭部最前端,眶上管(supraorbital canal)由額骨通向鼻骨。
前犁骨(prevomer, PV, 又名vomer)似三角形,位于腦顱前端腹側,上接中篩骨軟骨(ethmoid cartilage, EC)。
中篩骨(ethmoid, E, 又名mesethmoid)位于腦顱前部中線處,似長橢圓體,前側形成關節面,接觸吻軟骨。
側篩骨(lateral ethmoid, LET, 曾用名parethmoid和ectoethmoid)如翼狀,構成眼眶前緣,上有供嗅神經穿過的嗅神經孔(foramen for olfactory nerve, FON),側面有上下2個關節頭,分別與第1眶下骨和顎骨關節面接觸。
額骨(frontal, FRO)位于腦顱背面前半部分,構成眼眶上緣,上有眶上管和眶上連管(supraorbital commissure)的管道溝槽。
頂骨(parietal, PA)位于腦顱背面中央偏后區,呈鋸齒邊片狀。側線副神經支(ramus lateralis accessorius)從其孔穿出。緊密接合周圍各骨。
蝶耳骨(sphenotic, SPO)位于腦顱中部外側,構成眼眶后上緣,緊密接合周圍各骨,在與前耳骨的接縫處形成關節窩,與舌頜骨前上部髁突接觸。
翼蝶骨(pterosphenoid, PTS,曾用名alisphenoid)位于眼眶內側后上方,后方通過軟骨帶接前耳骨。
前耳骨(prootic, PRO)構成眼眶后緣以及耳囊的前側和外側面。
翼耳骨(pterotic, PTO)位于腦顱后外角,外側面中部有一個小窩,與舌頜骨中上部髁突相接。
上耳骨(epiotic, EPO)位于腦顱后上方,與周圍除頂骨外的骨頭通過軟骨帶連接,后顳骨內臂通過強韌結締組織與其連接。
后耳骨(intercalar, IC, 曾用名opisthotic)位于腦顱外側面的后部,與周圍骨頭緊密接合。
上枕骨(supraoccipital, SO)處于腦顱后上部中間,具一個向上凸起的不發達的脊。
側枕骨(exoccipital, EO)位于腦顱后方中間部,左右側枕骨后方構成枕骨大孔,神經從內穿出,其后端關節連接第1椎骨。
基枕骨(basioccipital, BO)是腦顱后下部骨頭,其后端關節連接第1椎骨。
副蝶骨(parasphenoid, PAS)是腦顱底部中軸處長骨。它后部分叉于兩側,緊密貼合基枕骨。
2.1.3 頜部(jaws)(圖3) 頜骨位于魚頭的前下部,腦顱外側,包括上頜和下頜兩部分。上頜包括前頜骨和上頜骨,下頜包括齒骨、關節骨和隅骨,皆左右成對。
前頜骨(premaxilla, PMX, 曾用名premaxillary)呈L型,由上升突(ascending process, ACP)、關節突(articular process, ARP)、牙槽突(alveolar process, ALP)和吻軟骨(rostral cartilage, RC)組成。牙槽突的腹面有齒板,吻軟骨附著于上升突上,與中篩骨及中篩骨軟骨接觸。
上頜骨(maxilla, MAX, 曾用名maxillary)呈扁條狀,前端膨大似半球面,前頜骨的關節突嵌合于其中。
齒骨(dentary, DEN)是下頜的前半部分,是深槽的V形骨,上方具齒板,后方關節骨和美克耳氏軟骨(Meckelian cartilage, MEC, 又名麥克氏軟骨)插入其深槽內。鰓蓋下頜管(operculomandibular sensory canal)穿過該骨,向外有4個開口即為下頜部黏液孔。
關節骨(anguloarticular, ANG, 曾用名articular和angular)是下頜的后半部分,后上方突起與方骨下端突起相嵌合。美克耳氏軟骨附著于該骨內側面,鰓蓋下頜管穿過該骨。
隅骨(retroarticular, RA, 曾用名angular)是一塊類三角形骨頭,緊密接合于關節骨上,上方有一美克耳氏板小骨(coronomeckelian, COM)很小,位于關節骨與美克耳氏軟骨的夾縫間。

圖3 上頜骨外側面圖(左上)和內側面圖(右上),下頜骨外側面圖(左下)和內側面圖(右下)
ACP:上升突;ALP:牙槽突;ANG:關節骨;ARP:關節突;COM:美克耳氏板小骨;DEN:齒骨;MAX:上頜骨;MEC:美克耳氏軟骨(麥克氏軟骨);PMX:前頜骨;RA:隅骨(后關節骨);RC:吻軟骨
ACP: Ascending process; ALP: Alveolar process; ANG: Anguloarticular; ARP: Articular process; COM: Coronomeckelian; DEN: Dentary; MAX: Maxilla; MEC: Meckelian cartilage; PMX: Premaxilla; RA: Retroarticular; RC: Rostral cartilage
2.1.4 懸器和鰓蓋骨(suspensorium and opercular bones)(圖4) 懸器在腦顱的左右兩側,下部被上頜覆蓋,下接舌弓,包括腭骨、翼骨+中翼骨復合體、后翼骨、舌頜骨、方骨和續骨。鰓蓋骨接于懸器后方,是構成鰓孔的重要部分,包括前鰓蓋骨、鰓蓋骨、下鰓蓋骨和間鰓蓋骨,皆左右成對。
腭骨(palatine, PAL)似扭曲的三角形,位于懸器最前部,其前端有一軟骨蓋,與上頜骨、第1眶下骨的關節面相接觸。其內側有一關節面與側篩骨下關節頭接觸。
翼骨+中翼骨復合體是由翼骨(ectopterygiod, ECP, 曾用名pterygoid)和中翼骨(endopterygiod, ENP, 曾用名mesopterygoid)融合而成,似梯形片狀,其后下方通過軟骨帶連方骨(不接觸),此軟骨帶向前延伸,橫貫本骨外側面中部。
后翼骨(metapterygoid, MEP)呈近似矩形的薄片狀,與舌頜骨的接合處有一神經孔。
舌頜骨(hyomandibula, HYO)呈倒L形,下部內側通過軟骨帶連接續骨和間舌骨。舌頜骨的上方有 2個髁突,與腦顱側面2個小窩接觸;向后延伸的長柄末端有第3個髁突,與鰓蓋骨前端的凹槽接觸。
方骨(quadrate, QUA)呈三角形,其內側面有一個凹槽,續骨的下部插入其中。
續骨(symplectic, SYM)是上下皆有軟骨蓋的棒狀骨頭,其后方與前鰓蓋骨之間有一個大間隙。
前鰓蓋骨(preopercle, POP)是鰓蓋骨中最大的,呈彎月狀。鰓蓋下頜管穿過該骨。
鰓蓋骨(opercle, 又稱主鰓蓋骨, OPC)呈細長三角形,下方內側與下鰓蓋骨接觸。
下鰓蓋骨(subopercle, SOP, 又名infraopercle)呈V形,兩枝都較細長。
間鰓蓋骨(interopercle, IOP)很細長,位于前鰓蓋骨的后方,下鰓蓋骨前方和鰓蓋骨下方,游離于其他骨頭,僅靠肌肉和韌帶連接。
2.1.5 舌弓(hyoid arch)(圖5) 舌弓位于下頜的內側、下鰓弓的外側,后上方連接懸器內側,包括下舌骨、角舌骨、上舌骨、間舌骨、鰓條骨和尾舌骨。無基舌骨(basihyal)。僅尾舌骨不成對。

圖4 懸器和鰓蓋骨的外側圖(左)和內側圖(右)
ECP+ENP:翼骨+中翼骨復合體;HYO:舌頜骨;IOP:間鰓蓋骨;MEP:后翼骨;OPC:(主)鰓蓋骨;PAL:腭骨;POP:前鰓蓋骨;QUA:方骨;SOP:下鰓蓋骨;SYM:續骨
ECP+ENP: Ectopterygiod+endopterygiod; HYO: Hyomandibula; IOP: Interopercle; MEP: Metapterygoid; OPC: Opercle; PAL: Palatine; POP: Preopercle; QUA: Quadrate; SOP: Subopercle; SYM: Symplectic

圖5 舌弓骨的外側圖(左)和內側圖(右),尾舌骨(左下)
BF:Beryciform foramen;BR:鰓條骨;CH:角舌骨;DHH:上下舌骨;EPH:上舌骨;IH:間舌骨;VHH:下下舌骨
BF: Beryciform foramen; BR: Branchiostegal ray; CH: Ceratohyal; DHH: Dorsal hypohyal; EPH: Epihyal; IH: Interhyal; VHH: Ventral hypohyal
下舌骨(hypohyal)位于舌弓的前部,由一條軟骨帶分為上下舌骨(dorsal hypohyal, DHH)和下下舌骨(ventral hypohyal, VHH)。左右側下舌骨于前端通過韌帶緊緊連接。
角舌骨(ceratohyal, CH)較大且厚,后方通過軟骨帶連接上舌骨,前下方內側和后下方外側分別附著 2條鰓條骨。其中央位置有1個孔(beryciform foramen, BF),舌動脈從其中穿過。
上舌骨(epihyal, EPH)呈三角形,后上方的凹槽與間舌骨下部軟骨蓋相接觸,下方外側附著2條鰓條骨。
間舌骨(interhyal, IH)呈棒狀,上下都有軟骨蓋,下方與上舌骨的凹糟相接觸,上方與舌頜骨下軟骨帶相接觸。
鰓條骨(branchiostegal ray, BR)每側有6條,皆呈細長弧形,上端附著于角舌骨或上舌骨,下端游離,靠肌肉拉動張合。
尾舌骨(urohyal, URO)位于下舌骨前端和第1基鰓骨的下方,呈半月狀。
2.1.6 鰓弓(branchial arches)(圖6) 鰓弓分為上下2部分,上鰓弓位于腦顱后下方、鰓蓋的內側,下鰓弓位于舌弓內側。上鰓弓包括上鰓骨和咽鰓骨,下鰓弓包括基鰓骨、下鰓骨、角鰓骨和中央軟骨。除基鰓骨和中央軟骨外,其余皆左右成對。
基鰓骨(basibranchial, BB)位于下鰓弓中線處,也是口腔腹面的中線處。第1基鰓骨為硬骨,位于最前端,呈較小的卵形,覆蓋于尾舌骨上,后方接觸由第2和第3基鰓骨融合形成的第2+3基鰓骨復合體,該復合體為軟骨,呈長圓柱狀,向后接觸中央軟骨,其左右兩側腹面接觸3對下鰓骨。

圖6 下鰓弓骨的背側圖(左上)和腹側圖(右上),上鰓弓骨的背側圖(左下)和腹側圖(右下)
BB1:第1基鰓骨;BB 2+3:第2+3基鰓骨復合體;CB 1~5:第1~5角鰓骨;CC:中央軟骨;EB 1~4:第1~4上鰓骨;HB 1~3:第1~3下鰓骨;PB:咽鰓骨;tooth plate:齒板
BB1: First basibranchial; BB 2+3: Second+third basibranchial; CB 1~5: First to fifth ceratobranchials; CC: Central cartilage; EB 1~4: First to fourth epibranchials; HB 1~3: First to third hypobranchials; PB: Pharyngobranchial
中央軟骨(central cartilage, CC),或被認作第4基鰓骨,為橢圓狀軟骨,與兩側第4和第5角鰓骨接觸。
下鰓骨(hypobranchial, HB)共3對,內側端有軟骨蓋,外邊緣有軟骨帶。第1~3下鰓骨外側分別與第1~3角鰓骨之間通過關節相連。
角鰓骨(ceratobranchial, CB)共5對。第1~3角鰓骨為長條狀,腹側有凹糟,后端以關節分別連接第1~3上鰓骨后端。第4角鰓骨呈弧形條狀,后端以關節連接第4上鰓骨后端。第5角鰓骨又名咽骨(pharyngeal bone),背生有齒板(tooth plate),附著咽齒(pharyngeal teeth),后端游離。
上鰓骨(epibranchial, EB)共4對,兩端有軟骨蓋,第1上鰓骨為細長棒狀,前端游離。第2上鰓骨呈略粗的棒狀,前端與咽鰓骨外側軟骨接觸。第3和第4上鰓骨的側面分支彼此相連,前端與咽鰓骨后部大軟骨接觸。
咽鰓骨(pharyngobranchial, PB)僅有1對。第1、2和4咽鰓骨缺失,第3咽鰓骨發達且腹面生有大齒板。
2.1.7 肩帶(pectoral girdle)(圖7) 肩帶位于魚頭部后方,構成鰓孔的后緣,與所承擔的鰭條共同構成胸鰭的骨骼框架。該部位骨頭皆左右成對,每側包括上顳骨、后顳骨、上匙骨、匙骨、肩胛骨、烏喙骨、和后匙骨各1塊,以及4塊支鰭骨。肩胛骨、烏喙骨和支鰭骨都支撐鰭條,連通的軟骨帶填充在肩胛骨、匙骨、烏喙骨、支鰭骨和鰭條之間。

圖7 肩帶骨的外側圖(左)和內側圖(中),后匙骨圖(右)
ACT:支鰭骨;CLE:匙骨;COR:烏喙骨;IF:支鰭骨間孔;PCL:后匙骨;PT:后顳骨;SCA:肩胛骨;SCL:上匙骨;ST:上顳骨;MLPT:后顳骨內臂
ACT: Actinost; CLE: Cleithrum; COR: Coracoid; IF: Interradial foramen; PCL: Postcleithrum; PT: Posttemporal; SCA: Scapula; SCL: Supracleithrum; ST: Supratemporal; MLPT: Medial limb of posttemporal
上顳骨(supratemporal, ST, 又名extrascapular)位于肩帶最上部,具不完全管狀結構,連通翼耳骨背面與后顳骨的管道。下方貼合在翼耳骨背面后端。
后顳骨(posttemporal, PT)分內外2個部分,外側部分有溝槽結構,經由上匙骨的感覺管通向前方;內側部分為長方形片狀,稱為后顳骨內臂(medial limb of posttemporal, MLPT),以強韌的結締組織緊密貼于上耳骨及其后方軟骨帶背面。
上匙骨(supracleithrum, SCL, 曾用名supraclavicle)的上部向后形成薄片狀扇面,有一溝槽供感覺管通過,下部為長棒狀,貼于匙骨上半部分外側。
匙骨(cleithrum, CLE, 曾用名clavicle)是肩帶最大的骨頭,呈弧形,下部分別向內外兩側擴張形成片狀板,后接肩胛骨、烏喙骨和周圍的軟骨板,形成的間隙供肩帶肌肉附著。兩側匙骨在前端通過結締組織相連。
肩胛骨(scapula, SCA)像缺了一角的正方形,通過軟骨帶向前連接匙骨,向下連接第1支鰭骨。肩胛骨孔(scapular foramen)由肩胛骨、匙骨和軟骨帶共同圍成。
烏喙骨(coracoid, COR)位于肩帶前下部,并通過軟骨帶向后上方連接第4支鰭骨。
支鰭骨(actinost, ACT,曾用名radial)由上至下排列于匙骨下部的后側,皆由軟骨帶包圍。第1支鰭骨上下各有1個支鰭骨間孔(interradial foramen, IF),前3個支鰭骨和第4支鰭骨之間距離很大。
后匙骨(postcleithrum, PCL, 曾用名postclavicle)很細長,貼于匙骨和上匙骨的內側,后下部深入腹壁肌肉。
2.1.8 腰帶(pelvic girdle)(圖8) 腰帶位于魚頭后下方腹面,外觀呈盤狀,其上部結構夾于左右匙骨之間。腹鰭骨骼包括1對腰骨、1對鰭棘和5對鰭條。
腰骨(pelvis,P, 又名basipterygia)包括前上部的扇形板(fan-shaped plate, FP)和前下部的下腰骨突(subpelvic process, SP),以及后側的擔鰭部分。扇形板向上插入左右匙骨之間,其上緣有軟骨帶,軟骨帶頂端和底端分別以關節連接匙骨內側中部和前下部的軟骨位點。下腰骨突向前延伸,并擔起鰭棘,鰭棘呈彎曲的縫合針狀。后側的擔鰭部分左右相對,其中間形成一個空隙,附著肌肉,兩邊外側腹面各附著 5條鰭條,鰭條的末端特化成半筒狀舌形結構。

圖8 腰帶骨的背側圖(上)﹑腹側圖(中)和左側圖(下)
FP:扇形板;P:腰骨;R:鰭條;S:鰭棘;SP:下腰骨突
FP: Fan-shaped plate; P: Pelvis; R: Ray; S: Spine; SBP: Subpelvic process
2.1.9 中軸骨架和奇鰭支鰭骨(axial skeleton and median fin supports)(圖9~圖10) 中軸骨骼由椎骨、脈弓小骨和腹肋組成。
椎骨分為軀椎(abdominal vertebrae)和尾椎(caudal vertebrae) 2種。前12節為軀椎,包括椎體(centrum, CT)、髓弓(neural arch)、髓棘(neural spine, NS, 又名神經棘)和椎管,除前2節之外都有椎體橫突(parapophysis)。后34節為尾椎,下部有脈弓(haemal arch, HA)和脈棘(hemal spine, HS, 又名血管棘)。

圖9 背鰭前端與部分腹椎骨骼圖
CT:錐體;DPD:背鰭末節;EPR:脈弓小骨;HA:脈弓;NS:髓棘;PPD:背鰭基節;R:鰭條
CT: Centrum; DPD: Distal pterygiophore of dorsal fin; EPR: Epipleural rib; HA:Haemal arch; NS: Neural spine; PPD: Proximal pterygiophore of dorsal fin; R: Ray

圖10 臀鰭前端與相關脊椎骨骼圖
CT:錐體;DPA:臀鰭末節;EPR:脈弓小骨;HS:脈棘;PPA:臀鰭基節;PR:腹肋;R:鰭條
CT: Centrum; DPA: Distal pterygiophore of anal fin; EPR: Epipleural rib; HS:Haemal spine; PPA: Proximal pterygiophore of anal fin; PR: Pleural rib; R: Ray
脈弓小骨(epipleural rib, EPR, 曾用名hypomeralia)每側有30條,附著于第2~31椎骨。
腹肋(pleural rib, PR, 又名ventral rib)每側有3條,分別附著于第10~12節椎骨的椎體橫突下端。
奇鰭包括背鰭和臀鰭,兩鰭延長至尾鰭部,主要由不分枝的鰭條、末節(distal pterygiophore, 又名遠位支鰭骨, 曾用名distal radial)和基節(proximal pterygiophore, 又名近位支鰭骨, 曾用名basal radial)組成,無鰭棘。左右兩側鰭條將末節夾在中間,無中節(medial pterygiophore, 又名間位支鰭骨, 曾用名median radial)、上髓棘(supraneural)和終端骨(stay)。
背鰭末節(distal pterygiophore of dorsal fin, DPD)是位于背鰭鰭條和基節之間的小骨頭,自第6鰭條下開始出現。
背鰭基節(proximal pterygiophore of dorsal fin, PPD)是細長的棒狀骨,第1基節退化為軟骨棒,不支撐鰭條。第2基節位于第3和第4椎骨的神經棘之間,之后與神經棘依次穿插排列。
臀鰭末節(distal pterygiophore of anal fin,DPA)形態結構與背鰭末節相似,自第3鰭條下開始出現。
臀鰭基節(proximal pterygiophore of anal fin,PPA)形態結構與背鰭基節相似,前2個臀鰭基節退化變小,支撐鰭條。
2.1.10 尾骨(caudal skeleton)(圖11) 尾部骨架包括尾椎前椎體、尾上骨、上尾下骨、下尾下骨+準尾下骨復合體及它們周圍的軟骨和鰭條。無尾髓棘(uroneural, 又名尾神經骨)。

圖11 尾鰭骨骼圖
CIEP:尾上骨周軟骨;CIHPU:尾椎前脈棘周軟骨;EU:尾上骨;HS:脈棘;LHYP+PHY:下尾下骨+準尾下骨復合體;NS:髓棘;PU2:第2尾椎前椎體;UHYP:上尾下骨;US:尾桿骨
CIEP: Inter-epural cartilage; CIHPU: Inter-hemal spine cartilage of preural; EU: Epural; HS: Hemal spine; LHYP+ PHY: Lower hypural+Parhypural; NS: Neural spine; PU2: Second preural centrum; UHYP: Upper hypural; US: Urostyle
尾椎前椎體包括尾桿骨(urostyle, US, 既第1尾椎前椎體preural centrum 1)和第2尾椎前椎體(preural centrum 2, PU2)。尾桿骨是最后一節椎骨,蝌蚪狀,末梢向斜上方伸出。第2尾椎前椎體的椎體較短,上部接髓棘,下部脈弓膨大接脈棘并支撐一條前傾鰭條,脈棘末端有一塊小軟骨,稱為尾椎前脈棘周軟骨(inter-hemal spine cartilage of preural, CIHPU)。
尾上骨(epural, EU)有2塊,位于尾骨后上部,支撐2條前傾鰭條。較大的一塊為倒T形,較小的一塊為棒狀,后方有一塊橢圓形軟骨,即尾上骨周軟骨(inter-epural cartilage, CIEP)。
尾下骨(hypural)分為2個部分,上方的稱為上尾下骨(upper hypural, UHYP),下方的稱為下尾下骨(lower hypural, LHYP),下尾下骨與準尾下骨融合形成復合體(詳見準尾下骨)。兩部分骨板似三角形,分別位于尾鰭水平線的上下兩側,構成尾部扇形結構主體。
準尾下骨(parhypural)位于尾下骨下方,第2尾椎前椎體脈棘的上方。準尾下骨與下尾下骨融合形成下尾下骨+準尾下骨復合體(LHYP+PHY),準尾下骨側突(parhypurapophysis)不明顯,骨板前部有一孔,前下部有一個倒鉤狀骨片。
尾鰭鰭條不分叉,分為2種:前傾鰭條(procurrent ray)和主鰭條(principal ray)。
2.2 肌肉學
2.2.1 頰肌(cheek muscles)(圖12~圖13) 頰肌位于魚頭兩側下頜及懸器區域,控制口的閉合,與攝食和呼吸動作相關,主要指的是下頜收肌(adductor mandibulae, 曾用名adductor mandibularis)系列肌肉。細紋獅子魚的下頜收肌分化成4部分:分肌1~3和分肌ω。分肌1又分為α和β兩部分,皆左右對稱出現。
下頜收肌分肌1 (adductor mandibulae section 1)又稱為頭部分肌a (cephalic portion a),分化為外側的α和內側的β兩個部分,即下頜收肌分肌1α (A1α)和分肌1β (A1β)。A1α是魚頭最外側的肌肉,收縮使頜骨向后方移動,起始于前鰓蓋骨下部、眶下骨架下緣、舌頜骨下緣和方骨上緣,止于原始韌帶(primordial ligament, PL, 又名ligamentum primordium)后部。A1β向內分出一部分稱為下頜收肌分肌1β′ (A1β′),其肌腱與A1β韌帶前端融合,向前連于上頜骨頭部內側,A1β韌帶的下端連于關節骨內側,該韌帶的后部與原始韌帶和A2的肌腱融合。A1β起于舌頜骨和前鰓蓋骨上部,收縮使頜骨向后上方移動;A1β′起始于后翼骨,收縮可使上頜骨向內向上運動。

圖12 頰肌圖:外側面全圖(上),移除眶下骨和A1α肌后圖(中),再移除A1β和A2后圖(下)
A1α、A1β、A1β′、A2和A3分別為下頜收肌分肌A1α、A1β、A1β′、A2和A3;ANG:關節骨;DEN:齒骨;DO:鰓蓋開肌;HYO:舌頜骨;IO:眶下骨;SS:眶下骨架;LAP:腭弓提肌;LO:鰓蓋提肌;MAX:上頜骨;MEP:后翼骨;OPC:(主)鰓蓋骨;PAL:腭骨;PL:原始韌帶;PMX:前頜骨;POP:前鰓蓋骨;PTO:翼耳骨;QUA:方骨;SCL:上匙骨;SPO:蝶耳骨;SYM:續骨
A1α, A1β, A1β′, A2, and A3 indicate adductor mandibulae section A1α, A1β, A1β′, A2, and A3, respectively; ANG: Anguloarticular; DEN: Dentary; DO: Dilatator operculi; HYO: Hyomandibula; IO: Infraorbital; SS: Suborbital stay; LAP: Levator arcus palatini; LO: Levator operculi; MAX: Maxilla; MEP: Metapterygoid; OPC: Opercle; PAL: Palatine; PL: Primordial ligament; PMX: Premaxilla; POP: Preopercle; PTO: Pterotic; QUA: Quadrate; SCL: Supracleithrum; SPO:Sphenotic; SYM: Symplectic
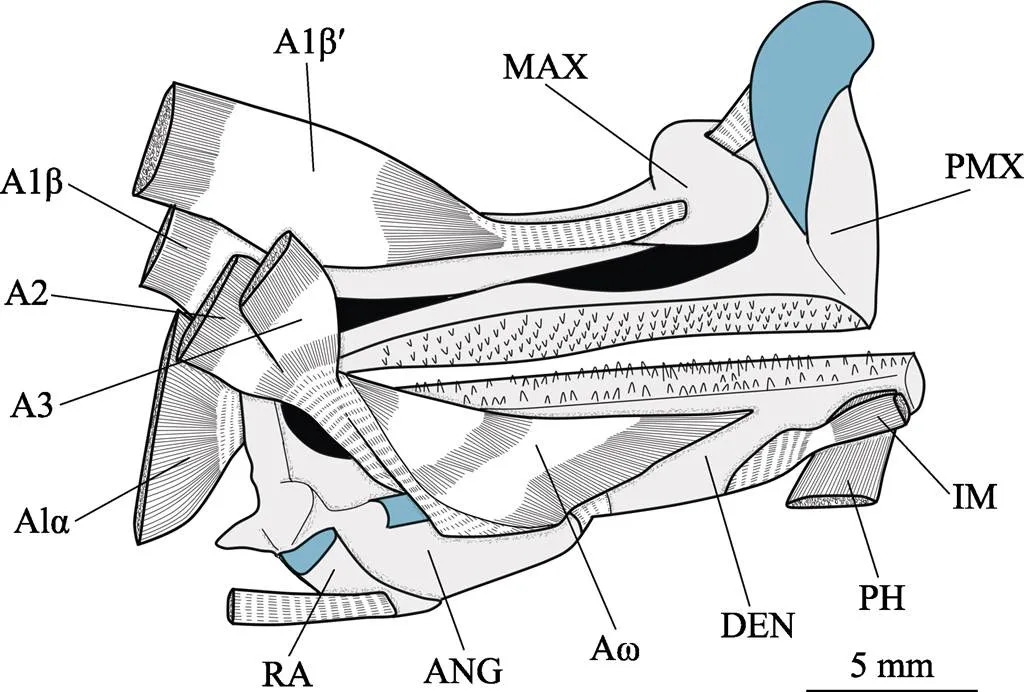
圖13 頜骨內側肌肉圖
A1α、A1β、A1β′、A2、A3和Aω分別為下頜收肌分肌A1α、A1β、A1β′、A2、A3和Aω;ANG:關節骨;DEN:齒骨;IM:下頜間肌;MAX:上頜骨;PH:頦舌肌;PMX:前頜骨;RA:隅骨
A1α, A1β, A1β′, A2, A3, and Aω indicated adductor mandibulae section A1α, A1β, A1β′,A2, A3 and Aω, respectively; ANG: Anguloarticular; DEN: Dentary; IM: Intermandibularis; MAX: Maxilla; PH: Protractor hyohyoidei; PMX: Premaxilla; RA:Retroarticular
下頜收肌分肌2 (adductor mandibulae section 2, A2)又稱為頭部分肌b (cephalic portion b),起始于后翼骨、舌頜骨下部、前鰓蓋骨中部和眶下骨架上緣,止于關節骨內側,收縮使下頜骨向后上方移動。A2與A3下部融合。三叉神經下頜枝(ramus mandibularis trigeminus)從A2和A1β間穿出。
下頜收肌分肌3 (adductor mandibulae section 3, A3)起始于后翼骨,下部與A2融合,具有相同止點,收縮使下頜骨向內向上移動。
下頜收肌分肌ω (adductor mandibulae section ω,Aω)又稱為下頜部分肌(mandibular portion),附著于齒骨和關節骨的內側,后部連接原始韌帶和A2、A3的肌腱,收縮時可協同其他分肌將口閉合。
2.2.2 腦顱與懸器鰓蓋骨間肌肉(cephalic muscles between cranium and suspensorium-opercular bones) (圖12、圖14和圖15) 該部肌肉位于頭骨和懸器鰓蓋骨之間,司該部分的運動和鰓蓋的開合,包括腭弓提肌、腭弓收肌、鰓蓋開肌、鰓蓋收肌、舌頜收肌和鰓蓋提肌,皆左右對稱。
腭弓提肌(levator arcus palatini, LAP)較強壯,起始于后翼骨和舌頜骨上部,止于蝶耳骨下部和翼耳骨外側,收縮可使懸器上提。
腭弓收肌(adductor arcus palatini, AAP)起始于副蝶骨外側,少量附于側篩骨內后側,止于腭骨內側與翼骨+中翼骨復合體的上緣,皆以腱膜連接,收縮使口角提起,懸器內收。

圖14 腦顱與懸器鰓蓋骨間肌肉:外側面圖(上),懸器鰓蓋骨內側圖(下)
AAP:腭弓收肌;AH:舌頜收肌;AO:鰓蓋收肌;ECP+ENP:翼骨+中翼骨復合體;FRO:額骨;HAD:鰓蓋條舌肌;HYO:舌頜骨;LAP:腭弓提肌;LE2:第2鰓弓外提肌;LO:鰓蓋提肌;MEP:后翼骨;OPC:(主)鰓蓋骨;PAS:副蝶骨;PTO:翼耳骨;LET:側篩骨;SPO:蝶耳骨
AAP: Adductor arcus palatini; AH: Adductor hyomandibulae; AO: Adductor opercula; ECP + ENP: Ectopterygiod + Endopterygiod; FRO: Frontal; HAD: Hyohyoideus adductores; HYO: Hyomandibula; LAP: Levator arcus palatini; LE2: Second levator externus; LO: Levator opercula; MEP: Metapterygoid; OPC: Opercle; PAS:Parasphenoid; PTO: Pterotic; LET: Lateral ethmoid; SPO: Sphenotic
鰓蓋開肌(dilatator operculi, DO, 曾用名dilatator operculis)起始于翼耳骨外側前部,止于鰓蓋骨的前端,收縮可使鰓蓋張開。
舌頜收肌(adductor hyomandibulae, AH)短粗且強壯,毗鄰鰓蓋收肌,控制懸器和鰓蓋的張合。起始于翼耳骨外側,止于舌頜骨內側上部。
鰓蓋收肌(adductor operculi, AO)不發達,位于舌頜收肌后側,收縮使鰓蓋閉合。起始于翼耳骨外側,止于鰓蓋骨前端內側。

圖15 鰓弓和胸鰭外側面肌肉圖
AH:舌頜收肌;ABP:肩帶深層展肌;ABS:肩帶淺層展肌;AO:鰓蓋收肌;ARV:肩帶腹側立肌;LE 1~4:第1~4鰓弓外提肌;LEP:胸鰭提肌;LI2:第2鰓弓內提肌;LP:鰓弓后提肌;PCE:外咽匙肌;PCI:內咽匙肌;PPEC:胸鰭牽引肌;RC:鰓弓縱直肌;RVc:鰓弓腹直肌分肌c;STH:胸舌肌;URO:尾舌骨
AH: Adductor hyomandibulae; ABP: Abductor profundus; ABS: Abductor superficialis; AO: Adductor operculi; ARV: Arrector ventralis; LE 1~4: First to fourth levator externus, respectively; LEP: Levator pectoralis; LI2: Second levator internus; LP: Levaror posterior; PCE: Pharyngoclavicularis externus; PCI: Pharyngoclavicularis internus; PPEC: Protractor pectoralis; RC: Rectus communis; RVc: Rectus ventralis section c; STH: Sternohyoideus; URO: Urohyal
鰓蓋提肌(levator operculi, LO)有三四條薄肌肉束,起點和止點分別位于上匙骨的扇形薄片和鰓蓋骨的內側上緣,收縮使鰓蓋骨提起。
2.2.3 頭部腹面肌肉(ventral muscles of head)(圖14、圖16和圖17) 頭部腹面肌肉主要包括下頜間肌、頦舌肌、舌弓展肌、舌肌和鰓蓋條舌肌,控制下頜和舌弓的運動。除下頜間肌之外,其余左右對稱。
下頜間肌(intermandibularis, IM)很短小,附著于左右齒骨前部內側面,收縮使兩側齒骨彼此靠近,與下方頦舌肌相鄰。
頦舌肌(protractor hyohyoidei, PH, 曾用名geniohyoideus)發達,起始于齒骨前端內側,向后延伸至中前部,兩側肌肉經肌隔接合,再分別向左右延伸,止于角舌骨外側中下部與第3鰓條骨上部。每側均由2個肌隔間隔成3段,控制下頜骨與舌弓骨以及左右兩側的位置關系,不同段的收縮對呼吸活動產生作用,吸氣時后段收縮,呼氣時前段收縮。

圖16 頭部腹面肌肉圖
BR:鰓條骨;CH:角舌骨;DEN:齒骨;HAB2:舌弓展肌分肌2;HIF:舌肌;IM:下頜間肌;PH:頦舌肌
BR: Branchiostegal ray; CH: Ceratohyal; DEN: Dentary; HAB 2: Second hyohyoidei abductor; HIF: Hyohyoides inferioris; IM: Intermandibularis; PH: Protractor hyohyoidei
舌弓展肌分為2個部分:上側的分肌1 (hyohyoidei abductor section 1, HAB1)和下側的分肌2 (hyohyoidei abductor section 2, HAB2)。HAB1有4束,控制鰓條骨的貼伏和直立,前3束起始于角舌骨內側前端底部,最后一束起始于第2鰓條骨內側頂端,4束肌肉依次止于前4條鰓條骨內側上部。HAB2起始于第1鰓條骨的中下部,向魚頭腹面中線延伸,兩側經中縫(median raphe)接合,收縮可使兩側第1鰓條骨彼此靠近。
舌肌(hyohyoides inferioris, HIF, 曾用名hyohyoideus)由角舌骨內側中后部底部起始,向前穿過HAB1和鰓條骨的間隙,再向魚頭腹面中線延伸,兩側經中縫接合,前側有透明的薄腱膜(幾乎不可見)連接下下舌骨和角舌骨腹側。該肌收縮可使兩側舌弓相互靠近。
鰓蓋條舌肌(hyohyoidei adductores, HAD,又名鰓條收肌, 曾用名branchiostegohyoideus)附著于鰓條骨下部,后方連接鰓蓋骨和下鰓蓋骨的內側,控制鰓條骨間開合。

圖17 舌弓肌肉圖:外側面(左)和內側面(右)
BR:鰓條骨;CH:角舌骨;HAB1、2:舌弓展肌分肌1、2;HAD:鰓蓋條舌肌;HIF:舌肌;PH:頦舌肌;RVc:鰓弓腹直肌分肌c
BR: Branchiostegal ray; CH: Ceratohyal; HAB1 and HAB2 indicate hyohyoidei abductors section 1 and section 2, respectively; HAD: Hyohyoidei adductor; HIF: Hyohyoides inferioris; PH: Protractor hyohyoidei; RVc: Rectus ventralis section c
2.2.4 鰓弓肌肉(branchial muscles)(圖15和圖18)
鰓弓部肌肉包括鰓弓提肌(levator arcum branchialium, 即鰓弓外提肌、鰓弓內提肌和鰓弓后提肌)、背面橫肌、鰓間背斜肌、鰓間后斜肌、鰓弓背面牽縮肌、鰓弓縱直肌、咽匙肌、鰓弓腹直肌及分肌c和u、鰓間腹斜肌、腹面橫肌、鰓弓收肌以及食道括約肌。
鰓弓外提肌(levator externus)分為4個部分,即第1~4鰓弓外提肌(first to fourth levator externus, LE1~4),分別起始于第1~4上鰓骨,分別止于蝶耳骨外側(腱膜連接)、蝶耳骨下側面和眼眶內側、前耳骨關節窩下方以及舌頜骨內側上部,收縮可使上鰓骨上提。
鰓弓內提肌(levator internus)分為2個部分,即第1和第2鰓弓內提肌(first and second levator internus, LI1~2),分別起始于咽鰓骨背面前端和后部,止于前耳骨上緣和翼耳骨下部,收縮可使咽鰓骨上提。
鰓弓后提肌(levator posterior, LP)起始于第4上鰓骨的末端軟骨,止于后耳骨,收縮可使第4上鰓骨向后上方移動。
背面橫肌(transversus dorsalis, 曾用名transversi dorsalis)分為背面前橫肌(transversus dorsalis anterior, TDA)和背面后橫肌(transversus dorsalis posterior, TDP) 2部分,它們在相接處部分融合。前者連接第2上鰓骨前部,后延伸至中縫左右接合;后者連接第3、第4上鰓骨前部,后延伸至中縫左右接合。背面橫肌收縮使兩側連接的上鰓骨彼此靠近,上鰓弓向下拱起。

圖18 鰓弓肌肉圖:下鰓弓腹面(上)和上鰓弓背面(下)
AD:鰓弓收肌;LE 1~4:第1~4鰓弓外提肌;LI1~2:第1和第2鰓弓內提肌;LP:鰓弓后提肌;OBD:鰓間背斜肌;OBP:鰓間后斜肌;OBV 1~3:第1~3鰓間腹斜肌;PCE:外咽匙肌;PCI:內咽匙肌;RC:鰓弓縱直肌;RD:鰓弓背面牽縮肌;RV:鰓弓腹直肌;RVc:鰓弓腹直肌分肌c;RVu:鰓弓腹直肌分肌u;SOE:食道括約肌;TDA:背面前橫肌;TDP:背面后橫肌;TVA:腹面前橫肌;TVP:腹面后橫肌
AD: Adductor; LE 1~4: First to fourth levator externus, respectively; LI 1~2: First and second levator internus; LP: Levator posterior; OBD: Obliquus dorsalis; OBP: Obliquus posterior; OBV 1~3: First to third obliquus ventralis, respectively; PCE: Pharyngoclavicularis externus; PCI: Pharyngoclavicularis internus; RC: Rectus communis; RD: Retractor dorsalis; RV: Rectus ventralis; RVc: Rectus ventralis from ceratohyal; RVu: Rectus ventralis from urohyal; SOE: Sphincter oesophagi; TDA: Transversus dorsalis anterior; TDP: Transversus dorsalis posterior; TVA: Transversus ventralis anterior, TVP: Transversus ventralis posterior
鰓間背斜肌(obliquus dorsalis, OBD, 曾用名interarcualis dorsalis obliqus)起始于咽鰓骨前部,止于第3和第4上鰓骨前部,前部夾于咽鰓骨和背面后橫肌之間,收縮使第3和第4上鰓骨靠近咽鰓骨。
鰓弓背面牽縮肌(retractor dorsalis, RD)連接咽鰓骨及其后側皮膚與第4~8節椎體的腹面,收縮使鰓弓后移,功能與咽骨退縮肌(retractor ossium pharyngealium)相似,推測為同源。
鰓弓縱直肌(rectus communis, RC)起始于尾舌骨的上緣,止于第5角鰓骨的腹面中外部,與外咽匙肌的止點相鄰。
咽匙肌位于下鰓弓中部,分為外咽匙肌(pharyngoclavicularis externus, PCE)和內咽匙肌(pharyngoclavicularis internus, PCI) 2種。外咽匙肌起始于胸鰭匙骨下部背面,止于第5角鰓骨腹面中外部,收縮使咽骨有下壓的趨勢。內咽匙肌起始于胸鰭匙骨下部背面,外咽匙肌起點的后側,止于第5角鰓骨腹面的前端,其前部夾在腹面前橫肌與腹面后橫肌之間,收縮使咽骨有縮向食道的趨勢。
鰓弓腹直肌(rectus ventralis, RV, 曾用名鰓弓連肌interarcuales branchialis)連接第4角鰓骨腹面的前端與第3下鰓骨腹面的前外側,收縮使第4角鰓骨與第3下鰓骨相靠近,下鰓弓隆起。
鰓弓腹直肌分肌c (rectus ventralis from ceratohyal, RVc, 曾用名鰓弓牽引肌protractor arcuum branchialum、角舌角鰓肌ceratohyoceratobranchialis)起始于角舌骨內側上緣,止于第1角鰓骨腹面前端,下部與鰓間腹斜肌分肌1接合,收縮使角舌骨與第1角鰓骨彼此靠近。
鰓弓腹直肌分肌u (rectus ventralis from urohyal, RVu)有2束前端融合的肌肉,它們的起點分別位于第2和第3下鰓骨腹面的前內側,止點位于尾舌骨上緣,與鰓弓縱直肌相鄰,收縮使下鰓弓靠近尾舌骨。
鰓間腹斜肌(obliquus ventralis, OBV, 曾用名interarcualis ventralis obliqus和下鰓角鰓肌hypoceratobranchialis)有3塊:第1~3鰓間腹斜肌(OBV1~3),分別連接第1~3角鰓骨前端與第1~3下鰓骨外部。該種肌肉收縮使角鰓骨與下鰓骨關節處向上拱起。
腹面橫肌(transversus ventralis, 曾用名transversi ventralis)分為腹面前橫肌(transversus ventralis anterior, TVA)腹面后橫肌(transversus ventralis posterior, TVP) 2部分。前者連接左右第4角鰓骨腹側前部,位于腹面后橫肌前部的下方。后者連接中央軟骨后部與左右第5角鰓骨的內側。腹面橫肌收縮使兩側第4和第5角鰓骨彼此靠近,從而使下鰓弓腹面收緊。
鰓弓收肌(adductor, AD, 曾用名adductor arcuum branchialium)連接成對的角鰓骨和上鰓骨的后端,通常有5部分,而細紋獅子魚的前4部分肌肉退化,僅留第5塊鰓弓收肌,連接第5與第4角鰓骨,并延伸至第4上鰓骨,收縮使第4和第5角鰓骨彼此靠近,并使上下鰓弓后部相靠近。
鰓間后斜肌(obliquus posterior, OBP, 曾用名interarcualis posterior obliqus)位于鰓弓的后部,連接第4上鰓骨和咽鰓骨的背面,至第4和第5角鰓骨的腹面。與鰓弓收肌相鄰,功能上也與其相似。
食道括約肌(sphincter oesophagi, SOE)包裹著鰓弓后側的食道,肌肉束橫向分布,控制食道的收縮擴張。
2.2.5 胸鰭肌肉(pectoral fin muscles)(圖15、圖19)
胸鰭肌肉包括肩帶淺層展肌、肩帶深層展肌、肩帶腹側立肌、肩帶淺層收肌、肩帶深層收肌和肩帶背側立肌。無肩帶橈收肌(adductor radialis)和喙橈肌(coracoradialis)。胸舌肌、胸鰭牽引肌和胸鰭提肌在此描述。脊神經腹枝前3支與側線副神經枝的分支相結合,由上至下伸入胸鰭形成網式結構,并延伸至鰭條上,稱為肩帶神經叢(pectoral plexus,曾用名branchial plexus)。

圖19 胸鰭內側肌肉圖:肌肉全圖(上)和移去肩帶淺層收肌后圖(下)
ADP:肩帶深層收肌;ADS:肩帶淺層收肌;ARD:肩帶背側立肌
ADP: Adductor profundus; ADS: Adductor superficialis; ARD: Arrector dorsalis
肩帶淺層展肌(abductor superficialis, ABS,又名abductor pectoralis superficialis)起始于匙骨外側片狀板的內側,止于除第1胸鰭鰭條外所有鰭條的前部外側,收縮使胸鰭遠離身體,上部往前、下部往前上外翻,鰭條間相靠近。
肩帶深層展肌(abductor profundus, ABP, 又名abductor pectoralis profundus)起始于匙骨外側片狀板的內側和烏喙骨、支鰭骨的外側,止于所有胸鰭鰭條的基部外側,收縮使胸鰭往前下方外翻,可使魚體在水底靜止時維持平衡。
肩帶腹側立肌(arrector ventralis, ARV, 又名肩帶伸肌extensor pectoralis)起始于匙骨外側片狀板內側的上部與肩胛骨的外側,止于第1胸鰭鰭條的前部外側,收縮使鰭條展開,胸鰭上部往前下方外翻。
肩帶淺層收肌(adductor superficialis, ADS,又名adductor pectoralis superficialis)起始于匙骨內側的中部與肩胛骨的內側,止于除第1胸鰭鰭條外所有鰭條的前部內側,收縮使胸鰭往后往內上方收起。
肩帶深層收肌(adductor profundus, ADP, 又名adductor pectoralis profundus)起始于匙骨前部與烏喙骨、支鰭骨的內側,止于所有胸鰭鰭條內側的基部,收縮使胸鰭往后往內收起,與肩帶淺層收肌配合使胸鰭靠近魚體。
肩帶背側立肌(arrector dorsalis, ARD)起始于匙骨與第1支鰭骨間軟骨帶的內側,止于第1胸鰭鰭條前部內側,收縮可展開鰭條,胸鰭上部往前下方內收。在早期的解剖學研究中,通常被作為肩帶深層收肌的一部分進行描述。
胸舌肌(sternohyoideus, STH)是很大的肌肉,起始于匙骨前部的背面以及外側面,并插入匙骨外側凹槽中,止于尾舌骨的外側和后緣。該肌被2個肌隔分為3段,中段肌節的少量肌肉纖維與軸下肌相連,收縮不同部位可調節匙骨的位置方向。
胸鰭提肌(levator pectoralis, LEP)連接腦顱后部與匙骨上端,收縮可使肩帶上部靠近腦顱。
胸鰭牽引肌(protractor pectoralis, PPEC, 又名上耳咽匙肌epioticopharyngoclavicularis、鎖骨提肌levator clavicularis)起始于后耳骨外后側,下方以薄薄的腱膜連接匙骨下部的背緣和上部的前緣,收縮可使肩帶下部向上移動。
2.2.6 腹鰭肌肉(pelvic fin muscles)(圖20) 腹鰭肌肉包括腰帶淺層展肌+腰帶深層展肌復合體、腰帶腹側立肌、腰帶淺層收肌、腰帶深層收肌和腰帶背側立肌。無腰帶固有伸肌(extensor proprius)。前下棱肌和中下棱肌也在此描述。
腰帶淺層展肌(abductor superficialis pelvicus, ABSP, 又名abductor pelvis superficialis)與腰帶深層展肌(abductor profundus pelvicus, ABPP, 又名abductor pelvis profundus)融合形成的復合體(ABSP+ABPP)起始于腰骨內側面下部,經下緣翻出,止點分別位于5條腹鰭鰭條的基部。收縮可使腹鰭鰭條基部上提,末端下降,腹鰭吸盤上拱。

圖20 腹鰭肌背面圖(左)和腹面圖(右)
ABSP+ABPP:腰帶淺層展肌+腰帶深層展肌復合體;ADPP:腰帶深層收肌;ADSP:腰帶淺層收肌;ARDP:腰帶背側立肌;ARVP:腰帶腹側立肌;ICARA:前下棱肌;ICARM:中下棱肌
ABSP+ABPP: Abductor superficialis pelvicus+Abductor profundus pelvicus; ADPP: Adductor profundus pelvicus; ADSP: Adductor superficialis pelvicus; ARDP: Arrector dorsalis pelvicus; ARVP: Arrector ventralis pelvicus; ICARA: Infracarinalis anterior; ICARM: Infracarinalis medius
腰帶腹側立肌(arrector ventralis pelvicus, ARVP, 曾用名腰帶降肌depressor pelvis)起始于下腰骨突的尖端,止于腹鰭鰭棘中后部。收縮使鰭棘向前運動。
腰帶淺層收肌(adductor superficialis pelvicus, ADSP, 又名adductor pelvis superficialis)完全分離為5塊,由前至后依次為分肌1~5。它們的起點由前至后依次排列于腰骨背部的外側,止點分別位于第1~5腹鰭鰭條背面的中部。收縮使腹鰭鰭條基部下降,末端上提,腹鰭吸盤下降。
腰帶深層收肌(adductor profundus pelvicus, ADPP, 又名adductor pelvis profundus)不發達,較小的一束肌肉連接腰骨背部外側的下方與最前方一條腹鰭鰭條的前部。收縮時輔助腰帶淺層收肌的功能。
腰帶背側立肌(arrector dorsalis pelvicus, ARDP, 曾用名腰帶提肌levator pelvis)起始于腰骨擔鰭部背面中縫內側,由上向外翻出后向前延伸,止于腹鰭鰭棘的后部。收縮使鰭棘向后上方運動。
前下棱肌(infracarinalis anterior, ICARA, 又名腹鰭牽引肌protractor pelvis)起始于胸鰭前端匙骨與烏喙骨內側下部,止于腹鰭扇形板外側和內側的下部。其前端與胸舌肌的后部纖維聯合。收縮使腹鰭向前移動。
中下棱肌(infracarinalis medius, ICARM, 又名腹鰭牽縮肌retractor pelvis)起始于腹鰭腰骨的后端,止于肛門往后至臀鰭前方。收縮使腹鰭向后移動。
2.2.7 奇鰭相關肌肉(muscles associated with median fins)(圖21) 背鰭相關肌肉包括背鰭傾肌、背鰭豎肌和背鰭降肌。前上棱肌在此描述。無中、后上棱肌。
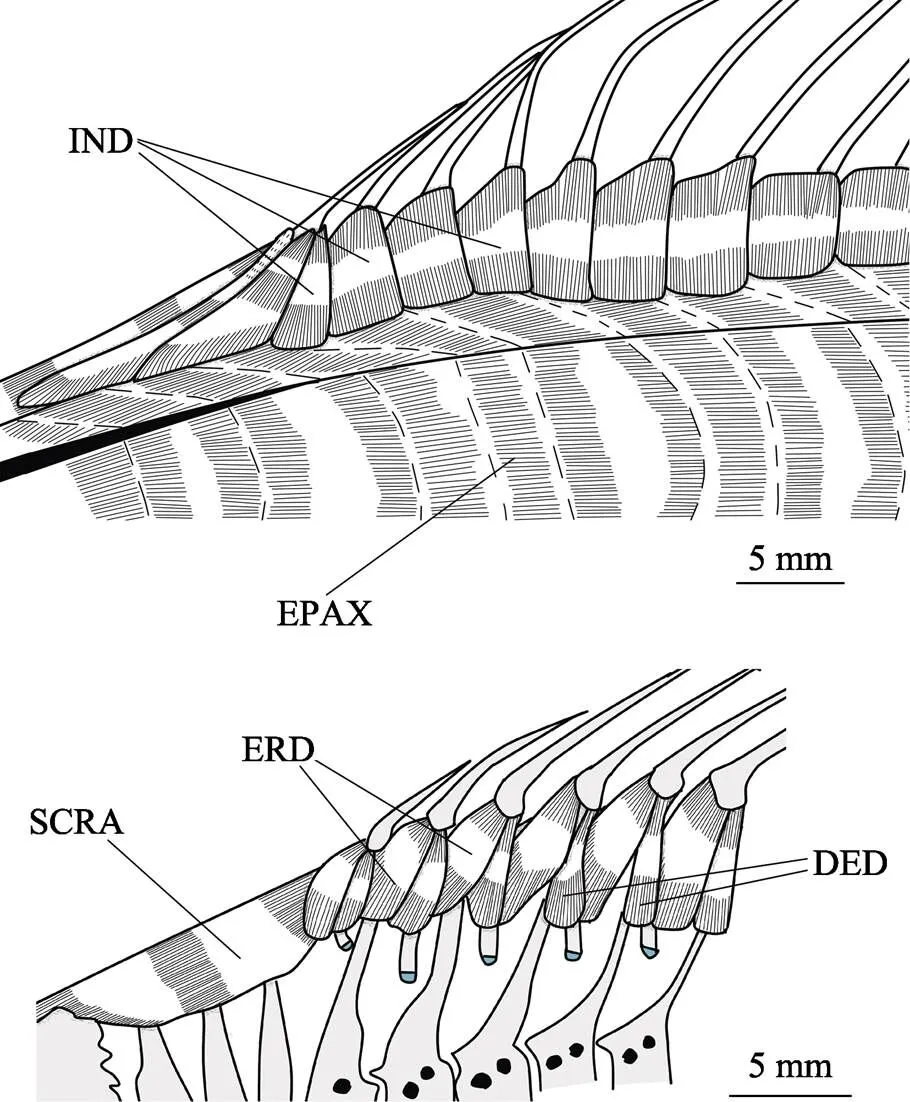
圖21 背鰭肌肉全圖(上)和移除軸上肌圖(下)
DED:背鰭降肌;ERD:背鰭豎肌;EPAX:軸上肌;IND:背鰭傾肌;SCRA:前上棱肌
DED: Depressores dorsales; ERD: Erectores dorsales; EPAX: Epaxialis; IND: Inclinatores dorsales; SCRA: Supracarinalis anterior
背鰭傾肌(inclinatores dorsales, IND)為小片狀肌肉,連于每條背鰭鰭條前部近基部位置,末端在背鰭基部皮膚下的腱膜上,覆蓋軸上肌(epaxialis, EPAX)的上緣,收縮使其附著的鰭條向該側傾倒。
背鰭豎肌(erectores dorsales, ERD)覆蓋于軸上肌之下,對應背鰭鰭條有多塊。它起始于相應基節前面一個髓棘和其周圍腱膜(除了第1塊背鰭豎肌起始于軟骨化基節),止于相應鰭條基部外側。后臨背鰭降肌,收縮使鰭條豎起。
背鰭降肌(depressores dorsales, DED)覆蓋于軸上肌之下,對應背鰭鰭條數有多塊。它起始于相應基節和其周圍腱膜,止于相應鰭條基部外側,收縮使鰭條降下。
前上棱肌(supracarinalis anterior, SCRA, 又名背鰭牽引肌protractor dorsalis)位于魚頭部后上方,連接上枕骨后上端和背鰭第1基節。收縮牽引背鰭向前,使魚背有拱起的趨勢。
臀鰭相關肌肉包括臀鰭傾肌、臀鰭豎肌和臀鰭降肌,結構與背鰭相同。無后下棱肌。
臀鰭傾肌(inclinatores anales)連于每條臀鰭鰭條前部近基部位置,末端在臀鰭基部皮膚下的腱膜上,覆蓋軸下肌(hypaxialis, HPAX)的下緣,收縮使其附著的鰭條向該側傾倒。
臀鰭豎肌(erectores anales)起始于相應基節前部與其前方腱膜,止于相應鰭條基部外側面的前部。后臨臀鰭降肌,收縮使鰭條豎起。
臀鰭降肌(depressores anales)起始于相應基節后部,止于相應鰭條基部外側面的后部,收縮使鰭條降下。
2.2.8 尾鰭肌肉(caudal fin muscles)(圖22) 尾鰭肌肉包括尾鰭下背屈肌、尾鰭上背屈肌、尾鰭上腹屈肌、尾鰭外腹屈肌、尾鰭下腹屈肌、尾鰭腹收肌和尾鰭條間肌。
尾鰭下背屈肌(flexor dorsalis, FD, 又名flexor caudalis dorsalis inferior)起始于上尾下骨板的上緣、尾桿骨后桿和最后一條髓棘后部,止于最上方2條尾鰭主鰭條的前部,收縮使尾鰭上部向該側彎曲。
尾鰭上背屈肌(flexor dorsalis superior, FDS, 又名flexor caudalis dorsalis superior)是尾鰭最上方的肌肉,起始于最后一條髓棘,并覆蓋尾鰭下背屈肌上部,止于背側2條前傾鰭條的前部,收縮使尾鰭上緣向該側下方彎曲。
尾鰭外腹屈肌(flexor ventralis externus, FVE, 又名尾鰭中腹屈肌flexor caudalis ventralis medialis)起始于最后2條脈棘前部和最后一個臀鰭基節,止于第6、第7尾鰭主鰭條的基部,收縮使尾鰭中部鰭條向該側屈轉。

圖22 尾鰭肌肉全圖(上)和移除軸上肌軸下肌后圖(下)
EPAX:軸上肌;FD:尾鰭下背屈肌;FDS:尾鰭上背屈肌;FV:尾鰭上腹屈肌;FVE:尾鰭外腹屈肌;FVI:尾鰭下腹屈肌;HL:尾鰭腹收肌;HPAX:軸下肌;INT:尾鰭條間肌
EPAX: Epaxialis; FD: Flexor dorsalis; FDS: Flexor dorsalis superior; FV: Flexor ventralis; FVE: flexor ventralis externus; FVI: Flexor ventralis inferior; HL: Hypochordal longitudinalis; HPAX: Hypaxialis; INT: Interradialis
尾鰭上腹屈肌(flexor ventralis, FV, 又名flexor caudalis ventralis superior)起始于下尾下骨+準尾下骨復合體,止于第8~10尾鰭主鰭條及腹側前傾鰭條的基部,收縮使尾鰭下部向該側卷曲。
尾鰭下腹屈肌(flexor ventralis inferior, FVI, 又名flexor caudalis ventralis inferior)起始于倒數第1、第2脈棘的后部,止于第10尾鰭主鰭條和腹側前傾鰭條的基部,收縮使尾鰭下緣向該側上方彎曲。
尾鰭腹收肌(hypochordal longitudinalis, HL, 又名adductor caudalis ventralis)起始于下尾下骨+準尾下骨復合體上緣,止于第1~4尾鰭主鰭條的基部,收縮使尾鰭上半部向該側腹面卷曲。
尾鰭條間肌(interradialis, INT, 又名interradialis caudalis)位于尾鰭主鰭條側面前部,連接于10條尾鰭主鰭條之間,從第5、第6鰭條之間分別向上下兩側延伸,上下各5塊,收縮使各鰭條向中軸靠近。
3 討論
3.1 術語與描述
本研究的中文術語主要采用水產類規劃教材中的名詞,但由于國內相關研究較少,大多沿用早期論文研究結果的原因,存在中文術語與現行英文術語不匹配、更新滯后和未定名描述等情況。隨著學科的發展,魚類解剖學的英文名詞更新多次,定義恰當或流傳最廣的被采納。例如,現在國外魚類學家廣泛使用的關節骨和隅骨的英文名分別為anguloarticular和retroarticular,但很多中文相關論文中標注的英文名分別為articular和angular。在20世紀60年代之前,關節骨和隅骨的英文名為articular和angular (Johnson, 1918; Tominaga, 1968),之后逐漸被angular (在此指代關節骨)和retroarticular代替(Fraser, 1968; Yabe, 1985),近年來,使用anguloarticular和retroarticular分別指代關節骨和隅骨成為主流(Datovo, 2012; Vilasri, 2013; Keivany, 2014)。這種英文名詞術語更新的情況也發生在眶下骨、后耳骨、側篩骨、翼蝶骨、翼骨、中翼骨等骨頭以及下頜收肌、鰓蓋開肌、舌肌和鰓間背斜肌等肌肉上。本研究在結果描述中采用的是現今國際廣泛認可的英文名詞術語,也標注了常用的曾用名,可供與早期論文對照。針對某些骨頭或肌肉至今仍存在同物異名的情況,也在結果描述中注明(如中篩骨、前犁骨、上顳骨和前下棱肌等)。部分肌肉在國內文獻中未被定義和描述過,在本研究中按照細紋獅子魚的情況進行了描述,并根據其功能、英文名約定俗成的命名規則確定了中文名(如舌弓展肌和鰓弓背面牽縮肌)。在早期研究中,舌頜骨通常被作為舌弓的一部分進行描述,本研究將舌頜骨作為懸器的組成之一,因為它與懸器部分的關聯性更強,與后翼骨和前鰓蓋骨的接合更緊密,有利于骨骼系統描述的連貫性。
3.2 特殊結構的功能探討
3.2.1 腹鰭吸盤的特殊結構 細紋獅子魚腹鰭的骨骼和肌肉構造與其他科目的魚類(圓鰭魚科除外)具有很大的差異。一般魚類的腹鰭腰骨呈三角狀,以較小的傾斜角度貼于腹部底面內側,大多依靠肌肉、韌帶和其他結締組織連接尾舌骨和肩帶等部位,常有下腰骨突、下腰骨龍骨和后腰骨突等附屬結構,鰭棘呈條狀,鰭條為分叉或條狀形態,主要附著在腰骨的后緣(Yabe, 1985; Vilasri, 2013; Kang, 2018)(圖5)。而細紋獅子魚的腰骨從腹側看則更趨向于橢圓形,背側的扇形板以軟骨帶與肩帶骨內側關聯,加上結締組織和肌肉的連接,使腹鰭與身體的關系更牢固,在不喪失活動能力的前提下,運動穩定性也更高;鰭棘呈縫合針狀,附著與下腰骨突后部,鰭條特化,末端呈半筒狀舌形結構,輻射式附著于腰骨腹側面,構成了吸盤的基本框架。一般魚類腹鰭的肌肉包括腰帶深層展肌、腰帶淺層展肌、腰帶腹側立肌、腰帶淺層收肌、腰帶深層收肌和腰帶背側立肌,縱向附著于背腹兩側(Sasaki, 1989; Winterbottom, 1993; Vilasri, 2013),背側肌肉收縮使其腹鰭鰭條向上收起貼向身體,腹側面肌肉收縮使其腹鰭鰭條向下張開遠離身體,從而達到腹鰭的收張運動。而細紋獅子魚腹鰭肌肉并不縱向排列,它的腰帶深層收肌退化,腰帶淺層展肌+腰帶深層展肌復合體和腰帶淺層收肌分別分化為5個分肌,近似輻射式連于5個腹鰭鰭條和腰骨之間,加上腰帶腹側立肌和腰帶背側立肌的作用,共同帶動盤狀腹鰭進行上拱和下凸的動作,結合外側褶皺狀皮膚和結締組織,使腹鰭具有吸盤的功能。通過該器官,細紋獅子魚能吸附在巖礁上面,有利于在流水中固定身體,因此,腹鰭吸盤是其適應生活環境的重要器官(金鑫波, 2006)。
3.2.2 頰肌的分化 魚類頰肌在一定程度上是研究系統進化和親緣關系時易于掌握的指標。其分肌的數量、形態和種類多種多樣,有的只有1塊下頜收肌,有的卻有10塊明顯的分肌(Winterbottom, 1974)。一般低等鱸形目魚類有3~5塊分肌(Kang, 2018),鲉形目魚類有2~5塊分肌(Yabe, 1985; Imamura, 1996)。細紋獅子魚的頰肌分化為6塊且較發達,有使頜骨向后方移動的A1α、使頜骨向后上方移動的A1β、使上頜骨向內向上運動的A1β′、使下頜骨向后上方移動的A2、使下頜骨向內向上移動的A3以及協同其他分肌將口閉合的Aω。這種分化相較于許多魚類來說更為復雜,能夠更靈活地控制頜骨的運動,使咬合動作更加精準、力度更強,從而使細紋獅子魚能更好地捕食魚蝦等生物,成為生態系統中的頂級捕食者(張波等, 2011)。
3.3 系統進化的問題
獅子魚科是種類最多、分布最廣的魚類類群之一,其成員在形態、分布和生活習性等方面千差萬別,至今仍有新物種不斷被發現(Wang, 2019),該類群內的分類學研究以及系統進化關系十分值得探討。不少學者采用比較骨學或分子生物學手段對獅子魚進行系統進化分析(Kido, 1988; Knudsen, 2007),但包含肌肉性狀分析的比較形態學研究尚無報道,并且細紋獅子魚的親緣關系也不清楚。要得到具體而可靠的細紋獅子魚的親緣關系以及更為詳細的獅子魚科魚類的系統進化關系,進一步的比較解剖學研究是十分必要的。
致謝:感謝日本北海道大學水產學部海洋生物學實驗室魚類分類學家Hisashi Imamura教授給與的寶貴意見。
CHEN Y X, LIU J, LIU L. Comparative osteology in eight sparid fishes (Osteichthyes: Perciformes) with remarks on their classification. Journal of Fisheries of China, 2014, 38(9): 1360–1374 [陳詠霞, 劉靜, 劉龍. 中國鯛科魚類骨骼系統比較及屬種間分類地位探討. 水產學報, 2014, 38(9): 1360–1374]
CHERNOVA N V. Systematics and phylogeny of fish of the genus(Liparidae, Scorpaeniformes). Journal of Ichthyology, 2008, 48(10): 831–852
DATOVO A, CASTRO R M C. Anatomy and evolution of the mandibular, hyopalatine, and opercular muscles in characiform fishes (Teleostei: Ostariophysi). Zoology (Jena, Germany), 2012, 115(2): 84–116
ENDO H, TAKAGI Y. Myology. In: YABE M, KUWAMURA T, TAKAGI Y. Ichthyology. Tokyo: Kouseisha Kouseikaku Company Limited, 2017, 377
FRASER T H. Comparative osteology of the Atlantic snooks (Pisces:). Copeia, 1968, 1968(3): 433–460
FREIHOFER W C. Cranial nerves of a percoid fish,(family Nandidae), a contribution to the morphology and classification of the order Perciformes. Occasional Papers of the California Academy of Sciences, 1978, 128: 78
IMAMURA H. Phylogeny of the family Platycephalidae and related taxa (Pisces: Scorpaeniformes). Species Diversity, 1996, 1(2): 123–233
JIN X B. Fauna Sinica: Osteichthyes Scorpaeniformes. Beijing: Science Press, 2006, 739 [金鑫波. 中國動物志: 硬骨魚綱鲉形目. 北京: 科學出版社, 2006, 739]
JIN X S, TANG Q S. Changes in fish species diversity and dominant species composition in the Yellow Sea. Fisheries Research, 1996, 26(3/4): 337–352
JOHNSON S E. Osteology of the gruntfish,. Journal of Morphology, 1918, 31(3): 461–487
KANG S, IMAMURA H, KAWAI T. Morphological evidence supporting the monophyly of the family Polynemidae (Teleostei: Perciformes) and its sister relationship with Sciaenidae. Ichthyological Research, 2018, 65(1): 29–41
KAWAI T. Osteology. In: YABE M, KUWAMURA T, TAKAGI Y. Ichthyology. Tokyo: Kouseisha Kouseikaku Company Limited, 2017, 377
KEIVANY Y. Comparative osteology of the jaws in representativesof the Eurypterygian fishes. Research in Zoology, 2014, 4(2): 29–42
KIDO K. Phylogeny of the family Liparidae, with the taxonomy of the species found around Japan. Memoirs of the Faculty of Fisheries Hokkaido University, 1988, 35(2): 125–256
KNUDSEN S W, M?LLER P R, GRAVLUND P. Phylogeny of snailfishes (Teleostei: Liparidae) based on molecular and morphological data. Molecular Phylogenetics and Evolution, 2007, 44(2): 649–666
MENG Q W, MIAO X S, YU Q J,. Ichthyology: Morphology and classification. Shanghai: Shanghai Science and Technology Press, 1994, 298 [孟慶聞, 繆學祖, 俞秦濟, 等. 魚類學: 形態分類. 上海: 上海科學技術出版社, 1994, 298]
MYOUNG S H, BAN T W, KIM J K. Population structure of(PISCES, Liparidae) from Korea based on morphological and molecular traits. Korean Journal of Fisheries and Aquatic Sciences, 2016, 49(2): 198–207
NAKABO T. Fishes of Japan with pictorial keys to the species, English edition. Tokyo: Tokai University Press, 2002, 1749
PING C, PAO S, CHEN J S. A preliminary observation of the gross anatomy of the nerve system of the carp (). Acta Zoologica Sinica, 1959, 11(3): 370–389 [秉志, 鮑璿, 陳進生. 鯉魚神經系統的初步觀察. 動物學報, 1959, 11(3): 370–389]
PING C, PAO S, YANG H Y. The skeletal musculature of the carp (). Acta Zoologica Sinica, 1958, 10(3): 289–315 [秉志, 鮑璿, 楊慧一. 鯉魚骨骼肌的初步觀察. 動物學報, 1958, 10(3): 289–315]
SASAKI K. Phylogeny of the family Sciaenidae, with notes on its zoogeography (Teleostei, Perciformes). Memoirs of the Faculty of Fisheries Sciences, Hokkaido University, 1989, 36: 1–137
SHAN X J, CHEN Y L, ZHOU Z P,. Inter-annual variations in propagational biological characteristics ofin central and southern Yellow Sea. Progress in Fishery Sciences, 2014, 35(3): 1–8 [單秀娟, 陳云龍, 周志鵬, 等. 黃海中南部細紋獅子魚繁殖生物學特征的年際變化. 漁業科學進展, 2014, 35(3): 1–8]
SHUI B N, ZHANG S L, HAN Z Q,. Ichthyological systematics. Beijing: Ocean Press, 2019, 462 [水柏年, 張盛龍, 韓志強, 等. 系統魚類學. 北京: 海洋出版社, 2019, 462]
TOMINAGA Y. Internal morphology, mutual relationships and systematic position of the fishes belonging to the family Pempheridae. Japanese Journal of Ichthyology, 1968, 15(2): 43–95
VILASRI V. Comparative anatomy and phylogenetic systematics of the family Uranoscopidae (Actinopterygii: Perciformes). Memoirs of the Faculty of Fisheries Sciences, Hokkaido University, 2013, 55(1/2): 1–106
WANG K, SHEN Y, YANG Y,. Morphology and genome of a snailfish from the Mariana Trench provide insights into deep-sea adaptation. Nature Ecology and Evolution, 2019, 3(5): 823–833
WINTERBOTTOM R. A descriptive synonymy of the striated muscles of the Teleostei. Proceedings of the Academy of Natural Sciences of Philadelphia, 1974, 125: 235–317
WINTERBOTTOM R. Myological evidence for the phylogeny of recent genera of surgeonfishes (Percomorpha, Acanthuridae), with comments on the Acanthuroidei. Copeia, 1993, 1993(1): 21–39
YABE M. Comparative osteology and myology of the superfamily Cottoidea (Pisces: Scorpaeniformes), its phyogenetic classification. Memoirs of the Faculty of Fisheries Sciences, Hokkaido University, 1985, 32: 1–130
YANG T Y, LIU F, LI Y J,. Comparison of skeletal characteristics of four fish species in Stromateoidei. Fisheries Science, 2014, 33(10): 643–648 [楊太有, 劉芳, 李艷潔, 等. 鯧亞目4種魚類骨骼系統的比較研究. 水產科學, 2014, 33(10): 643–648]
ZHANG B, JIN X S, DAI F Q. Feeding habits and their variation of seasnail () in the central and southern Yellow Sea. Journal of Fisheries of China, 2011, 35(8): 1199–1207 [張波, 金顯仕, 戴芳群. 黃海中南部細紋獅子魚的攝食習性及其變化. 水產學報, 2011, 35(8): 1199–1207]
Osteological and Myological Characteristics offrom Chinese Coasts
KANG Shihua1, JIN Xianshi1,2, WANG Jun1,3①
(1. Yellow Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Key Laboratory of Sustainable Development of Marine Fisheries, Ministry of Agriculture and Rural affairs, Shandong Provincial Key Laboratory of Fishery Resources and Ecological Environment, Qingdao, Shandong 266071, China; 2. Laboratory for Marine Fisheries Science and Food Production Processes, Pilot National Laboratory for Marine Science and Technology (Qingdao), Qingdao, Shandong 266071, China; 3. Laboratory for Marine Ecology and Environmental Science, Pilot National Laboratory for Marine Science and Technology (Qingdao), Qingdao, Shandong 266237, China)
Fish anatomy, including osteology and myology, is very important for traditional ichthyological research and systematics. It provides the basis for taxonomy, phylogeny, morphology, veterinary, evolution, and other studies. However, few related papers on fish anatomy have been published in China, and hence, research is currently insufficient., is a dominant species in the Yellow Sea and Bo Hai, which occupies an important position in the ecosystem, but no study has so far described its anatomical characteristics in detail. The osteological and myological descriptions ofare given in this study, and some associated nerves are also mentioned here. The osteological descriptions comprise ten parts: circumorbital bones, neurocranium, jaws, suspensorium and opercular bones, hyoid arch, branchial arches, pectoral girdle, pelvic girdle, axial skeleton, median fin supports, and caudal skeleton; and myological descriptions comprise eight parts: cheek muscles, cephalic muscles between the cranium and suspensorium-opercular bones, ventral muscles of the head, branchial muscles, pectoral fin muscles, pelvic fin muscles, muscles associated with median fins, and caudal fin muscles. Examinations were made on specimens stained with alcian blue and alizarin red. The specimens were dissected under a stereomicroscope used to prepare drawings. Some renewal conditions of terminology and synonyms were stated, and few portions unnamed in Chinese, such as hyohyoidei abductor and retractor dorsalis, were named. The special structure of the pelvic fin and its locomotory mechanism were illustrated, along with the differentiation and function of the cheek muscles.
Osteology; Myology;;Morphology; Fish anatomy
WANG Jun, E-mail: wangjun@ysfri.ac.cn
Q954.54
A
2095-9869(2022)02-0062-18
10.19663/j.issn2095-9869.20201013001
* 中國水產科學研究院中央級公益性科研院所基本科研業務費(2020TD19)資助[This work was supported by Central Public-Interest Scientific Institute Basal Research Fund, CAFS (2020TD19)]. 亢世華,E-mail: kangsh@ysfri.ac.cn
王 俊,研究員,E-mail: wangjun@ysfri.ac.cn
2020-10-13,
2020-12-19
亢世華, 金顯仕, 王俊. 中國產細紋獅子魚的骨學和肌肉學研究. 漁業科學進展, 2022, 43(2): 62–79
KANG S H, JIN X S, WANG J. Osteological and myological characteristics offrom Chinese coasts. Progress in Fishery Sciences, 2022, 43(2): 62–79
(編輯 馬璀艷)
