赤擬谷盜氣味結(jié)合蛋白研究進展
2022-04-26 08:16:52桑育黎辛躍強韓甜甜范廣文
遼寧大學學報(自然科學版) 2022年1期
桑育黎,石 磊,辛躍強,韓甜甜,范廣文
(遼寧大學 藥學院,遼寧 沈陽 110036)
赤擬谷盜(TriboliumcastunemHerbst)為鞘翅目,擬步甲科,擬谷盜屬昆蟲,是一種重要的世界性倉儲害蟲[1],廣泛分布于熱帶與較溫暖地區(qū),在我國至少23個省(區(qū))有分布[2].研究表明,赤擬谷盜對溫度要求較高,35℃時卵孵化率最高[3-4].其食性復雜,主要寄生于小麥、玉米、稻谷、高粱、干果、中草藥、生藥材等.其成蟲體表具有臭腺,該腺體可分泌苯醌等致癌物質(zhì)[5],會使面粉等商品在儲藏過程中出現(xiàn)結(jié)塊、變色及發(fā)臭等現(xiàn)象而不能使用[6].由于其壽命較長,繁殖迅速,食性復雜,適應性強,可對多種商品造成嚴重損害,導致巨大的經(jīng)濟損失.目前,赤擬谷盜的基因組測序已經(jīng)完成,已成為第二大模式昆蟲[7].
昆蟲借助嗅覺系統(tǒng)感知外界環(huán)境信息,指導其進行覓食、交配、繁殖及躲避天敵等活動[7].據(jù)報道,其外周嗅覺系統(tǒng)包含多種蛋白,如氣味受體(Odorant receptors,Ors)[8]、氣味結(jié)合蛋白(Odorant-binding proteins,OBPs)[9-11]、化學感受蛋白(Chemosensory proteins,CSPs)[12]、離子受體(Ionotropic receptors,IRs)[13]、感覺神經(jīng)元膜蛋白(Sensory neuron membrane proteins,SNMPs)[14]和氣味降解酶(Odor degrading enzymes,ODEs)[15-16]等功能蛋白.氣味分子等化學信號通過感受器小孔進入感受器管腔,并通過淋巴液擴散,淋巴液中含有多種蛋白,其中包括氣味結(jié)合蛋白等,氣味結(jié)合蛋白與氣味分子結(jié)合到達嗅覺感受器,激活受體,以上過程是生物感受外界氣味信息的第一步.文獻報道昆蟲可以通過嗅覺特異性感知環(huán)境中的氣味分子,從而指導其行為.例如Gao等[17]在研究赤擬谷盜幼蟲接觸北艾揮發(fā)油的機制時,發(fā)現(xiàn)赤擬谷盜幼蟲的OBP/CSP等蛋白表達量顯著上調(diào),當使用RNAi抑制TcOBP11與TcCYP4BN6表達時,赤擬谷盜幼蟲在接觸北艾揮發(fā)油后,致死量顯著增加,提示TcOBP11可以通過驅(qū)使赤擬谷盜遠離北艾揮發(fā)油,參與到赤擬谷盜防御機制中.
昆蟲氣味結(jié)合蛋白的特性對赤擬谷盜防治有著特殊意義,而在生物防治過程中,采用環(huán)保、經(jīng)濟的天然植物揮發(fā)油制成的驅(qū)避劑不僅具有環(huán)保及商業(yè)價值,也成為研究的熱點,赤擬谷盜作為第二大模式昆蟲,研究其氣味結(jié)合蛋白亦對昆蟲氣味結(jié)合蛋白的總體研究進展有著重要的影響.
1 氣味結(jié)合蛋白
目前,學界對昆蟲OBPs研究較為豐富,研究發(fā)現(xiàn)OBPs多儲存在昆蟲的嗅覺受體神經(jīng)元周圍的細胞外液之中和感受器的淋巴液之中.OBPs是一類水溶性小分子蛋白,一般偏酸性,典型的OBPs通常由120~150個氨基酸組成[18],它屬于載體蛋白超家族——脂質(zhì)蛋白.昆蟲的OBPs與脊椎動物的OBPs具有不同的折疊模式,脊椎動物OBPs通常由8個β-筒狀結(jié)構(gòu)和羧基末端1個α-螺旋組成,而昆蟲OBPs則是由6個α-螺旋所組成,且排列緊湊,包圍出1個疏水腔[19-22].昆蟲的OBPs結(jié)構(gòu)中還通過3個二硫鍵進行穩(wěn)定結(jié)構(gòu),使其具有更好的熱穩(wěn)定性與保守性.
OBPs傳遞氣味分子的功能最早見于鱗翅目類昆蟲的觸角之中,對多種昆蟲的體內(nèi)克隆,均得到了OBPs基因,其不同種類間存在極大差異.
2 氣味結(jié)合蛋白的種類
第一個鑒定出的昆蟲OBPs為1981年Vogt等[23]從多音大蠶蛾中得到的性外激素結(jié)合蛋白(pheromone-binding proteins,PBPs),隨后越來越多的OBPs被鑒定出來.昆蟲嗅覺感受器中含有大量OBPs.Engsontia等[8]研究發(fā)現(xiàn),赤擬谷盜相較于其他昆蟲具有更多的氣味受體基因,在341個氣味受體基因中,含有259個氣味結(jié)合蛋白基因.目前許多研究將OBPs分為3個亞家族,包含PBPs[23]與普通氣味結(jié)合蛋白(general odorant-binding proteins,GOBPs),PBPs主要分布在雄性昆蟲體內(nèi),而GOBPs則與之有根本不同,在雄性雌性體內(nèi)皆有分布與表達,其基因序列具有高度保守性,在不同物種之中可能具有相似作用,一般認為與PBPs不同的是,GOBPs除了能結(jié)合性激素外,也發(fā)揮著與一般氣味分子結(jié)合的功能,因此被稱為GOBPs,且根據(jù)其保守氨基酸序列不同又可分為GOBPs1與GOBPs2[10].不同亞家族的OBPs具有不同的特性,且與不同氣味特異性識別感覺神經(jīng)元相關.還有一些研究依據(jù)保守的半胱氨酸數(shù)量和特征將OBPs分為經(jīng)典OBPs(含有6個半胱氨酸殘基)、Plus-C OBPs(較經(jīng)典OBPs多出1個脯氨酸和2個半胱氨酸殘基)、Atypic OBPs(非經(jīng)典氣味結(jié)合蛋白具有9~10個半胱氨酸殘基位點)和Minus-C OBPs(較經(jīng)典OBPs缺少2個半胱氨酸殘基).
3 氣味結(jié)合蛋白的表達分布
對于赤擬谷盜OBPs表達部位,Dippel等[24]研究發(fā)現(xiàn),赤擬谷盜50個OBPs中,共49個分別在赤擬谷盜觸角、頭、口器、腿及軀干中有表達,并通過MALDI-TOF MS蛋白質(zhì)指紋圖譜分析發(fā)現(xiàn),有3個經(jīng)典OBPs和大部分C-OBPs在其他部位有表達,這說明昆蟲OBPs表達不僅僅局限在觸角中.現(xiàn)代研究也發(fā)現(xiàn),昆蟲OBPs在其他部位也有高表達,與上述論述相符,如南方家蠅庫蚊[25]、岡比亞按蚊[26]、蜜蜂[27]、煙草煙粉虱[28]等昆蟲的某些OBPs,在昆蟲的其他部位也有高表達現(xiàn)象.
研究亦發(fā)現(xiàn),赤擬谷盜大多數(shù)經(jīng)典OBPs集中分布在觸角、腿和口器等部位的化學感受器中,20個經(jīng)典OBPs中的15個在觸角及口器中顯著表達[24].目前研究發(fā)現(xiàn),OBPs大量分布于感受器的淋巴液中.電鏡掃描結(jié)果顯示,赤擬谷盜成蟲不同性別,其觸角形態(tài)類似,共有5種感受器,分別為Bohm氏鬃毛、毛形感受器、刺形感受器、錐形感受器和栓錐形感受器,上述感受器僅在長度上不相同,而其中毛形感受器1型(ST1)、分叉錐形感受器(SB1)、錐形感受器2型(SB2)、錐形感受器3型(SB3)、錐形感受器4型(SB4)被推測具有嗅覺感受功能或化學感受功能[29].
大量研究發(fā)現(xiàn),不同齡、性別昆蟲OBPs表達量亦不同.如張穎[30]通過蛋白印跡分析發(fā)現(xiàn)SzeaOBP1和SzeaOBP28雖在玉米象幼蟲及成蟲階段都有表達,然而表達量并不相同,SzeaOBP10和SzeaOBP40在低齡幼蟲中表達更為豐富;松褐天牛MaltOBP9基因在雌蟲中表達顯著高于雄蟲[31].針對赤擬谷盜各階段及不同性別之間表達量的差異,目前尚不明確.
4 氣味結(jié)合蛋白的生理功能
在參與昆蟲行為時,OBPs能對識別外界氣味分子起重要作用.一般認為,OBPs的作用是對某些氣味分子特異性識別并結(jié)合,助溶并運輸氣味分子,以及使已完成刺激的氣味分子失活等.OBPs在參與昆蟲生理及行動過程中主要存在3種假說機制:1)OBPs與氣味分子結(jié)合,形成復合物,使脂溶性的氣味分子水溶性增大,最終以復合物的形式同受體分子結(jié)合[32];2)OBPs先與氣味分子結(jié)合形成復合物,通過淋巴液后解離[33];3)OBPs與氣味分子結(jié)合形成復合物,再通過淋巴液并不及時解離,而是先與受體膜的跨膜蛋白結(jié)合,在該作用下,促進氣味分子和OBPs解離[34].無論哪一種機制都認為,OBPs與氣味分子的特異性結(jié)合能增強氣味分子的水溶性,并通過感受器中水溶性的淋巴液到達神經(jīng)樹突上的受體,使氣味受體接受刺激,最終氣味降解酶將氣味分子降解,避免持續(xù)刺激達到保護作用.圖1所示昆蟲感受器內(nèi)氣味結(jié)合蛋白識別運輸氣味分子過程.
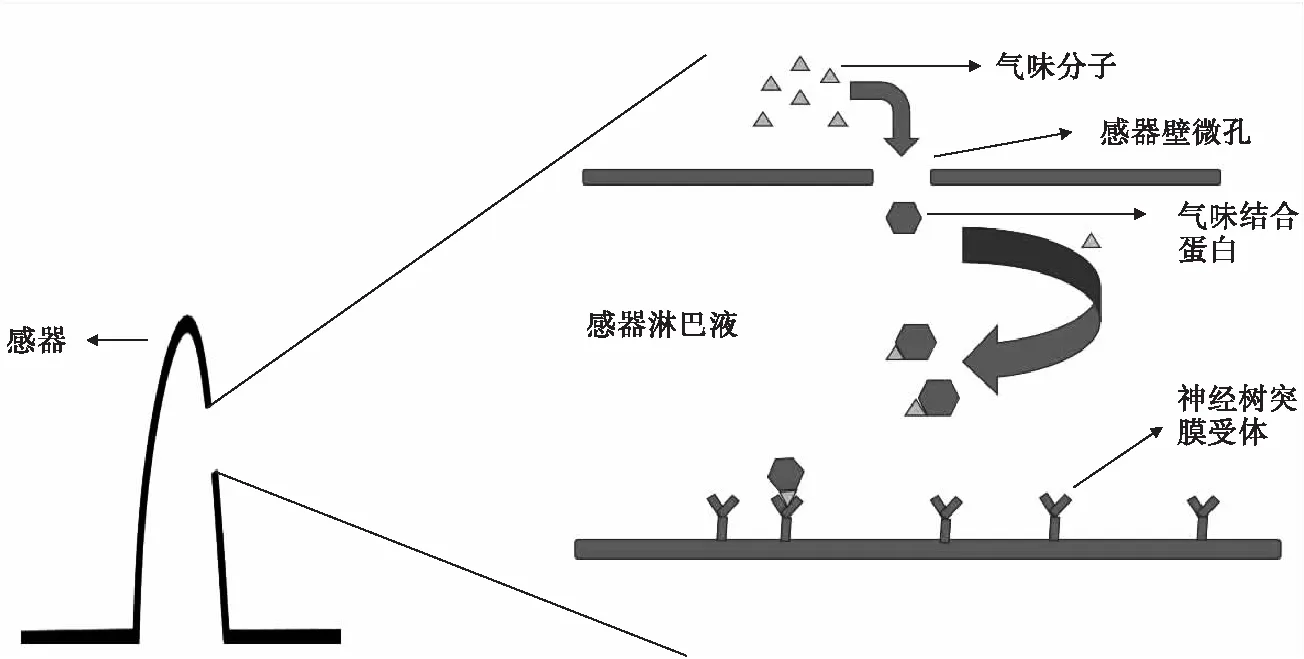
圖1 昆蟲感器內(nèi)氣味結(jié)合蛋白識別運輸氣味分子過程
OBPs可能使赤擬谷盜避開某種有害物質(zhì)氣味源,或以使其趨向某種氣味源等方式指導并參與赤擬谷盜行為調(diào)節(jié).Zhang等[35]研究報道,中藥艾蒿揮發(fā)油對赤擬谷盜具有較強的觸殺作用,且顯著誘導赤擬谷盜OBPC11基因.研究亦發(fā)現(xiàn),針對赤擬谷盜OBPC11進行RNA干擾后,赤擬谷盜在接觸艾蒿揮發(fā)油后具有更高的死亡率,提示赤擬谷盜OBPC11基因與赤擬谷盜的防御機制有關.Conteras等[36]研究發(fā)現(xiàn),赤擬谷盜幼蟲在防御特異性蘇云金芽孢桿菌Cry毒性時,OBPC12等蛋白表達量顯著性上調(diào),通過使用RNA抑制敲除OBP基因后,研究驗證了赤擬谷盜OBP參與到防御過程中,提示OBP參與赤擬谷盜幼蟲的基礎防御機制.李豫強等[37]研究小麥氣味中誘集赤擬谷盜,發(fā)現(xiàn)2-庚酸、己酸、辛酸等與小麥細麩皮混合后,可提高小麥對赤擬谷盜的誘集效果,提示赤擬谷盜可感知3種成分,并引誘赤擬谷盜.研究發(fā)現(xiàn)[38],赤擬谷盜感染縮小膜殼絳蟲后,其性外激素結(jié)合蛋白顯著上調(diào),提示其可能參與赤擬谷盜防御機制,但具體機制尚不清楚.
5 氣味結(jié)合蛋白的研究方法
5.1 獲取方法
在對OBPs研究時,體外研究是常采用的手段,通過對離體蛋白進行測定,可以對其結(jié)合特性進行一系列的研究,以解釋其表達特性、分布規(guī)律、結(jié)合靶點等.經(jīng)典的OBPs研究方法往往采用從昆蟲體內(nèi)直接提取的方法.然而,由于昆蟲體內(nèi)OBPs含量并不足以支撐實驗所需,如果想要得到足以達到實驗所需的蛋白量,則需要大量的昆蟲樣本,因此目前研究人員常采用基因克隆,原核表達的方式,即先構(gòu)建所需基因序列,克隆后連接表達載體即質(zhì)粒,之后轉(zhuǎn)入原核細胞中誘導表達,最終在體外完成表達得到重組蛋白,再通過親和層析柱對蛋白進行純化,得到純化蛋白的方法.孫小潔等[39]通過該方法得到綠盲蝽的氣味結(jié)合蛋白AlucOBP33蛋白;張穎[30]亦通過SzeaOBPs的序列分析及原核表達最終得到SzeaOBP1與SzeaOBP40;李秋玲[40]通過構(gòu)建班氏跳小蜂觸角轉(zhuǎn)錄組,并以表達量為挑選標準得到一部分基因序列,通過以上方法,最終得到大量的AbamOBPs;余海忠[41]通過該方法得到二化螟GOBP2.在測定昆蟲OBPs表達特性時,常采用實時熒光定量PCR等手段來分析其表達規(guī)律及分布特點,研究OBPs的表達規(guī)律與分布特點亦有助于理解OBPs的生理功能.
5.2 檢測方法
在探究昆蟲OBPs功能的過程中,測定OBPs與氣味分子的結(jié)合能力是一項重要的工作,并且,由于OBPs的功能特性,通過測定某種特定昆蟲OBPs與特定的氣味分子結(jié)合能力,往往可以推斷出該氣味分子是否對昆蟲的行為產(chǎn)生影響,從而可以篩選出能夠?qū)崿F(xiàn)目標價值的驅(qū)避劑、殺蟲劑,甚至擴展昆蟲OBPs應用場景,實現(xiàn)對某種特定揮發(fā)性分子的識別與捕捉.目前,對于昆蟲OBPs的研究方法主要包括分子對接技術(shù)、熒光競爭性結(jié)合實驗、熒光猝滅實驗和同源建模等,這些技術(shù)在研究昆蟲OBPs與目標分子的結(jié)合機制研究中被廣泛應用.分子對接技術(shù)往往先通過對蛋白質(zhì)3D建模,而后通過DOCK、Auto DOCK或FlexX等分子對接軟件進行剛性對接、半柔性對接或柔性對接匹配,如Zhang等[42]就通過Auto DOCK軟件實現(xiàn)對柑橘木虱OBPs和CSPs的分子對接實驗,完成對柑橘木虱OBPs結(jié)合的分子機制研究.然而分子對接技術(shù)只是計算機系統(tǒng)上模擬OBPs與氣味分子的結(jié)合,結(jié)果仍需其他實驗進行佐證,在單獨作為例證時存在一定的爭議.熒光競爭性結(jié)合實驗常用來檢驗所得到的OBPs和CSPs與目標配體結(jié)合的情況,其基本原理:當氣味分子進入OBPs疏水腔時,與腔內(nèi)氨基酸殘基結(jié)合,而熒光分子探針與疏水腔內(nèi)氨基酸殘基亦發(fā)生分子間作用力,并在特定波長下,激發(fā)出熒光,氣味分子和熒光分子探針與OBPs發(fā)生競爭性結(jié)合,因此,氣味分子與OBPs結(jié)合能力越強,熒光強度越弱.Li等[43]得到梨小食心蟲的GmolOBP12和GmolOBP16,通過熒光競爭性結(jié)合實驗發(fā)現(xiàn),二者均與宿主植物揮發(fā)性物質(zhì)梨酸酯親和力強,且能特異性識別其他某些化學物質(zhì).相較于分子對接技術(shù),熒光競爭性結(jié)合實驗針對離體蛋白進行實驗具有更好的說服力.然而,無論是通過原核生物(或真核生物)構(gòu)建的OBPs還是純化分離所得到OBPs,其體內(nèi)外生物活性是否具有一定差異,而這種差異是否會引發(fā)實驗數(shù)據(jù)結(jié)果誤差,仍在學界廣泛討論中.
6 展望
目前,許多研究發(fā)現(xiàn),天然植物提取物對昆蟲具有驅(qū)避、觸殺等作用.Zhang等[44]報道,北艾揮發(fā)油對赤擬谷盜具有觸殺作用,可用于對赤擬谷盜的防治;Chen等[45]報道北蒼術(shù)揮發(fā)油對赤擬谷盜以及嗜卷書虱具有觸殺和驅(qū)避作用;Abouelatta等[46]報道天竺葵揮發(fā)油對赤擬谷盜及谷蠹具有顯著的驅(qū)避、觸殺及熏蒸活性;韓群鑫等[47]報道丁香花蕾粉末與丁香酚對黑菌蟲具有驅(qū)避作用,對煙草甲具有引誘作用,其中丁香酚為主要化學作用物質(zhì),研究同樣發(fā)現(xiàn)丁香對赤擬谷盜具有驅(qū)避作用;Kheloul等[48]發(fā)現(xiàn)薰衣草揮發(fā)油對赤擬谷盜具有驅(qū)避效應.因此在天然植物揮發(fā)油中尋找合適的物質(zhì),對赤擬谷盜等害蟲進行防治得到學界的廣泛關注,具有非常重要的意義.然而目前,關于赤擬谷盜OBPs的組織分布情況,不同齡、性別中的不同蛋白表達差異研究仍不足,有待進一步考察,而分布情況及差異不僅在對赤擬谷盜防治工作中,也在天然植物揮發(fā)性物質(zhì)對赤擬谷盜的驅(qū)避、觸殺作用機制研究中非常重要.如何選擇合適的物質(zhì)在特定的時間段對赤擬谷盜進行防治,影響其繁殖等行為,以達到最佳防治效果,值得學界對赤擬谷盜OBPs進一步研究,也同樣對其他種害蟲防治具有借鑒意義.
在采用天然植物揮發(fā)油對赤擬谷盜的驅(qū)避、觸殺、熏蒸及胃毒等研究實驗中,針對赤擬谷盜OBPs通過何種機制影響赤擬谷盜的行為的研究仍存在不足.PBPs是OBPs家族中重要一員,在昆蟲繁衍過程中起關鍵作用,然而實驗中亦發(fā)現(xiàn)PBPs也可能參與到昆蟲的防御機制中,那么,PBPs如何參與到赤擬谷盜防御機制中,亦需要進一步的研究證實.是否能利用PBPs對昆蟲進行防治,抑制其繁殖從而對害蟲進行防治,保護儲糧安全,及關于PBPs的生理功能和意義亦值得學界進一步研究探索.
當前針對OBPs的研究方法,多數(shù)集中在對昆蟲離體蛋白的研究,而離體蛋白盡管在結(jié)構(gòu)上可以保持與體內(nèi)蛋白一致,但是二者的生理功能是否存在差異仍有疑點,受到學界的質(zhì)疑.若能采用體內(nèi)蛋白進行實驗可能是更好更具有說服力的一種方法.將計算機系統(tǒng)模擬實驗與體內(nèi)外實驗相結(jié)合,亦是一種較好的實驗方式.采用計算機系統(tǒng)模擬實驗對蛋白與配體進行篩選,對蛋白與配體結(jié)合時的作用力進行理論分析,可大幅度減少實驗成本,節(jié)省時間,同時也為體內(nèi)外實驗提供一些理論支持.
通過研究昆蟲OBPs有助于深入理解昆蟲化學感器,更好地理解昆蟲的自然行為,從而創(chuàng)造更高的經(jīng)濟效益,篩選出環(huán)保與經(jīng)濟并重的化學驅(qū)避劑或天然植物驅(qū)避劑.同時,OBPs作為載體蛋白,具有識別、運輸氣味分子的能力,因此對OBPs的研究不應僅局限于害蟲的防治,還可應用于揮發(fā)性物質(zhì)識別裝置,具有廣闊的發(fā)展前景.
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
中學生數(shù)理化·中考版(2022年11期)2022-02-16 07:01:20
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
發(fā)明與創(chuàng)新(2016年38期)2016-08-22 03:02:52
