基于線粒體基因組的鲹科魚類進化分析
2022-04-28 03:09:38王海山蔣歡陳治楊超杰葉樂
水產學雜志 2022年2期
王海山 ,蔣歡 ,陳治 ,楊超杰 ,葉樂
(1.海南熱帶海洋學院水產與生命學院,海南 三亞 572022;2.寧波大學海洋學院,浙江 寧波 315211)
鲹科(Carangidae)魚類隸屬硬骨魚綱(Osteichthyes)、輻鰭魚亞綱(Actinopteri)、鱸形目(Perciformes),下有4 個亞科:鲹亞科(Caranginae)、鰤亞科(Seriolinae)、鯧鲹亞科(Trachinotinae)和鰆鲹亞科(Chorineminae)。該科魚類廣泛分布于印度洋、太平洋、大西洋的熱帶和亞熱帶水域。現記錄的鲹科魚類共34 屬約147 種,其中鲹亞科的種類最豐富,共20 屬,占全科的50%以上[1]。中國海域有鲹科魚類64 種,分布于東海、黃海以及南海,全部鲹科魚類在南海均有分布,東海分布約25 種。每到春夏季,魚群會隨暖流從低緯度區洄游到高緯度出生地產卵,也有少數幾種能夠進入淡水流域。大部分鲹科魚類喜歡集群游動于沿岸水域或珊瑚礁海域[2],其中體型中等的種類游速較快。根據鲹科魚類體形特點和游動時的運動學特征,常把其游動模式運用于水下推進器的研究[3]。鲹科魚類多是重要的海洋經濟魚類,如竹筴魚(Trachurus trachurus)、細鱗圓鲹(Decapterus macarellus)和長體圓鲹(Decapterus macrosoma)等是南海外海漁業的重要捕撈對象[4],而五條鰤(Seriola quinqueradiata)、黃條鰤(Seriola lalandi)和卵形鯧鲹(Trachinotus ovatus)等高級食用海水魚,也是目前我國海南、廣東和廣西深水抗風浪網箱養殖重要種類。為了促進我國海水養殖業的發展,不同學者對多種鲹科魚類人工繁殖、網箱養殖技術、營養生理等展開了研究,并取得了突破性的進展[5-7]。
關于魚類分類,目前也做了許多工作。朱元鼎和鄭文蓮(1958)[8]描述了在中國南海海域發現的鈍鰭葉鲹(Caranx pectoralis)和海南鰆鲹(Chorinemus hainanensis)兩個新種的外部形態;Suzuki(1962)[9]比較了日本海域不同鲹科魚類的骨骼系統,探討了鲹科魚類的系統問題;鄭文蓮(1981)[10,11]采用傳統形態測量和解剖學方法,從骨骼、耳石、鱗片、嗅覺器官、消化器官等系統研究了中國海域鲹科魚類分類、分布和形態,為推進后續相關魚類科、亞科、屬、種的鑒別提供了重要參考資料;歐利國等(2020)[12,13]等基于地標點法鑒定了南海海域4 種鲹科魚類的108 尾樣品,同時依據耳石形態特征聚類分析了南海東沙群島海域7 種鲹科魚類,并建立了系統分類檢索表,為南海鲹科魚種識別以及分類提供了參考資料。總體而言,從18 世紀60 年代開始,關于鲹科魚類分類是以新種描述、形態分類、生物地理分類為主。
隨著生物信息統計軟件以及線粒體DNA(mitochondrial DNA,mtDNA)特征被普遍應用于各種生物的系統發育分析,分子生物學在鲹科魚類的研究中逐步開展。牛素芳等(2018)[14]分析了203 尾藍圓鲹的線粒體DNA 細胞色素b 序列的遺傳多樣性和遺傳結構,研究了南海北部藍圓鲹的群體遺傳變異特征;許則灘等(2015)[15]探討了東海舟山群島普陀海域18 尾鲹科樣本的線粒體COI 序列,通過比較種間和種內差異研究了本海域鲹科魚類的分類地位及分子系統進化關系,得出了鲹科魚類同一屬內種間遺傳距離明顯大于種內遺傳距離的結論,并認為普陀海域的18 尾鲹分別屬于7 屬8 種;朱世華等(2007)[16-18]結合數據庫已測定完成的3 種鲹科魚類序列,依據線粒體DNA 控制區、細胞色素b 序列、16S rRNA 序列生成矩陣和構建的分子系統樹,分析了中國海域的8 屬9 種鲹科魚類,結果支持鲹科四個亞科的分類階元,也提出獅鼻鯧鲹(Trachinotus blochii)為卵形鯧鲹的誤鑒,以及將麗葉鲹(Caranx kalla)的中文名改為“麗副葉鲹”的觀點。該結論與國外魚類分類學者David 等(2002)[19]基于形態特征分析和分子數據構建的部分鲹科魚類的分子發育樹相似。
魚類線粒體基因組不僅是研究真核細胞核酸與蛋白質合成的模型系統,也是研究DNA 結構與復制的良好模型。線粒體基因組為母性遺傳,進化主要是以堿基替換為主,進化速度較核DNA 快5~10 倍[20],被廣泛應用于魚類群體遺傳結構分析和類群識別。目前,盡管16S rRNA、D-loop 區和COI序列等片段分別被運用于鮭科、鯉科、鯛科和鳀科等魚類遺傳多樣性的研究中,但是,多認為線粒體上不同的區域進化速度不同,用序列更長的線粒體DNA 序列來構建統發育樹可以減少信息分布的不均一性和序列長度太短而降低系統進化樹的置信度帶來的影響,因此其結果更精確、可靠。
本文從GenBank 數據庫中下載已測定完成的33 種鲹科魚類的線粒體基因組序列,通過Clustal X和MEGA X 軟件進行DNA 序列比對和核酸多態性、遺傳距離等分析,結合鄰接法(Neighbor-Joining,NJ)、最小進化法(Minimum Evolution,ME)系統發育樹,探討鲹科種屬間的序列差異及其親緣關系,為鲹科的分類提供新的視角。
1 材料與方法
1.1 全基因組序列信息
通過檢索NCBI 的GenBank 數據庫獲取33 種鲹科魚類的線粒體基因組序列,并保存為FASTA格式。其種名、分類地位及相應序列號等相關信息見表1。
1.2 數據分析
將下載的基因組序列用Clustal X 軟件進行序列比對,再輔以人工校對;運用MEGA X 軟件統計序列的全長、堿基含量、變異位點數(比例)及簡約位點(Parsimony informative sites)等信息(表2)。多數學者認為Kimura 雙參數模型(Kimura2-Parameter,K-2-P)是評估低遺傳變異序列的最佳核酸替代模型,本研究利用MEGA X 軟件中Kimura 雙參數法計算種屬間的遺傳距離;利用DnaSP 5.0 軟件統計33 種鲹科魚類線粒體基因組序列的堿基多態性;運用MEGA 軟件中的鄰接法及最小進化法構建33 種鲹科魚類的系統進化樹,采用Bootstrap 法檢驗各分支的支持率,以自展值為1 000 次做重復抽樣檢驗,其余參數為軟件默認值。
2 結果與分析
2.1 核苷酸組成
序列分析顯示,鲹科魚類線粒體基因組序列全長在16 530~16 609 bp 之間,與大多數硬骨魚類線粒體序列長度相似[21],其中鰤屬的高體鰤(Seriola dumerili)序列最短,黃條鰤的序列最長。核苷酸多樣性指數為37.3%,檢測到序列總的變異位點共7072個,占42.0%,簡約信息位點共6 297 個,變異位點數大于簡約信息位點數。轉換/ 顛換比值(R)1613/1026=1.6<2.0,說明基因序列的突變達到飽和;同屬的個體堿基差異為1.68%~12.89%,平均差異為6.75%,表明鲹科魚類種間的堿基組成差異很小,呈現一定的核酸組成穩定性。線粒體全基因組堿基平均含量由高到低依次為C(29.5%)>A(27.8%)>T(26.0%)>G(16.7%),其中A+T 的含量(53.8%)大于G+C 含量(46.2%),表現出AT 含量相對豐富和GC含量相對缺乏的現象,符合硬骨魚類線粒體堿基組成偏好于A 和T 的特點[22-24]。本文所選全部物種堿基均有不同程度的AT 偏向(表1)。
DnaSP 5.0 計算位點多態性,發現了715 個單一變異位點、5 943 個簡約信息位點,其中發生種堿基突變的單一變異位點676 個、簡約信息位點3 120個,發生三種堿基突變的單一變異位點36 個、簡約信息位點1 363 個,發生四種堿基突變的單一變異位點3 個,簡約信息位點1 460 個。DnaSP 5.0 計算表明,33 種鲹科魚類線粒體基因組核酸多樣性(Pi)為0.15549,以100 bp 為單位在總長約16 800 bp 的線粒體基因組上步移,計算獲得核酸多樣性曲線(圖1),并結合鲹科魚類線粒體基因結構圖分析可以發現,33 種鲹科魚類線粒體基因組在D-Loop 區域具有最高的核酸多樣性(第15 861~16 072 位堿基核酸多樣性指數Pi 為0.48081),其次為ND6 和ND5 區域(核酸多樣性指數Pi 分別為0.26080 和0.26313),ND2 和ND1 區域也具有較高的核酸多樣性指數(0.23428 和0.23282)。16S rRNA、tRNA 區域核酸多樣性較低(最低為0.005)。即在33 種鲹科魚類線粒體基因組上堿基突變頻率最高的區域出現在線粒體復制D-Loop 控制區和NADH 基因的亞基位置。16S、12S rRNA 和tRNA 位置堿基突變頻率較低,基因較為保守。
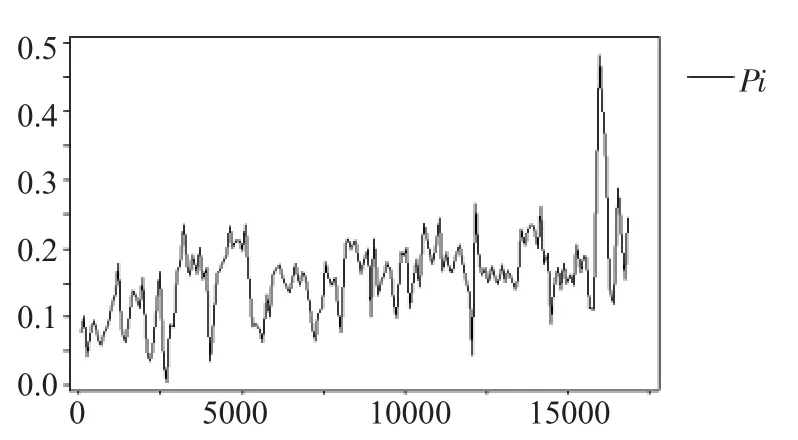
圖1 33 種魚線粒體基因組核酸多樣性曲線Fig.1 Mitochondrial DNA diversity of 33 fish species
2.2 遺傳距離矩陣
遺傳距離通常反映遺傳多樣性的高低,物種內個體之間的遺傳距離大,表明個體之間的親緣關系遠,物種間遺傳多樣性較高;相反的,物種內個體之間的遺傳距離小,表明個體之間的親緣關系近,遺傳多樣性較低。K-2-P 計算的遺傳距離顯示,鲹科魚類種間遺傳距離平均值為0.183;種間遺傳最大值為卵形鯧鲹與高體若鲹(Carangoides equula)的0.250,最小值為黑紋小條鰤(Seriolina nigrofasciata)與白舌尾甲鲹(Uraspis helvola)的0.002。其中卵形鯧鲹與鯧鲹屬外物種的遺傳距離最大,平均值為0.229;北美鯧鲹(Trachinotus carolinus)與鯧鲹屬外物種的平均遺傳距離為0.228;布氏鯧鲹(Trachinotus blochii)與鯧鲹屬外物種的平均遺傳距離為0.226;泰勃圓鲹(Decapterus tabl)與其他鲹科魚類平均遺傳距離最小為0.170;細鱗圓鲹與其他鲹科魚類平均遺傳距離為0.172;絲鲹(Alectis ciliaris)與其他鲹科魚類平均遺傳距離為0.175。值得注意的是,高體若鲹與若鲹屬其他種的遺傳距離并不是最小的,其與甲若鲹(Carangoides armatus)、馬拉巴若鲹(Carangoides malabaricus)、竹筴魚、藍圓鲹、日本竹筴魚(Trachurus japonicus)和泰勃圓鲹的遺傳距離分別為0.184、0.179、0.156、0.161、0.159 和0.158;與細鱗圓鲹、珍鲹(Caranx ignobilis)、烏鲹(Parastromateus niger)的遺傳距離均小于與馬拉巴若鲹的遺傳距離(表2)。

表2 33 種魚線粒體基因序列的種間遺傳距離Tab.2 Genetic distance of 33 fish species based on mitochondrial genome
綜上,鯧鲹屬在進化上與其他鲹科魚類分化較大,圓鲹屬與其他鲹科魚類遺傳距離更近,在線粒體基因組水平上高體若鲹與圓鲹屬魚類親緣關系更近,不符合傳統分類地位。
2.3 系統發育樹分析
早期對鲹科魚類的分類主要以外部形態為標準,由于地區、發育階段和環境的影響,相關研究中的特征描述或多或少存在差異,因而對鲹科魚類各屬種的分類存在爭議。本文分別采用NJ 法及ME法構建了基于線粒體基因組核苷酸序列的33 種鲹科魚類系統進化樹。Bootstrap 的參數值大于70%時,其結果符合真實的系統發育史的可靠性大于95%;如果參數值小于50%,則構建的系統發育樹無法反映實際情況。本實驗所建的NJ、ME 系統發育樹的絕大部分支的Bootstrap 都是100%,表明本研究可靠地反映了鲹科魚類的系統發育史(圖2、圖3)。
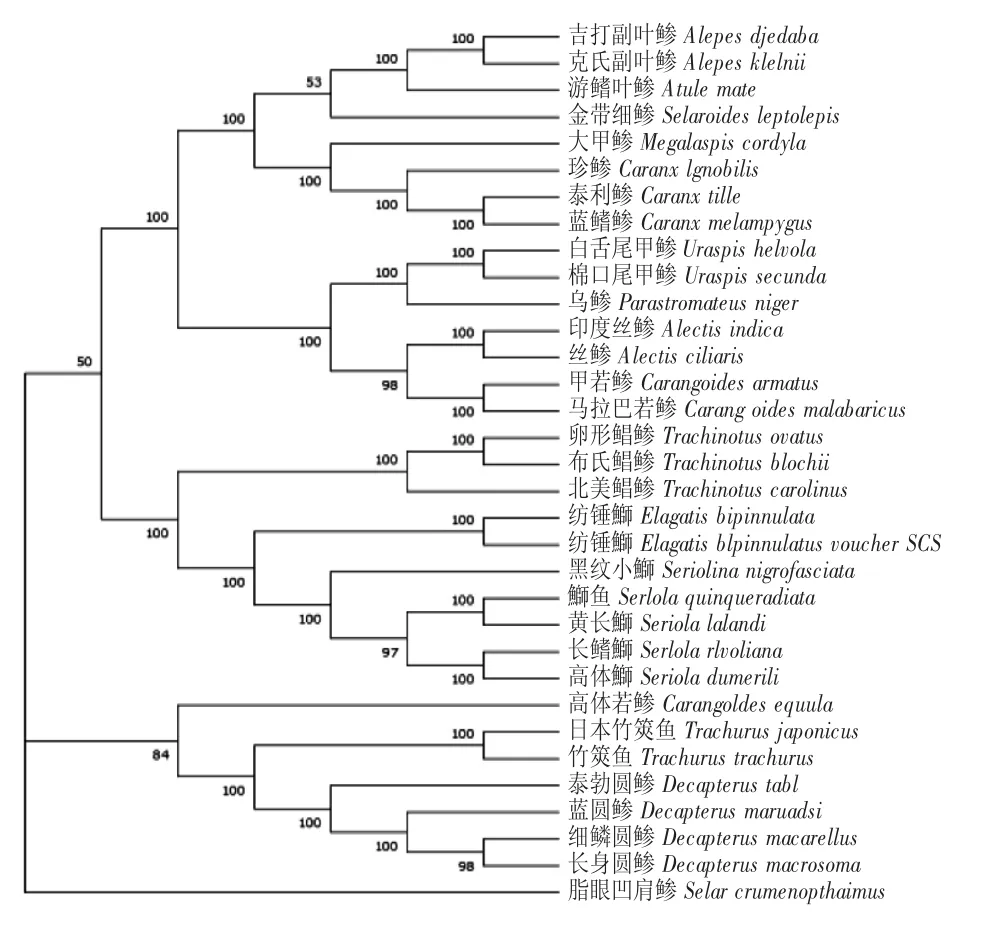
圖2 33 種魚基于線粒體序列的NJ 系統發育樹Fig.2 Phylogenetic trees of 33 fish species based on the complete mitochondrial genome by NJ analysis
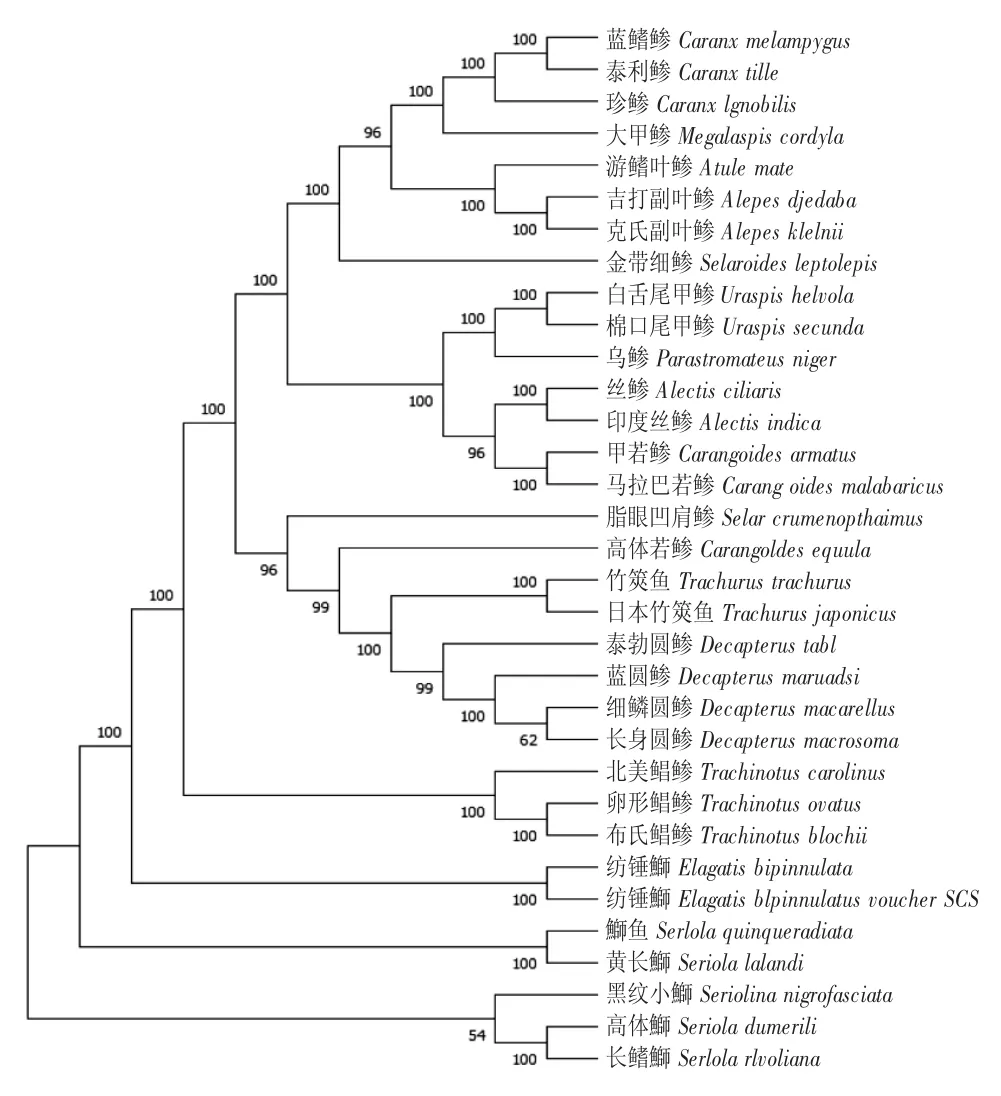
圖3 基于線粒體序列的ME 系統發育樹Fig.3 Phylogenetic trees based on the complete mitochondrial genome by ME analysis
NJ 樹顯示:鲹科魚類分為3 大支系,從樹的基部第1 支系可知鲹亞科的脂眼凹肩鲹(Selar crumenophthalmus)是較早單獨分離出來的一支;第2支系是竹筴魚屬與圓鲹屬形成姐妹群,再與高體若鲹聚為一支(置信度為50%);第3 支系由鰤亞科、鯧鲹亞科以及鲹亞科共同組成,較早分為2 個進化分支,一個分支顯示鯧鲹亞科的卵形鯧鲹、布氏鯧鲹以及北美鯧鲹所組成一個進化支,另一分支包含紡錘屬和鰤屬,再與鯧鲹屬聚集形成姐妹群。由此可知,鯧鲹亞科與鰤亞科有相當密切的親緣關系;樹的頂部為鲹亞科的魚類,包含副葉鲹屬、葉鲹屬、細鲹屬、大甲鲹屬以及鲹屬,每個屬都單獨成為一個支系;泰利鲹(Caranxtille)和藍鰭鲹(Caranxmelampygus)、棉口尾甲鲹(Uraspis secunda)和白舌尾甲鲹、吉打副葉鲹(Alepes djedaba)和克氏副葉鲹(Alepes kleinii)、竹筴魚和日本竹筴魚、長鰭鰤(Seriola rivoliana)和高體鰤、卵形鯧鲹和布氏鯧鲹這六組物種遺傳距離相同,進化速率相同;這種聚類關系與孫立元(2014)[25]的結果一致。
ME 樹顯示,鰤亞科進化距離最長,位于系群的基部,是鲹科的原始種群,其次是鯧鲹亞科;鲹亞科又分化出3 個分支,聚集到樹枝末端——物種組成與其在NJ 樹上所處的分支基本相同,只在排列順序上有差異,表明兩種方法所得到的結果一致。
3 討論
魚類線粒體基因組具有分子小、進化速度快、母系遺傳和信息相對穩定等優勢,適于從序列信息、基因組成及排列方式等角度來分析分子系統學及種群遺傳學[20]。本研究中,鲹科魚類線粒體基因組核苷酸序列的相關數據呈現AT 含量相對豐富和GC 含量相對缺乏的現象,符合硬骨魚類線粒體堿基組成偏好于A 和T 的特點。以NJ 法和ME 法構建的鲹科3 亞科15 屬33 種魚類基因序列的系統進化樹結果基本一致,均表明鯧鲹亞科與鰤亞科為姐妹群,這與朱世華等利用線粒體DNA 控制區、細胞色素b 序列、16S rRNA 序列分析結果一致。鰤亞科是3 個亞科中的最原始種群[10,11]。
早期的系統發育分析主要通過化石和現存物種的生理、形態特征來構建系統發育關系。荷蘭自然學家Coenraad 和Hermann(1989)[26]首先于1844年根據從日本海域采集的高體若鲹標本進行了體長及背鰭、臀鰭、胸鰭和腹鰭數等特征的描述。1844—1907 年間,高體若鲹先后被更名為Carangoides acutus、Kaiwarinusequula、Carangoidesequula、Carangoides dasson 等。高體若鲹最開始被歸為鲹屬下的若鲹亞屬,后因分類系統修改鲹屬上升為鲹亞科,其下亞屬也相應上升為屬,高體若鲹歸為鲹亞科的若鲹屬。目前高體若鲹為鲹亞科、若鲹屬的分類地位被廣為接受。較多文獻中竹筴魚、日本竹筴魚學名、中文名的使用也較為混亂。從物種的命名以及相關文獻的記錄都顯示,鲹科魚類中的某些種類分類地位存在爭議。本研究構建的兩種系統發育樹均顯示:若鲹屬的馬拉巴若鲹、甲若鲹與絲鲹屬的絲鲹以及印度絲鲹(Alectis indica)形成姐妹群,而若鲹屬的高體若鲹并未與甲若鲹、馬拉巴若鲹聚為一支,而是與圓鲹屬、竹筴魚屬的種類聚為1 支(置信度84%~99%),表明高體若鲹與圓鲹屬、竹筴魚屬之間的親緣關系較近,與若鲹屬間的親緣關系較遠。本研究結果與史寶等(2019)[27]在探討黃條鰤線粒體基因組系統進化分析中所構建的系統進化樹結果基本一致,但后者并未討論高體若鲹處于的異常分支的結果。
以往分析鲹亞科時主要基于體形、肩帶下角有無深凹、上下頜有無牙齒、牙齒形狀、牙列數、第二背鰭有無延長鰭條等形態學特征進行分類。如歐國利和劉必林(2020)[12]利用地標點法對鲹科魚類耳石形態進行分類識別,將高體若鲹與短吻絲鲹、竹筴魚以及脂眼凹肩鲹區分開來;從外部形態學分析,高體若鲹與馬拉巴若鲹、甲若鲹都具有體形為卵圓形、高而側扁的形態特征,但卻在一定程度上受性別、發育階段以及適應環境的影響。如幼魚時,甲若鲹體側與高體若鲹同樣具六條暗色橫帶,但隨著個體成長,甲若鲹的暗色橫帶逐漸消失。因此,形態學鑒定在一定程度上受性別、發育階段以及適應環境的影響,僅依靠形態學對物種分類鑒定存在著較多問題。
