橘色雙冠麗魚體色相關(guān)基因mitf的結(jié)構(gòu)及表達(dá)調(diào)控特性*
2022-04-11 06:53:24陳旭東鄔國強(qiáng)宋紅梅汪學(xué)杰牟希東胡隱昌
漁業(yè)科學(xué)進(jìn)展 2022年2期
陳旭東 鄔國強(qiáng) 宋紅梅 汪學(xué)杰 牟希東 劉 奕 劉 超 胡隱昌
橘色雙冠麗魚體色相關(guān)基因的結(jié)構(gòu)及表達(dá)調(diào)控特性*
陳旭東1,2鄔國強(qiáng)1,2宋紅梅2①汪學(xué)杰2牟希東2劉 奕2劉 超2胡隱昌2
(1. 上海海洋大學(xué) 水產(chǎn)科學(xué)國家級(jí)實(shí)驗(yàn)教學(xué)示范中心 上海 201306;2. 中國水產(chǎn)科學(xué)研究院珠江水產(chǎn)研究所農(nóng)業(yè)農(nóng)村部休閑漁業(yè)重點(diǎn)實(shí)驗(yàn)室 廣東省水產(chǎn)動(dòng)物免疫技術(shù)重點(diǎn)實(shí)驗(yàn)室 廣東 廣州 510380)
為了解小眼畸形相關(guān)轉(zhuǎn)錄因子基因在魚類早期體色褪黑過程中的調(diào)控作用,本研究采用cDNA末端快速擴(kuò)增(RACE)技術(shù)獲得橘色雙冠麗魚()基因cDNA序列全長(zhǎng),利用實(shí)時(shí)熒光定量PCR (qRT-PCR)技術(shù)檢測(cè)其在橘色雙冠麗魚胚胎不同發(fā)育時(shí)期、體色褪黑不同時(shí)期和各個(gè)組織中的表達(dá)規(guī)律。獲得基因2個(gè)亞型,其中,的cDNA全長(zhǎng)為1816 bp,包括5′非編碼區(qū)(UTR) 158 bp、3′UTR 428 bp、開放閱讀框(ORF) 1230 bp,共編碼409個(gè)氨基酸;的cDNA全長(zhǎng)為1638 bp,包括5′UTR 160 bp、3′UTR 428 bp和ORF 1050 bp,共編碼349個(gè)氨基酸。同源性和系統(tǒng)進(jìn)化分析顯示,和聚在一小支,與慈鯛科(Cichlidae)魚類同源性最高,與哺乳類動(dòng)物同源性較低。qRT-PCR結(jié)果顯示,在成魚各個(gè)組織中,和均有不同程度表達(dá),其中,眼部表達(dá)量最高且顯著高于其他組織(<0.05),肌肉、腦和腎臟也有較高表達(dá);和在胚胎各個(gè)發(fā)育時(shí)期均有表達(dá),在受精卵時(shí)期表達(dá)量最高,顯著高于其他胚胎期;隨著橘色雙冠麗魚體色由黑色過渡到橘黃色,和在魚皮膚、鱗片、尾鰭中的表達(dá)均呈逐漸下降的趨勢(shì),表明基因表達(dá)與魚體色由黑到黃轉(zhuǎn)變的表型間存在關(guān)聯(lián)性,推測(cè)與魚體色發(fā)育階段色素細(xì)胞的分化和分布比例的動(dòng)態(tài)變化相關(guān)。本研究通過了解魚類體色發(fā)育和變異的分子基礎(chǔ),可為魚類色素細(xì)胞發(fā)育和體色人工改良積累資料。
橘色雙冠麗魚;小眼畸形相關(guān)轉(zhuǎn)錄因子(MITF);qRT-PCR;黑色素細(xì)胞
MITF (microphthalmia-associated transcription factor)即小眼畸形相關(guān)轉(zhuǎn)錄因子,于放射誘導(dǎo)的突變小鼠中發(fā)現(xiàn)(Hertwig, 1942),突變體的后代會(huì)出現(xiàn)小眼畸形、早發(fā)性耳聾、皮毛和虹膜色素減退等癥狀。MITF是動(dòng)物皮膚、眼睛和羽毛色素中黑色素細(xì)胞發(fā)育的關(guān)鍵調(diào)控因子(Lin, 2019),具有基本–螺旋–環(huán)–亮氨酸拉鏈(basic helix-loop-helix-leucine zipper, bHLHZip)結(jié)構(gòu),以二聚體的形式與酪氨酸家族啟動(dòng)子Mbox高度結(jié)合,通過直接調(diào)控、、、和等基因的表達(dá),從而在黑色素細(xì)胞的發(fā)育、存活、遷移、增殖和分化過程中發(fā)揮重要作用(Steingrimsson, 2004)。在哺乳動(dòng)物中,基因的突變或缺失可能會(huì)導(dǎo)致動(dòng)物耳朵、眼睛、毛發(fā)、體色等表型發(fā)生改變,如鵪鶉()白色羽毛的產(chǎn)生(Minvielle, 2010)、馬() (Hauswirth, 2019)和犬() (Korberg, 2014)皮毛白色斑點(diǎn)的出現(xiàn)等。在魚類中,作為治療黑色素瘤潛在的靶基因,已在斑馬魚()中開展了大量研究(Lister, 2014),在青鳉() (Li, 2013)、錦鯉() (Liu, 2015)、鯽魚(, red var.) (Zhang, 2017)等魚類中也開展了在胚胎和各個(gè)成魚組織中表達(dá)情況的研究,初步探索了在魚類體色形成過程中的作用,但其具體的調(diào)控機(jī)制及與其他體色相關(guān)基因的聯(lián)級(jí)作用仍未被闡明。
橘色雙冠麗魚(),俗稱紅魔鬼,原產(chǎn)于中南美洲的尼加拉瓜、哥斯達(dá)黎加等地,是一種既可食用又可觀賞的大型熱帶魚類(Barluenga, 2010; Kautt, 2012)。通常1齡可達(dá)性成熟,成熟后的麗魚體色為橘紅色或橘色,生產(chǎn)上常將其作為父本與紅頭麗魚()雜交,產(chǎn)生更具觀賞性的子一代血鸚鵡魚(♂♀)。橘色雙冠麗魚在發(fā)育過程中會(huì)出現(xiàn)體色過渡的現(xiàn)象,體色由最初黑色過渡至灰色,灰色再過渡至亮黃色,這一現(xiàn)象稱為體色褪黑(蔣燕玲, 2016),主要是由于色素細(xì)胞的形成、增殖、遷移和分化所致。橘色雙冠麗魚含4種色素細(xì)胞,包括黑色素細(xì)胞、黃色素細(xì)胞、紅色素細(xì)胞和虹彩細(xì)胞,其中,紅色素細(xì)胞和虹彩細(xì)胞只在特定部位分布且數(shù)量較少,黑色素細(xì)胞和黃色素細(xì)胞在“黑色–灰色–黃色”3個(gè)時(shí)期均有分布,孵化后至體色由黑到黃的轉(zhuǎn)變過程中,黑色素細(xì)胞數(shù)量呈逐漸增加而后又減少的趨勢(shì),黃色素細(xì)胞數(shù)量則呈一直增加趨勢(shì)(韋敏俠等, 2015)。目前,國內(nèi)外對(duì)魚類早期體色褪黑這一復(fù)雜生物學(xué)過程的研究相對(duì)較少,其調(diào)控機(jī)制仍不明確。
魚類體色的形成和分布主要是由其體表鱗片和皮膚中色素細(xì)胞的類型、分布和數(shù)量所決定(Shi, 2015; Yu, 2012),目前,已在魚類中鑒定出6種色素細(xì)胞,包括黑色素細(xì)胞、黃色素細(xì)胞、紅色素細(xì)胞、虹彩細(xì)胞、白色素細(xì)胞和藍(lán)色素細(xì)胞(Volkening, 2018)。其中,黑色素細(xì)胞分布最為廣泛,含有大量的黑色素顆粒,能夠吸收特定波長(zhǎng)的入射光,使魚的顏色呈現(xiàn)黑色/灰色(Zhang, 2017)。黑色素細(xì)胞起源于外胚層神經(jīng)嵴細(xì)胞,由神經(jīng)嵴細(xì)胞經(jīng)黑素母細(xì)胞、黑素干細(xì)胞發(fā)育成黑色素細(xì)胞(Cohen, 2016),黑色素細(xì)胞的形成受一系列通路和基因的嚴(yán)格調(diào)控(Hou, 2008),其中,MC1R/α-MSH信號(hào)通路(Newton, 2007)、PI3K/Akt信號(hào)通路(Khaled, 2002)、MAPK信號(hào)通路(Wang, 2017)、WNT/ β-catenin信號(hào)通路(Yamada, 2010)、NO信號(hào)通路(Park, 2009)為最常見的5條信號(hào)通路,作為這5條通路共有的靶向基因,直接關(guān)聯(lián)黑色素細(xì)胞發(fā)育所必需的多個(gè)基因的表達(dá),包括和等,對(duì)黑色素細(xì)胞存活、遷移、增殖和分化起著關(guān)鍵性作用(Steingrimsson, 2004)。本研究擬聚焦黑色素合成關(guān)鍵基因,檢測(cè)其在橘色雙冠麗魚各組織、胚胎發(fā)育各時(shí)期和體色褪黑轉(zhuǎn)換期的表達(dá)模式,了解其在魚類體色褪黑調(diào)控中的作用規(guī)律。
1 材料與方法
1.1 實(shí)驗(yàn)材料
橘色雙冠麗魚取自中國水產(chǎn)科學(xué)研究院珠江水產(chǎn)研究所。選取性成熟且體色為橘色的健康雙親進(jìn)行配對(duì),獲取不同發(fā)育時(shí)期的胚胎,包括受精卵、卵裂期、原腸期、神經(jīng)期、視泡期、聽泡期、心臟形成期、血液循環(huán)期等8個(gè)時(shí)期胚胎(蔣燕玲, 2016)。在體色發(fā)育的3個(gè)褪色階段“黑色、灰白、黃色”各取3尾魚,剝離鱗片、皮膚和尾鰭。同樣選取性成熟且體色為淡黃色的橘色雙冠麗魚3尾,分離鰓、腦、肌肉、性腺、眼、腎和心臟7個(gè)組織,均用Trizol處理,于–80℃冰箱保存,用于RNA提取和熒光定量分析。
1.2 實(shí)驗(yàn)方法
1.2.1基因cDNA全長(zhǎng)克隆 取性成熟橘色雙冠麗魚皮膚、鱗片、尾鰭于Trizol中,按Tissue RNA kit (OMEGA)操作步驟提取總RNA,通過1%瓊脂糖凝膠電泳和SynergyTMNEO HTS多功能酶標(biāo)儀檢測(cè)RNA的完整性、純度及濃度。利用Prime ScriptTMⅡ 1st strand cDNA synthesis kit (TaKaRa)反轉(zhuǎn)錄試劑盒合成cDNA第一鏈,置于–20℃冰箱保存。
根據(jù)已知慈鯛科(Cichlidae)魚類保守序列,利用Primer 5.0軟件設(shè)計(jì)擴(kuò)增引物-F和-R (表1),以cDNA第一鏈為模板,擴(kuò)增目的基因片段。25 μL擴(kuò)增體系:16.75 μL ddH2O,4 μL dNTP mix,2.5 μL 10×Buffer,0.5 μL模板,上下游引物各0.5 μL,0.25μL r酶。反應(yīng)條件:94℃預(yù)變性3 min,94℃變性30s,55℃退火60 s,72℃延伸60 s,35個(gè)循環(huán),72℃后延伸5 min。PCR產(chǎn)物經(jīng)1.0%瓊脂糖凝膠電泳檢測(cè)后,送廣州艾基生物有限公司測(cè)序。根據(jù)測(cè)序結(jié)果設(shè)計(jì)5′race和3′race巢氏引物(表1),通過Smarter Race 5′/3′ kit components (TaKaRa)進(jìn)行5′末端和3′末端序列擴(kuò)增。擴(kuò)增后PCR產(chǎn)物進(jìn)行膠回收、連菌、37℃培養(yǎng)過夜,挑選陽性克隆進(jìn)行測(cè)序。利用軟件Vector NTI將5′端、3′端及其中間序列拼接,從而獲得橘色雙冠麗魚完整cDNA序列(圖1和圖2)。
1.2.2 生物信息學(xué)分析 將拼接好的橘色雙冠麗魚基因在NCBI數(shù)據(jù)庫中通過blast功能進(jìn)行同源性分析,利用ORF finder查找開放閱讀框;采用ExPASy ProtParam分析MITF蛋白的理化性質(zhì),包括蛋白質(zhì)分子量、理論等電點(diǎn)、親水系數(shù)等;借助Signal IP 4.1進(jìn)行信號(hào)肽預(yù)測(cè);利用TMHMM 2.0查找跨膜結(jié)構(gòu)域;使用NetPhos 3.1工具進(jìn)行磷酸化位點(diǎn)的查找;通過NetNGlyc 4.0查找糖基化位點(diǎn),借助InterPro進(jìn)行氨基酸結(jié)構(gòu)域的分析;使用SOPAMA預(yù)測(cè)蛋白質(zhì)二級(jí)結(jié)構(gòu);利用MEGA 7軟件構(gòu)建系統(tǒng)進(jìn)化樹。
1.2.3基因熒光定量分析 通過RNA提取試劑盒提取各胚胎、組織樣品RNA,利用瓊脂糖凝膠電泳和酶標(biāo)儀檢測(cè)其完整性及濃度。取1 ng RNA用EVOM-MLV反轉(zhuǎn)錄試劑盒合成cDNA第一鏈,存于–20℃冰箱待用。根據(jù)、基因全長(zhǎng)設(shè)計(jì)特異性引物(表1),以β-actin為內(nèi)參基因,采用Real-Time PCR方法在QuantStudio6 Flex儀器上對(duì)橘色雙冠麗魚各時(shí)期胚胎和不同組織進(jìn)行熒光定量分析。20 μL反應(yīng)體系:SYBR Green master mix 10 μL,ddH2O 8μL,cDNA 1 μL,上下游引物(10 μmol/L)各0.5 μL。反應(yīng)條件:50℃ 2 min,95℃ 5 min;95℃變性15 s,55℃退火30 s,72℃延伸30 s,40個(gè)循環(huán),每個(gè)樣品3個(gè)重復(fù)。基因相對(duì)表達(dá)量的具體計(jì)算公式參考蔣燕玲等(2016)。利用SPSS 22.0軟件進(jìn)行單因素方差分析(one-way ANOVA)和Duncan多重比較。
2 結(jié)果
2.1 mitf基因cDNA全長(zhǎng)和氨基酸序列分析
獲得橘色雙冠麗魚基因2個(gè)亞型和,其中,cDNA全長(zhǎng)為1816 bp,5′UTR長(zhǎng)158 bp,3′UTR長(zhǎng)428 bp,開放閱讀框(ORF)為1230 bp,共編碼409個(gè)氨基酸。預(yù)測(cè)其蛋白質(zhì)分子量為39.1 kDa,理論等電點(diǎn)為5.23,親水性系數(shù)為–0.603。TMHMM2.0查找發(fā)現(xiàn)無跨膜結(jié)構(gòu)。磷酸化位點(diǎn)分析顯示,該蛋白包含27個(gè)絲氨酸(S)磷酸化位點(diǎn)、9個(gè)蘇氨酸(T)磷酸化位點(diǎn)、5個(gè)酪氨酸(Y)磷酸化位點(diǎn)。使用SOPAMA預(yù)測(cè)蛋白二級(jí)結(jié)構(gòu),其中,α螺旋結(jié)構(gòu)131個(gè)(占37.01%),β折疊22個(gè)(占6.21%),無規(guī)則卷曲結(jié)構(gòu)177個(gè)(占50%),延展鏈結(jié)構(gòu)24個(gè)(占6.78%)。
表1 引物序列

Tab.1 Primer sequences

圖1 橘色雙冠麗魚mitf1基因cDNA全長(zhǎng)及氨基酸序列
方框?yàn)槠鹗济艽a子ATG;*代表終止密碼子;灰色陰影區(qū)域代表bHLHzip結(jié)構(gòu)域。下同
Grey box is the initiation codon; “*” is the termination codon; Grey shade is bHLHzip domain. The same as below

圖2 橘色雙冠麗魚mitf2基因cDNA全長(zhǎng)及氨基酸序列

圖3 橘色雙冠麗魚mitf1、mitf2基因的氨基酸序列比對(duì)

圖4 橘色雙冠麗魚MITF1、MITF2蛋白二級(jí)結(jié)構(gòu)預(yù)測(cè)
A:MITF1;B:MITF2;h:α螺旋;e:延展鏈;c:無規(guī)則卷曲;t:β折疊;灰色方框?yàn)镸ITF1、MITF2蛋白二級(jí)結(jié)構(gòu)的相同部分
A: MITF1; B: MITF2; h: Alpha helix; e: Extended strand; c: Random coil; t: Beta turn; Grey boxed are the consensus of the secondary structure of MITF1 and MITF2 proteins
的cDNA全長(zhǎng)為1638 bp,5′UTR長(zhǎng)160 bp,3′UTR長(zhǎng)428 bp,ORF為1050 bp,共編碼349個(gè)氨基酸。預(yù)測(cè)其蛋白質(zhì)分子量為38.5 kDa,理論等電點(diǎn)為5.23,親水系數(shù)為–0.645和氨基酸序列比對(duì)結(jié)果顯示(圖3)亞型氨基酸序列與亞型相比,在0~53的位置多53個(gè)氨基酸,82~86的位置多5個(gè)氨基酸,其余位置完全相同。TMHMM 2.0查找發(fā)現(xiàn)無跨膜結(jié)構(gòu)。磷酸化位點(diǎn)分析顯示,該蛋白包含27絲氨酸(S)磷酸化位點(diǎn)、9個(gè)蘇氨酸(T)磷酸化位點(diǎn)、5個(gè)酪氨酸(Y)磷酸化位點(diǎn)。使用SOPAMA預(yù)測(cè)蛋白二級(jí)結(jié)構(gòu)(圖4),其中,α螺旋結(jié)構(gòu)142個(gè)(占40.69%),β折疊14個(gè)(占4.01%),無規(guī)則卷曲結(jié)構(gòu)174個(gè)(占49.86%),延展鏈結(jié)構(gòu)19個(gè)(占5.44%)。與蛋白二級(jí)結(jié)構(gòu)相比,除0~53和82~86位置多出氨基酸及個(gè)別位點(diǎn)結(jié)構(gòu)不同外,二級(jí)結(jié)構(gòu)基本一致。
2.2 mitf基因同源比對(duì)和系統(tǒng)進(jìn)化樹分析
橘色雙冠麗魚2個(gè)亞型和的bHLHzip結(jié)構(gòu)氨基酸序列與大多數(shù)物種高度一致,表明bHLHzip結(jié)構(gòu)在進(jìn)化過程中較為保守。利用MegAlign軟件分析橘色雙冠麗魚mitf氨基酸序列與其他物種的同源性(表2),結(jié)果顯示,橘色雙冠麗魚氨基酸序列與斑馬擬麗魚()、尼羅羅非魚()和薩伊藍(lán)六間()等魚類同源性最高,分別為90.7%、91.9%和93%,與其他硬骨魚類如大黃魚()、斑魚(s)和孔雀魚()也有較高的同源性,分別達(dá)到了85.6%、87.4%和82.1%,而與人()、黑猩猩()、雞()和非洲爪蟾()則相對(duì)較低,只有60%、60.9%、58.9%和63.1%。橘色雙冠麗魚氨基酸序列與人、黑猩猩、雞和非洲爪蟾同源性同源也相對(duì)較低,分別為59.8%、60.9%、58.5%和64.4%,而與尼加拉瓜湖始麗魚()、伯氏樸麗魚()、布氏新亮麗鯛()和盲曹魚()同源性則相對(duì)較高,都達(dá)到了90%以上。
表2氨基酸序列的同源性

Tab.2 GenBank accession number and the identity of mitf1 and mitf2

圖5 MITF1、MITF2蛋白質(zhì)序列系統(tǒng)進(jìn)化樹
以鄰接法(Neighbor-joining)構(gòu)建橘色雙冠麗魚MITF1、MITF2系統(tǒng)進(jìn)化樹(圖5)。進(jìn)化樹分為兩大支,哺乳類、鳥類、爬行類聚為一大支,橘色雙冠麗魚與其他慈鯛科、鮭科、鯉科魚類聚為另一大支。其中,橘色雙冠麗魚MITF1、MITF2和尼加拉瓜湖始麗魚聚為一小支,與斑馬擬麗魚、布氏新亮麗鯛、伯氏樸麗魚、薩伊藍(lán)六間、尼羅羅非魚等魚類親緣關(guān)系最近;而與人、黑猩猩、小鼠、原雞、非洲爪蟾等親緣關(guān)系則相對(duì)較遠(yuǎn)。
2.3 mitf基因的表達(dá)差異分析
2.3.1 胚胎不同發(fā)育時(shí)期中的基因表達(dá)在8個(gè)胚胎發(fā)育時(shí)期均有不同程度的表達(dá),其中,受精卵時(shí)期表達(dá)量最高,顯著高于其他7個(gè)時(shí)期(<0.05),隨著胚胎的發(fā)育,表達(dá)量逐漸下降,在神經(jīng)期和視泡期只有微量表達(dá)。胚胎在發(fā)育至聽泡期時(shí)表達(dá)量驟然上升(<0.05),隨后在心臟形成期和血液循環(huán)期又呈現(xiàn)下降趨勢(shì)(圖6);在橘色雙冠麗魚胚胎發(fā)育的8個(gè)時(shí)期也均有表達(dá),在受精卵時(shí)期表達(dá)量最高,顯著高于其他發(fā)育時(shí)期(<0.05)。在卵裂期、原腸期表達(dá)量驟然降低,只有微量表達(dá)。發(fā)育至神經(jīng)期時(shí)表達(dá)量升高,直至血液循環(huán)期均呈上升趨勢(shì),表達(dá)量不斷增加(圖7)。

圖6 mitf1在胚胎8個(gè)發(fā)育時(shí)期的表達(dá)差異分析
不同字母代表差異顯著(<0.05)。下同
Different letters show significant difference (<0.05). The same as below

圖7 mitf2在胚胎8個(gè)發(fā)育時(shí)期的表達(dá)差異分析
2.3.2 體色過渡3個(gè)時(shí)期基因的表達(dá)模式 橘色雙冠麗魚在“黑色–灰色–黃色”3個(gè)典型的體色過渡時(shí)期,在皮膚、鱗片、尾鰭,、均有不同程度的表達(dá)(圖8和圖9)。其中,在黑色過渡期皮膚中表達(dá)量最高,顯著高于灰色和黃色過渡期(<0.05),灰色過渡期表達(dá)量略高于黑色過渡期,但無顯著差異,整體呈逐漸下降的趨勢(shì),在鱗片和尾鰭中,的表達(dá)趨勢(shì)與在皮膚中基本一致;在皮膚中,在黑色過渡期表達(dá)量最高,隨著橘色雙冠麗魚發(fā)育,表達(dá)量逐漸降低,但無顯著差異(>0.05),在鱗片和尾鰭中,在3個(gè)過渡時(shí)期的表達(dá)量同樣呈現(xiàn)下降的趨勢(shì),黑色過渡期表達(dá)量最高,顯著高于灰色過渡期和黃色過渡期(<0.05),灰色過渡期表達(dá)量高于黃色過渡期,但無顯著差異(>0.05)。
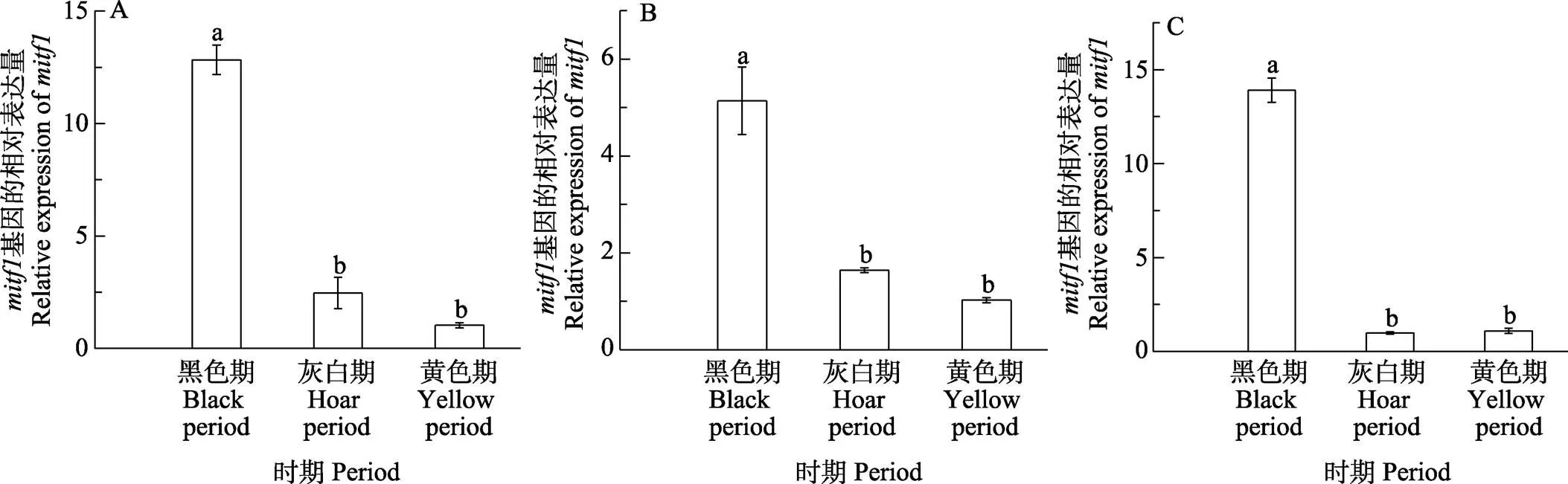
圖8 mitf1在不同體色過渡時(shí)期的表達(dá)差異分析
A:皮膚;B:鱗片;C:尾鰭
A: Skin; B: Scale; C: Tail fin
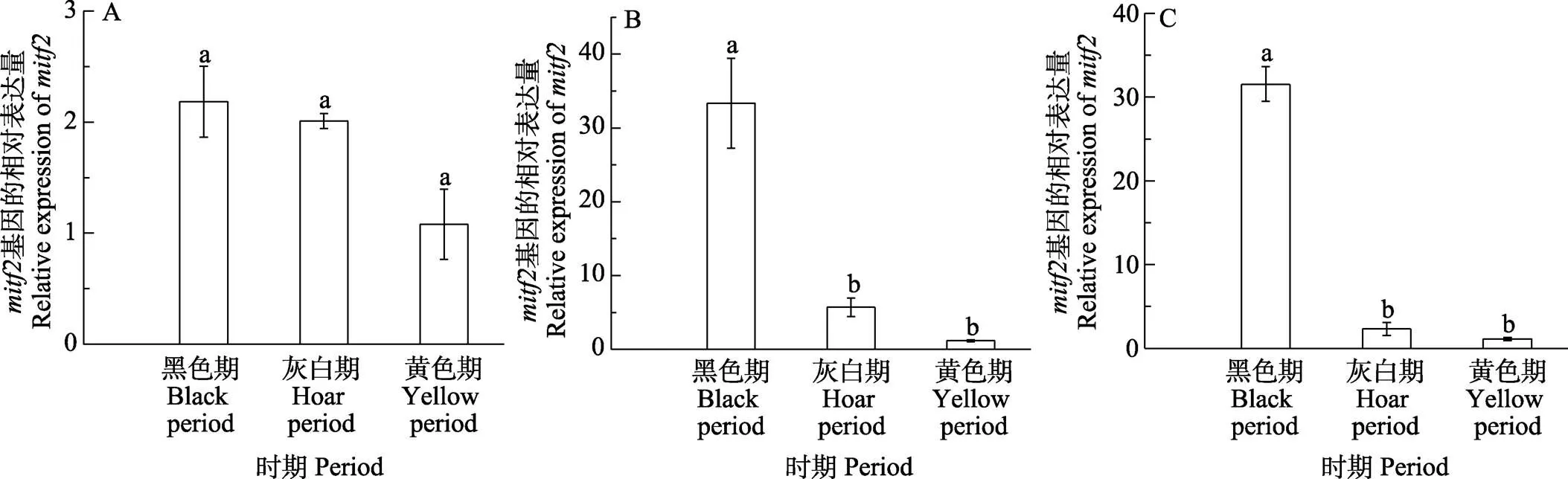
圖9 mitf2在不同體色過渡時(shí)期的表達(dá)差異分析
A:皮膚;B:鱗片;C:尾鰭
A: Skin; B: Scale; C: Tail fin
2.3.3 成魚不同組織中的基因表達(dá)差異 通過實(shí)時(shí)熒光定量PCR (qRT-PCR)檢測(cè)和在橘色雙冠麗魚不同組織中的表達(dá)差異(圖10)。結(jié)果顯示,在各個(gè)組織中均有表達(dá),眼部表達(dá)量最高,顯著高于其他6個(gè)組織(<0.05),肌肉、腎、心臟、腦表達(dá)量也相對(duì)較高,性腺和鰓表達(dá)量則相對(duì)低一些。在7個(gè)組織中也均有表達(dá),眼部表達(dá)量同樣顯著高于其他組織(<0.05),在鰓、腦、性腺、心臟組織中的表達(dá)量極低,只有微量的表達(dá),腎和肌肉中的表達(dá)量也相對(duì)較低(圖11)。

圖11 mitf2在7種組織中的表達(dá)差異分析
3 討論
3.1 mitf基因的cDNA全長(zhǎng)和氨基酸序列分析
通過RACE技術(shù)獲得橘色雙冠麗魚基因cDNA全長(zhǎng),2個(gè)亞型MITF1和MITF2均具有3個(gè)結(jié)構(gòu)域:靠近N末端的MITF_TFEB_C_3_N結(jié)構(gòu)域、bHLHZip結(jié)構(gòu)域和一個(gè)未知功能結(jié)構(gòu)域(DUF3371)。bHLHZip結(jié)構(gòu)域作為MITF蛋白的功能區(qū)域,在進(jìn)化過程中高度保守,其中,HLHZip區(qū)域能夠與自身或轉(zhuǎn)錄因子TFEB (transcription factor EB)、TFEC、TFE3形成同源或異源二聚體,通過Basic區(qū)域與特異的DNA結(jié)合行使蛋白功能,特異性DNA指包含5′-CACGTG-3′或5′-TCATGTG-3′結(jié)構(gòu)的DNA序列,即E-box或M-box (Bentley, 1994; Hemesath, 1994)。在哺乳動(dòng)物中,受不同啟動(dòng)子調(diào)控,編碼多個(gè)蛋白質(zhì)異聚體,包括MITF-A、B、C、D、E、H、J、M、MC、CM等(Hershey, 2005; Shiohara, 2008),在魚類中,從斑馬魚、青鳉分離得到的2種亞型和(Altschmied, 2002),這2種亞型在功能上發(fā)生了分化,對(duì)黑色素細(xì)胞的形成、增殖、遷移和分化起著重要的調(diào)控作用,而不參與黑色素細(xì)胞的發(fā)育,只在眼色素上皮細(xì)胞中表達(dá),參與眼調(diào)控的發(fā)育(Lister, 2014),本研究克隆的和為基因2個(gè)亞型,均屬于亞型,理化性質(zhì)分析結(jié)果顯示和的蛋白質(zhì)二級(jí)結(jié)構(gòu)極為相似,熒光定量結(jié)果顯示和除了在胚胎期表達(dá)量有差異外,在各個(gè)組織和各轉(zhuǎn)換時(shí)期表達(dá)量變化趨勢(shì)基本一致,因此,推測(cè)和更多保留了基因功能,未出現(xiàn)明顯基因功能分化。同源比對(duì)發(fā)現(xiàn),橘色雙冠麗魚、與尼加拉瓜湖始麗魚、薩伊藍(lán)六間、斑魚等魚類具有較高的同源性,與哺乳類、鳥類、爬行類等同源性較低,符合橘色雙冠麗魚傳統(tǒng)進(jìn)化地位。
3.2 mitf基因表達(dá)差異分析
3.2.1基因在胚胎發(fā)育時(shí)期的表達(dá)和在橘色雙冠麗魚8個(gè)胚胎發(fā)育時(shí)期均有表達(dá),其中,在受精卵時(shí)期表達(dá)量最高,顯著高于其他發(fā)育期(<0.05),表現(xiàn)出明顯的時(shí)期特異性。研究發(fā)現(xiàn),基因也在其他動(dòng)物胚胎發(fā)育期開始表達(dá),但表達(dá)的時(shí)期較晚,如非洲爪蟾胚胎在21/22期開始表達(dá)(Kumasaka, 2010),雞(Mochii, 1998)和鼠()(Nakayama, 1998)胚胎則在5日齡和9.5日齡開始表達(dá)。在青鳉中,基因在胚胎整個(gè)發(fā)育過程中最早表達(dá)于2細(xì)胞期(Li, 2013a、b),這與橘色雙冠麗魚表達(dá)模式相類似。蔣燕玲等(2016)對(duì)橘色雙冠麗魚胚胎組織學(xué)觀察發(fā)現(xiàn),黑色素細(xì)胞在血液循環(huán)期才開始出現(xiàn),而在受精卵時(shí)期即出現(xiàn)高表達(dá)現(xiàn)象,結(jié)合前人報(bào)道,推測(cè)魚類發(fā)育早期不僅調(diào)節(jié)黑色素細(xì)胞分化和發(fā)育,對(duì)視網(wǎng)膜色素上皮細(xì)胞、破骨細(xì)胞和肥大細(xì)胞也有不同程度的調(diào)控作用(Bauer, 2009)。在卵裂期,、基因表達(dá)量驟減,隨胚胎發(fā)育表達(dá)量繼續(xù)降低直至聽泡期驟升后又繼續(xù)下降,而隨胚胎發(fā)育表達(dá)量逐漸升高直至血液循環(huán)期,仍呈現(xiàn)上升趨勢(shì),預(yù)示基因可能在黑色素細(xì)胞的發(fā)育過程中扮演著十分重要的角色。
3.2.2基因在不同褪色時(shí)期表達(dá)模式 在3個(gè)體色過渡時(shí)期中,皮膚、鱗片、尾鰭的和的表達(dá)量均呈逐漸下降的趨勢(shì),其中,黑色時(shí)期表達(dá)量最高,除皮膚的基因表達(dá)量在3個(gè)過渡時(shí)期無顯著差異外,黑色時(shí)期鱗片和尾鰭中的表達(dá)量顯著高于灰色期和黃色期,與下游基因在橘色雙冠麗魚不同褪黑時(shí)期逐漸降低的表達(dá)模式基本一致(蔣燕玲等, 2016)。這與魚體表面各種色素細(xì)胞的比例與分布也存在對(duì)應(yīng)關(guān)系,韋敏俠等(2015)研究發(fā)現(xiàn),橘色雙冠麗魚在黃色時(shí)期皮膚中的黃色素細(xì)胞和紅色素細(xì)胞數(shù)量和分布比例增多,黑色素細(xì)胞相對(duì)減少且處于凝集狀態(tài)。在紅鯽魚發(fā)育過程中,基因也呈現(xiàn)相同的表達(dá)趨勢(shì),通過組織學(xué)觀察發(fā)現(xiàn),紅鯽魚黑色素細(xì)胞數(shù)量隨著其發(fā)育的進(jìn)行逐漸減少,體色由灰色逐漸過渡至紅色,表達(dá)量隨之降低(Zhang, 2017)。Liu等(2015)檢測(cè)了不同品系錦鯉中基因表達(dá)量,發(fā)現(xiàn)含有黑色素細(xì)胞較多的,體色偏白或偏黑的錦鯉表達(dá)量較高,而含黑色素細(xì)胞較少的,體色呈紅白或全黃的錦鯉皮膚的表達(dá)量偏低,表明表達(dá)量與黑色素細(xì)胞數(shù)量和分布比例呈正相關(guān),印證了基因?qū)谏丶?xì)胞發(fā)育和分布的重要作用,本研究中和均屬于,且也在黑色期表達(dá)量最高,與此結(jié)果相對(duì)應(yīng)。而且黃色時(shí)期橘色雙冠麗魚皮膚中可檢測(cè)到基因的表達(dá)(Liu, 2015),可見其可能還參與調(diào)控其他色素細(xì)胞形成。
3.2.3基因在成魚不同組織間的表達(dá)差異、亞型在橘色雙冠麗魚各組織中均有表達(dá),與在青鳉中的表達(dá)趨勢(shì)相類似(Li, 2013a、b),其中,眼部表達(dá)量最高且顯著高于其他組織,表明基因參與了橘色雙冠麗魚眼的發(fā)育調(diào)節(jié)。在哺乳動(dòng)物中,大量研究表明,基因的突變可能導(dǎo)致眼畸形的發(fā)生,如小鼠眼畸形(Hertwig, 1942)、犬眼睛出現(xiàn)缺陷(Stritzel, 2009)、牛眼睛變小(Wiedemar, 2014)等均與缺失有著密切關(guān)系,基因在哺乳動(dòng)物眼睛發(fā)育中有著不可或缺的作用。脈絡(luò)膜裂(CF)的閉合是眼睛發(fā)育的一個(gè)關(guān)鍵步驟,研究顯示,基因在顱神經(jīng)嵴細(xì)胞中發(fā)揮作用,促進(jìn)脈絡(luò)膜裂的閉合,是眼睛正常發(fā)育必不可少的調(diào)節(jié)基因(Michael, 2018)。但Lane等(2012)研究發(fā)現(xiàn),在具有基因突變的斑馬魚或、雙基因突變的斑馬魚中,其眼部發(fā)育正常,與野生型斑馬魚無明顯差異,這一結(jié)果說明了基因在斑馬魚眼睛發(fā)育過程中并非是必要的,這一結(jié)論具有顛覆性。因此,基因在動(dòng)物眼睛發(fā)育過程中是否為必需基因值得探討,但本研究中,和均表現(xiàn)為眼部表達(dá)量最高,仍然支持其在眼睛發(fā)育中起著必不可少的作用。
ALTSCHMIED J, DELFGAAUW J, WILDE B,. Subfunctionalization of duplicategenes associated with differential degeneration of alternative exons in fish. Genetics, 2002, 161(1): 259–267
BARLUENGA M, MEYER A. Phylogeography, colonization and population history of the Midas cichlid species complex (spp.) in the Nicaraguan crater lakes. BMC Evolutionary Biology, 2010, 10(1): 326–326
BAUER G L, PRAETORIUS C, BERGSTEINSDOTTIR K,. The role of MITF phosphorylation sites during coat color and eye development in mice analyzed by bacterial artificial chromosome transgene rescue. Genetics, 2009, 183(2): 581– 594
BENTLEY N J, EISEN T, GODING C R. Melanocyte-specific expression of the human tyrosinase promoter: Activation by the microphthalmia gene product and role of the initiator. Molecular and Cellular Biology, 1994, 14(12): 7996–8006
COHEN M A, WERT K J, GOLDMANN J,. Human neural crest cells contribute to coat pigmentation in interspecies chimeras after in utero injection into mouse embryos. Proceedings of the National Academy of Sciences of the United States of America, 2016, 113(6): 1570–1575
HAUSWIRTH R, HAASE B, BLATTER M,. Mutations in MITF and PAX3 cause “splashed white” and other white spotting phenotypes in horses. PLoS Genetics, 2019, 15(8): e1002653
HEMESATH T J, STEINGRIMSSON E, MCGILL G,. Microphthalmia, a critical factor in melanocyte development, defines a discrete transcription factor family. Genes and Development, 1994, 8(22): 2770–2780
HERSHEY C L, FISHER D E. Genomic analysis of the Microphthalmia locus and identification of the MITF-J/-J isoform. Gene, 2005, 347(1): 73–82
HERTWIG P. Neue Mutationen und Koppelungsgruppen bei der Hausmaus. Zeitschrift für Induktive Abstammungs- und Vererbungslehre, 1942, 80(1): 220–246
HOU L, PAVAN W J. Transcriptional and signaling regulation in neural crest stem cell-derived melanocyte development: Do all roads lead toCell Research, 2008, 18(12): 1163–1176
JIANG Y L, SONG H M, LIU Y,. Cloning and analysis of TYR gene and its development stages and tissue expression in. Chinese Journal of Agricultural Biotechnology, 2016, 24(5): 697–707 [蔣燕玲, 宋紅梅, 劉奕, 等. 橘色雙冠麗魚TYR基因的克隆及其發(fā)育時(shí)序和組織表達(dá)分析. 農(nóng)業(yè)生物技術(shù)學(xué)報(bào), 2016, 24(5): 697–707]
JIANG Y L. The development of body color ofand the cloning and analysis of body color-related gene TYR expression. Master′s Thesis of Shanghai Ocean University, 2016 [蔣燕玲. 橘色雙冠麗魚體色發(fā)育變化及體色相關(guān)基因TYR的克隆與表達(dá)研究. 上海海洋大學(xué)碩士研究生學(xué)位論文, 2016]
KAUTT A F, ELMER K R, MEYER A. Genomic signatures of divergent selection and speciation patterns in a natural experiment, the young parallel radiations of Nicaraguan crater lake cichlid fishes. Molecular Ecology, 2012, 21(19): 4770–4786
KHALED M. Glycogen synthase kinase 3beta is activated by cAMP and plays an active role in the regulation of melanogenesis. Journal of Biological Chemistry, 2002, 277(37): 33690–33697
KORBERG I B, SUNDSTROM E, MEADOWS J R S,. A simple repeat polymorphism in the MITF-M promoter is a key regulator of white spotting in dogs. PLoS One, 2014, 9(8): e104363
KUMASAKA M, SATO H, SATO S,. Isolation and developmental expression ofin. Developmental Dynamics, 2010, 230(1): 107–113
LANE B M, LISTER J A. Otx but nottranscription factors are required for zebrafish retinal pigment epithelium development. PLoS One, 2012, 7(11): e49357
LI M, ZHU F, HONG N,. Alternative transcription generates multipleisoforms with different expression patterns and activities in medaka. Pigment Cell and Melanoma Research, 2013a, 27(1): 48–58
LI M, ZHU F, HONG Y. Differential evolution of duplicated medaka fishgenes. International Journal of Biological Sciences, 2013b, 9(5): 496–508
LIN R, LIN W, ZHOU S,. Integrated analysis of mRNA expression, CpG island methylation, and polymorphisms in the MITF gene in ducks (). BioMed Research International, 2019(3): 8512467
LISTER J A, CAPPER A, ZENG Z Q,. A conditional zebrafish MITF mutation reveals MITF levels are critical for melanoma promotion vs. regression. Journal of Investigative Dermatology, 2014, 134(1): 133–140
LISTER J A, ROBERTSON C P, LEPAGE T,. Nacre encodes a zebrafish microphthalmia-related protein that regulates neural-crest-derived pigment cell fate. Development, 1999, 126(17): 3757–3767
LIU J H, WEN S, LUO C,. Involvement of thegene in the development of pigment cell in Japanese ornamental (Koi) carp (L.). Genetics and Molecular Research, 2015, 14(1): 2775–2784
MICHAEL H T, GRAFF-CHERRY C, CHIN S,. Partial rescue of ocular pigment cells and structure by inducible ectopic expression of-M in MITF-deficient mice. Investigative Ophthalmology and Visual Science, 2018, 59(15): 6067–6073
MINVIELLE F, BED'HOM B, COVILLE J L,. The “silver” Japanese quail and the MITF gene: Causal mutation, associated traits and homology with the “blue” chicken plumage. BMC Genetics, 2010, 11: 15
MOCHII M, MAZAKI Y, MIZUNO N,. Role ofin differentiation and transdifferentiation of chicken pigmented epithelial cell. Developmental Biology, 1998, 193(1): 47–62
NAKAYAMA A, NGUYEN M T T, CHEN C C,. Mutations in microphthalmia, the mouse homolog of the human deafness gene MITF, affect neuroepithelial and neural crest-derived melanocytes differently. Mechanisms of Development, 1998, 70(1/2): 155–166
NEWTON R A, ROBERTS D W, LEONARD J H,. Human melanocytes expressing MC1R variant alleles show impaired activation of multiple signaling pathways. Peptides, 2007, 28(12): 2387–2396
PARK H Y, KOSMADAKI M, YAAR M,. Cellular mechanisms regulating human melanogenesis. Cellular and Molecular Life Sciences, 2009, 66(9): 1493–1506
SHI X Y, XU Y J, WU N N,. Preliminary studies on blind-side hypermelanosis of: Chromatophores observation and expression of proopiomelanocortin. Progress in Fishery Sciences, 2015, 36(2): 45–54 [史學(xué)營, 徐永江, 武寧寧, 等. 半滑舌鰨()體表色素細(xì)胞觀察及POMC表達(dá)特性分析. 漁業(yè)科學(xué)進(jìn)展, 2015, 36(2): 45–54]
SHIOHARA M, SHIGEMURA T, SUZUKI T,. MITF-CM, a newly identified isoform of microphthalmia-associated transcription factor, is expressed in cultured mast cells. International Journal of Laboratory Hematology, 2008, 31(2): 215–226
STEINGRIMSSON E, COPELAND N G, JENKINS N A. Melanocytes and the Microphthalmia transcription factor network. Annual Review of Genetics, 2004, 38(1): 365–411
STRITZEL S, FRITSCHE J, WOEHLKE A,. Multiple ocular malformations in two sheepdogs homozygous for the merle mutation. Mutation analysis of the SILV gene and association study using MITF-associated markers. Tieraerztliche Praxis Ausgabe Kleintiere Heimtiere, 2009, 37(4): 229–238
VOLKENING A, SANDSTEDE B. Iridophores as a source of robustness in zebrafish stripes and variability inpatterns. Nature Communications, 2018, 9(1): 3231
WANG Y, VIENNET C, ROBIN S,. Precise role of dermal fibroblasts on melanocyte pigmentation. Journal of Dermatological Science, 2017, 88(2): 159–166
WEI M X, SONG H M, QI B L,. Pigment cells development and body color variation of postembryonic development in. Journal of Shanghai Ocean University, 2015, 24(1): 28–35 [韋敏俠, 宋紅梅, 祁寶倫, 等. 橘色雙冠麗魚胚后色素細(xì)胞發(fā)育與體色變化. 上海海洋大學(xué)學(xué)報(bào), 2015, 24(1): 28–35]
WIEDEMAR N, DROEGEMUELLER C. A 19-Mb de novo deletion on BTA 22 including MITF leads to microphthalmia and the absence of pigmentation in a Holstein calf. Animal Genetics, 2014, 45(6): 868–870
YAMADA T, AKAMATSU H, HASEGAWA S,. Melanocyte stem cells express receptors for canonical-signaling pathway on their surface. Biochemical and Biophysical Research Communications, 2010, 396(4): 837–842
YU D D, LIU H J, GUAN J,. Early ontogeny of chromatophores and body color changes of. Progress in Fishery Sciences, 2012, 33(5): 1–7 [于道德, 劉洪軍, 關(guān)健, 等. 黑棘鯛早期色素細(xì)胞發(fā)育與體色變化. 漁業(yè)科學(xué)進(jìn)展, 2012, 33(5): 1–7]
ZHANG Y, LIU J, PENG L,. Comparative transcriptome analysis of molecular mechanism underlying gray-to-red body color formation in red crucian carp (, red var.). Fish Physiology and Biochemistry, 2017, 43(5): 1387–1398
Structure and Expression Analysis of Body Color-RelatedGene in
CHEN Xudong1,2, WU Guoqiang1,2, SONG Hongmei2①, WANG Xuejie2, MU Xidong2, LIU Yi2, LIU Chao2, HU Yinchang2
(1.201306,; 2510380,)
Microphthalmia-associated transcription factor (MITF) is a transcription factor with a typical basic helix-loop-helix-leucine zipper structure. It is a key transcription factor in the melanin synthesis pathway in animals by activating downstream genes to initiate melanin synthesis. In order to explore the regulatory action ofduring body color variation in, the full-length cDNA of thegene was cloned using the rapid amplification of cDNA ends technique, and the difference in gene expression in different developmental stages of embryos, different stages of body melatonin, and various tissues inwere analyzed through qRT-PCR. Two mutant sequences ofwere obtained. The full-length ofcDNA was 1816 bp, including a 158 bp 5′UTR, a 428 bp 3′UTR, and a 1230 bp ORF coding a polypeptide of 409 amino acids. The full-length ofcDNA was 1638 bp, including a 160 bp 5′ UTR, a 428 bp 3′UTR, and a 1050 bp ORF coding a polypeptide of 349 amino acids. The analysis of the homology and phylogenetic tree results showed thatandclustered into a small branch in the phylogenetic tree, which had the highest homology with Cichlidae fish and low homology with mammals. The results of the qRT-PCR showed thatandwere expressed to varying degrees in various tissues of adult fish, among which the expression in the eyes was the highest and was significantly higher than in other tissues (<0.05), and the expression in muscle, brain, and kidney tissue was also higher. Theandgenes were expressed in all stages of embryonic development, and the expression level was highest in the cleavage stage, which was significantly higher than the other embryonic stages. With the body color transition from black to orange, the expression ofandgenes decreased gradually in the fish skin, scales, and tail fins. It showed that the expression ofwas positively correlated with changes in body color from black to yellow in, which may be related to the dynamic change in the differentiation and distribution ratio of pigment cells in the body color development stage of fish. Through understanding the molecular basis of fish body color development and variation, this study accumulates data for fish pigment cell development and the artificial improvement of fish body color.
;; qRT-PCR; Melanocyte
SONG Hongmei, E-mail: shm1227@126.com
S91
A
2095-9869(2022)02-0107-12
10.19663/j.issn2095-9869.20201215004
* 廣東省基礎(chǔ)與應(yīng)用基礎(chǔ)研究基金項(xiàng)目(2020A1515010304)、中國東盟海上合作基金(CAMC-2020F)和國家水產(chǎn)種質(zhì)資源共享服務(wù)平臺(tái)(NFGR-2020)共同資助 [This work was supported by Basic and Applied Basic Research Foundation of Guangdong Province (2020A1515010304), China-ASEAN Maritime Cooperation Fund (CAMC-2020F), and National Freshwater Genetic Resource Center (NFGR-2020)].陳旭東,E-mail: 979926926@qq.com
宋紅梅,副研究員,E-mail: shm1227@126.com
2020-12-15,
2021-01-26
陳旭東, 鄔國強(qiáng), 宋紅梅, 汪學(xué)杰, 牟希東, 劉奕, 劉超, 胡隱昌. 橘色雙冠麗魚體色相關(guān)基因的結(jié)構(gòu)及表達(dá)調(diào)控特性. 漁業(yè)科學(xué)進(jìn)展, 2022, 43(2): 107–118
CHEN X D, WU G Q, SONG H M, WANG X J, MU X D, LIU Y, LIU C, HU Y C. Structure and expression analysis of body color-relatedgene in. Progress in Fishery Sciences, 2022, 43(2): 107–118
(編輯 馮小花)
