慢性自發性蕁麻疹發病機制的生物信息學研究
2022-04-19 13:33:32賈秀娟烏日娜薩琦拉王百靈賀希格圖特格喜白音郭靖雪齊寶鵬木其日
中國麻風皮膚病雜志 2022年6期
賈秀娟 烏日娜 薩琦拉 王百靈 賀希格圖 特格喜白音 郭靖雪 齊寶鵬 木其日
1內蒙古國際蒙醫醫院皮膚科,呼和浩特,010020;2內蒙古國際蒙醫醫院創新蒙醫藥工程研究中心,呼和浩特,010070;3內蒙古醫科大學,呼和浩特,010110
蕁麻疹為一種以皮膚、黏膜小血管擴張并滲透性增加而出現的局限性水腫反應,臨床以風團及瘙癢為主要表現,較為常見且易復發,臨床分類主要包括自發性與誘導性兩類,自發性蕁麻疹根據病程是否大于6周分為急性自發性蕁麻疹與慢性自發性蕁麻疹,而誘導性蕁麻疹的分類依據則根據其發生是否與物理因素相關分為物理性與非物理性蕁麻疹,前者主要包括人工蕁麻疹(皮膚劃痕癥)、冷接觸性蕁麻疹、延遲壓力性蕁麻疹、熱接觸性蕁麻疹、日光性蕁麻疹、振動性血管性水腫、膽堿能性蕁麻疹,后者主要包括水源性蕁麻疹、接觸性蕁麻疹,其中病程持續至少6周的蕁麻疹為慢性蕁麻疹(chronic urticaria, CU),根據誘發因素的相關性,CU通常分為兩個亞型:慢性自發性蕁麻疹(chronic spontaneous urticaria, CSU)和慢性誘導性蕁麻疹(chronic inducible urticaria, CIndU)。CSU主要指無明確誘發因素反復出現皮膚、黏膜一過性(<24 h)的瘙癢性風團和/或血管性水腫,且病程持續至少6周;CIndU主要指由物理因素誘發的風團,反復出現病程持續6周以上,均嚴重影響患者的生活質量[1-3]。CSU的發病機制較復雜且尚不完全明確,故于治療方面也面臨巨大的的挑戰性,CSU的發生主要由肥大細胞驅動,由于其兼備自身免疫性與變態反應性疾病的特征,涉及到多因素,近些年各種炎癥遞質、自身免疫、肥大細胞與嗜堿粒細胞信號失調、凝血與氧化應激等被認為與其發病密切相關[4,5]。
CSU的發生是較為復雜的生物學過程,涉及多種機制且存在相互串擾、相互激活的關系[4]。近些年來,高通量測序技術聯合生物信息學分析在炎癥性皮膚病領域研究方面為探索疾病發病機制與治療靶點提供研究方向與思路的新手段[6,7]。因此,對參與CSU發病相關的關鍵基因及其相互作用、通路進行研究,可能為CSU的相關研究提供新方向與思路。本研究使用從GEO數據庫獲取的數據完備且可靠的GSE72541數據集,使用生物信息學方法,確定差異表達基因,并進一步對篩選出的差異表達基因進行GO分析、KEGG通路分析同時構建蛋白質相互作用(PPI)網絡,篩選顯著的PPI模塊,尋找關鍵通路與基因,進一步探究CSU發病的機制。
1 材料與方法
1.1 基因芯片數據 從Gene Expression Omnibus(GEO)數據庫獲取GSE72541數據集,該研究是基于GPL16699平臺(Agilent-039494 SurePrint G3 Human GE v2 8x60K Microarray 039381),由Giménez-Arnau A等提交,包括30個樣本,來自20個CSU及10個健康對照者的靜脈血標本。
1.2 差異基因篩選 對獲取的GSE72541芯片的矩陣數據進行背景校正、標準化、匯總與探針質量控制,同時獲取注釋包文件,將探針矩陣轉換為gene symbol矩陣。安裝加載limma包進行差異基因表達分析,篩選標準為(|logFC|>1 & P.Val<0.05),繪制差異表達基因的熱圖與火山圖。
1.3 功能富集分析 使用clusterProfiler包對差異基因(pvalue<0.05 & qvalue<0.05)進行功能富集分析,包括生物過程(BP)、分子功能(MF)、細胞組份(CC) 及 KEGG 通路的富集分析,得出顯著富集的功能與通路,此外分別對上調及下調基因進行功能及KEGG通路富集分析,P<0.05具有統計學意義。
1.4 蛋白質相互作用(PPI)網絡構建與分析 使用10.5版本STRING軟件對差異基因相應蛋白的相互作用關系進行構建與分析。PPI網絡圖中綜合得分>0.4的蛋白具有統計學意義,并使用Cytoscape插件MCODE篩選出有意義的PPI模塊,篩選標準為: Node score cutoff>0.2和K-core>2。
2 結果
2.1 篩選差異基因 以|logFC|>1 & adj.P.Val<0.05為標準共篩選出86個差異表達基因,其中上調基因64個,如ITGB3、AQP10等;下調基因22個,如HLA-DQA1、KRTAP19-2。采用R語言繪制火山圖(圖1a)與上調基因與下調基因的熱圖(圖1b)。其中,上調及下調差異表達基因Top10與差異倍數見表1。
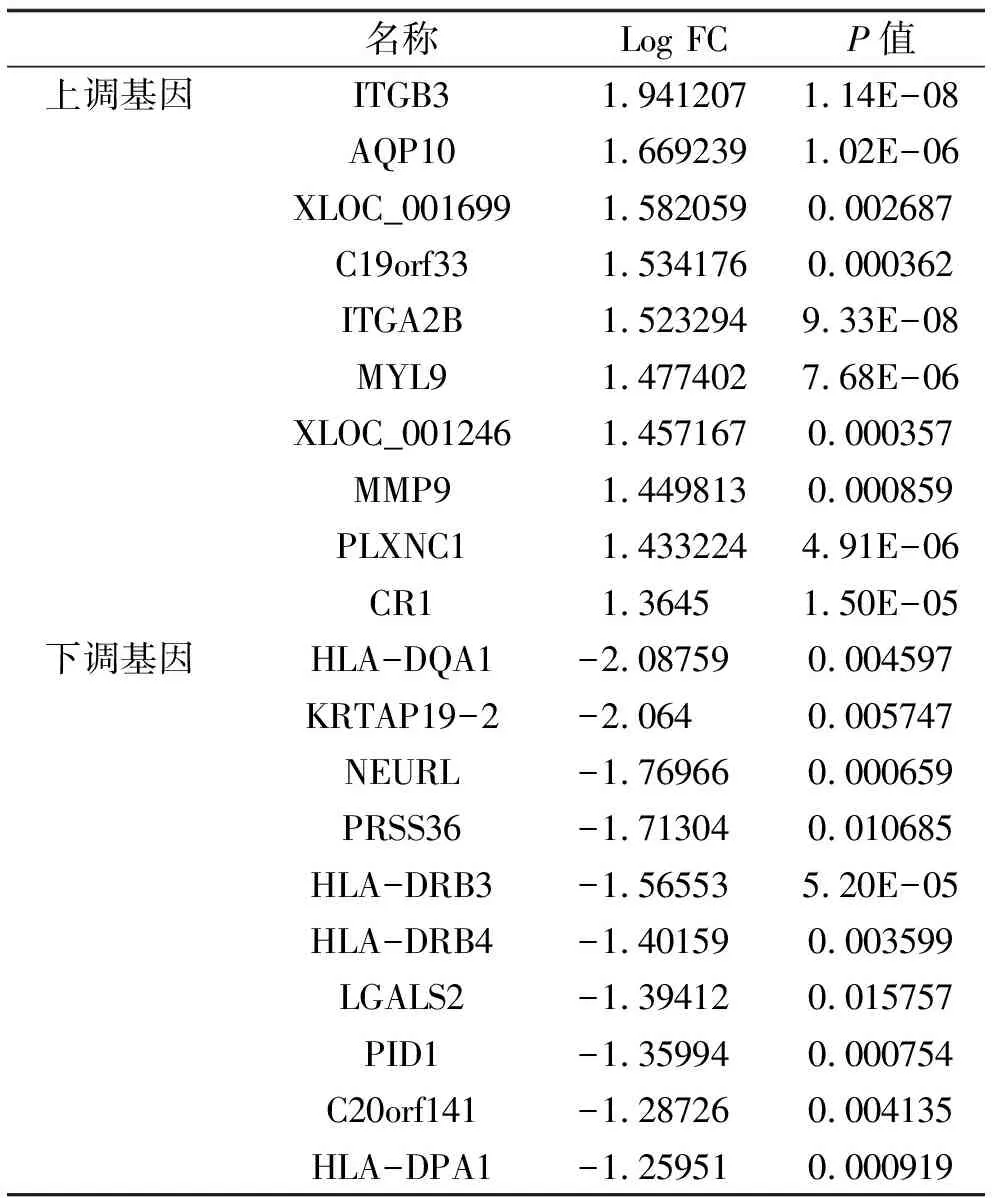
表1 Top10上調及下調的基因
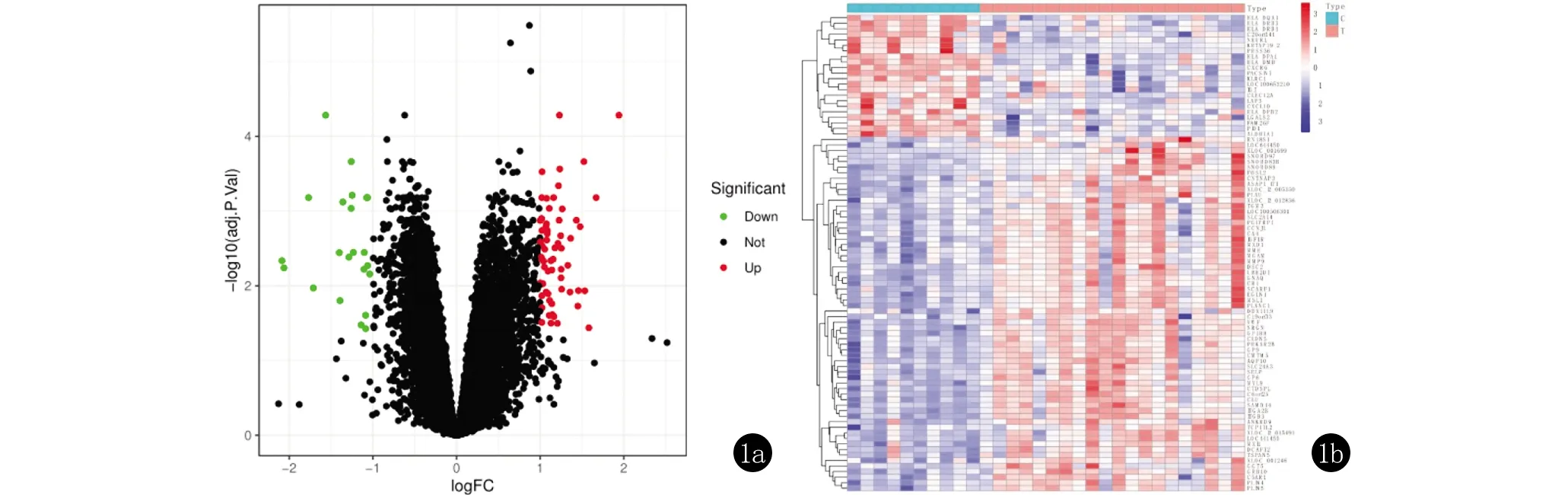
圖1 差異表達基因的火山圖(1a)與熱圖(1b)
2.2 GO及KEGG通路分析 GO富集分析(圖2)表明,CSU組與健康對照組相比的差異表達基因主要涉及血小板活化、血液凝固、止血、凝固、β淀粉樣蛋白清除(amyloid-beta clearance)、MHC II類分子介導的外源性肽抗原的抗原加工和提呈(antigen processing and presentation of exogenous peptide antigen via MHC class II)、MHC II類分子介導的肽抗原的抗原加工和提呈(antigen processing and presentation of peptide antigen via MHC class II)、MHC II類分子介導的肽或多糖抗原的抗原加工和提呈(antigen processing and presentation of peptide or polysaccharide antigen via MHC class II)、內源性凝血途徑、蛋白質活化級聯等生物學過程;富集在MHC II類蛋白復合物(MHC class II protein complex)、MHC蛋白復合物(MHC protein complex)、分泌顆粒膜、內質網膜腔側的組成部分(integral component of lumenal side of endoplasmic reticulum membrane)、內質網膜腔側(lumenal side of endoplasmic reticulum membrane)、血小板α顆粒、網格蛋白包被的內吞泡膜(clathrin-coated endocytic vesicle membrane)、反向高爾基網(trans-Golgi network)、三級顆粒、血小板α顆粒膜等細胞組份;參與MHC II類受體活性(MHC class II receptor activity)、肽抗原結合、糖結合、肽結合、胰島素樣生長因子I結合(insulin-like growth factor I binding)、酰胺結合、膠原結合、轉移酶活性,轉移氨基酰基、補體結合、胰島素受體結合、MHC蛋白復合物結合(MHC protein complex binding)等分子功能。
KEGG分析顯示,富集的主要通路包括:造血細胞系、移植物抗宿主病、金黃色葡萄球菌感染、哮喘、同種異體移植排斥反應、利什曼病(Leishmaniasis)、抗原加工和提呈、I型糖尿病、血小板活化、產生IgA的腸道免疫網絡、ECM受體相互作用(ECM-receptor interaction)、自身免疫性甲狀腺疾病、細胞間黏附分子、病毒性心肌炎、炎癥性腸病、補體途徑、Th1和Th2細胞分化、類風濕性關節炎、吞噬體、甲型流感、Th17細胞分化、弓形蟲病、結核病、中性粒細胞胞外陷阱形成、EB 病毒感染、系統性紅斑狼瘡、黏著斑、人T細胞白血病病毒1感染、致心律失常性右室心肌病、前列腺癌(prostate cancer)等信號通路過程。
此外,對篩選出的差異表達基因按照上調及下調分別進行了GO與KEGG富集分析,結果見圖3。功能富集分析結果顯示,上調基因主要富集于血小板活化、血液凝固、止血、凝固等生物學過程,下調基因則主要富集于MHC II類分子介導的外源性肽抗原的抗原加工和提呈、MHC II類分子介導的肽抗原的抗原加工和提呈、MHC II類分子介導的肽或多糖抗原的抗原加工和提呈等生物學過程。KEGG通路富集分析,上調基因富集的主要通路為血小板活化、ECM受體相互作用、造血細胞系、中性粒細胞胞外陷阱形成、而下調基因主要富集在移植物抗宿主病、抗原加工和提呈、甲型流感、EB 病毒感染等信號通路過程。
PPI網絡為一種突出蛋白-蛋白間相互作用的重要方式,可進一步反應機體的各項功能[8]。將篩選出的差異表達基因上傳至STRING數據庫中,構建、繪制PPI網絡圖,該網絡由62個節點蛋白和257條邊構成(圖4a)。使用Cytoscape的MCODE插件進行子網絡分析,得到PPI網絡的子網絡模塊,篩選出3種最顯著的模塊(圖4b~4d、表2)。其中,最顯著的相互作用模塊見4B,包括CXCL10, MMP9, SELP, MME, ITGA2B, ITGB3, CLU, GP6, C5AR1, C6orf25, GP1BB, VWF, CR1, GP9, PLAU共15個關鍵基因,存在密切的相互作用關系。
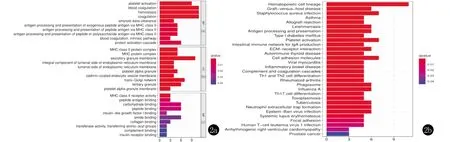
2a :GO功能富集柱狀圖(a)生物學過程;(b)細胞組份;(c)分子功能圖;2b:KEGG通路分析柱狀圖
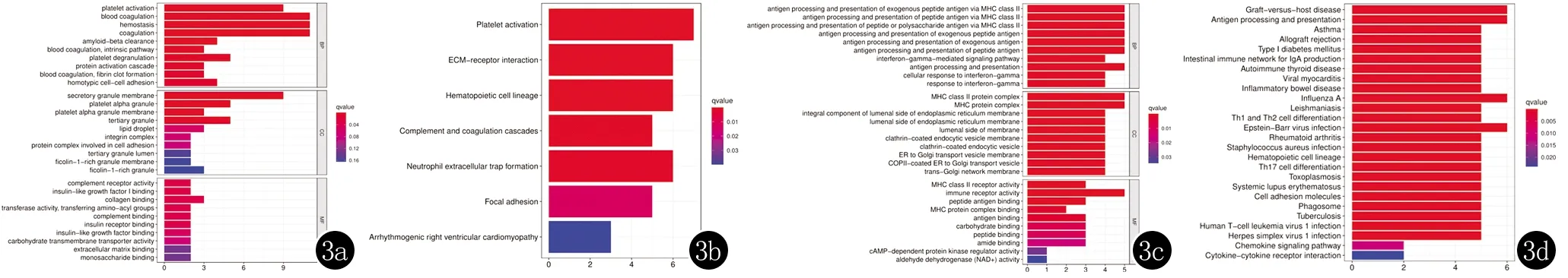
3a :上調基因的GO功能富集柱狀圖;3b:上調基因的KEGG通路分析柱狀圖;3c :下調基因的GO功能富集柱狀圖;3d:下調基因的KEGG通路分析柱狀圖
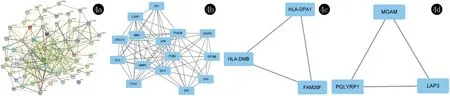
4a:蛋白-蛋白相互作用網絡圖;4b~4d:顯著模塊分析圖

表2 3種最顯著蛋白-蛋白相互作用模塊
3 討論
本研究通過分析CSU的基因芯片結果,發現CSU與健康對照者相比共有86個差異基因表達,其中上調基因64個,下調基因22個。將篩選出的差異基因進行GO富集及KEGG通路分析,這些差異表達基因主要富集在凝血、免疫調節、感染等過程。凝血級聯反應參與了CSU的發病機制,其主要為組織因子與補體(如C3a、C5a)所觸發,目前對組織因子及補體所觸發的凝血級聯反應參與CSU皮膚肥大細胞與外周嗜堿粒細胞的機制不清楚[3,9,10]。肥大細胞與嗜堿粒細胞激活誘導的促炎癥因子釋放為CSU的重要的病理生理學特征,諸多證據表明高達50%的CSU患者存在潛在的自身免疫相關的病因,如Th1/Th2細胞因子失衡、補體活化、肥大細胞與嗜堿粒細胞異常分布等,CSU的發生常合并自身免疫性疾病,如甲狀腺疾病、乳糜瀉、類風濕關節炎、系統性紅斑狼瘡等[11-13]。盡管多數情況下CSU發生的確切原因無法明確,但其發生與感染具有一定的關聯性,關于感染誘發CSU的機制尚不完全清楚,但目前認為感染誘發的免疫與非免疫機制均介導了CSU的發生,主要包括IgE依賴和非IgE依賴的免疫機制,而非免疫機制亦涉及多方面,如通過Toll樣受體介導肥大細胞活化、通過誘發系統性炎癥反應活化肥大細胞、感染后刺激機體產生抗體,結合后形成抗原抗體復合物后活化補體,進而由補體介導肥大細胞脫顆粒,或直接刺激肥大細胞脫顆粒等[14,15]。總之,CSU的發生是涉及多種因素的較為復雜的生物學過程。
通過構建蛋白-蛋白間相互作用網絡圖篩選顯著模塊,共篩選出了15個關鍵基因,包括CXCL10, MMP9, SELP, MME, ITGA2B, ITGB3, CLU, GP6, C5AR1, C6orf25, GP1BB, VWF, CR1, GP9, PLAU。其中,ITGB3,ITGA2B,MMP9及CR1是差異表達最為顯著的top10上調基因。ITGB3 (β3 integrin, β3整合素),又稱CD61或GP3A;ITGA2B(integrin Alpha 2b, Alpha 2b整合素),又稱CD41,兩者均為整合素家族成員,整合素作為與細胞外基質配體、細胞表面配體和可溶性配體結合的細胞黏附受體家族,是細胞表面的黏附受體及細胞表面的重要分子,參與多種生物學過程,如發育、免疫反應、止血和傷口愈合等,其還作為信號轉導受體發揮作用,參與調控調節細胞存活、增殖和細胞命運的胞內途徑等功能[16-18]。細胞間的相互黏附作為人類機體免疫功能正常運作的基礎,黏附分子的異常表達可導致免疫功能異常,其中整合素在細胞之間的相互作用中發揮著重要作用[19]。目前對于ITGB3, ITGA2B參與蕁麻疹病理生理學的確切機制與意義不完全清楚。此外,MMP9在本研究中上調表達也較為顯著,基質金屬蛋白酶(MMPs)為鋅依賴性蛋白水解酶, 其中MMP9是一種最復雜的基質金屬蛋白酶,MMP9具有降解細胞外基質成分的能力,在病理生理功能中起重要作用,其過度表達和失調與多種疾病相關[20]。MMP9作為一種促進炎癥和組織重塑的酶,可能也參與了蕁麻疹的病理生理過程,但多項研究表明了MMP9水平與疾病嚴重程度的相關性的矛盾結果,部分研究認為MMP9水平升高與疾病嚴重程度呈正相關,升高的MMP9水平并被視為CSU的生物標志物,然而近期研究表明MMP9水平與CSU的嚴重程度無關,MMP9水平并未隨著治療過程而降低,結論認為MMP9不能作為CSU疾病活動的生物標志物,也不能用于評估治療效果[21-24]。因此,盡管于本研究MMP9呈現顯著上調表達,但基于現有的研究分析MMP9參與CSU的病理生理學機制、作為CSU生物標志物的臨床意義及其潛在價值仍有爭議,亦值得進一步研究。CR1(complement receptor 1, 補體受體1)為C3b/C4b補體的受體,存在于不同的細胞、血漿與尿液中,CR1作為一種多功能多態性糖蛋白,在紅細胞、嗜酸性粒細胞、單核細胞、巨噬細胞、B淋巴細胞、CD4+T細胞亞群、樹突狀細胞、皮膚中的朗格罕細胞等表達[25-29]。補體過度激活是一系列自身免疫性和炎癥性疾病發病機制中的關鍵事件,而CR1在免疫復合物沉積和補體系統過度激活導致組織損傷的疾病過程中是一個極其重要的分子[26]。補體在蕁麻疹發病中起主要或加重因素的作用,補體(C3a和C5a)觸發的外源性凝血級聯參與了CSU的發病機制[9,30]。外周血紅細胞CR1的表達水平在慢性蕁麻疹患者中顯著高于健康對照組,且血清總免疫球蛋白E水平與外周血紅細胞CR1表達水平也呈顯著正相關,CR1參與了CSU發病機制[31]。
綜上所述,本研究使用生物信息學方法全面分析了CSU與健康對照組間的差異表達基因,并篩選出CXCL10, MMP9, SELP, MME, ITGA2B, ITGB3, CLU, GP6, C5AR1, C6orf25, GP1BB, VWF, CR1, GP9, PLAU 15個關鍵差異表達基因在CSU的發生和發展中發揮重要作用,它們參與的生物過程、細胞組份、分子功能及相關信號通路可能與CSU的發病密切相關。生物信息學分析對進一步深入了解、研究CSU的生物學機制與治療靶點提供了新思路與方向。
猜你喜歡
四川勞動保障(2021年9期)2022-01-18 05:11:08
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
文苑(2018年21期)2018-11-09 01:23:06
山東工業技術(2016年15期)2016-12-01 05:31:22
中國衛生(2016年9期)2016-11-12 13:28:08
中國衛生(2015年9期)2015-11-10 03:11:12
中國衛生(2014年3期)2014-11-12 13:18:12
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
中國火炬(2014年4期)2014-07-24 14:22:19
