姜黃素對高糖誘導的人視網膜血管內皮細胞增殖及VEGF和NF-κB p65表達的抑制作用
2022-05-05 13:21:00潘穎喆謝安琪
國際眼科雜志 2022年5期
關鍵詞:實驗
項 彪,潘穎喆,謝安琪,程 希,鄧 玲
0引言
糖尿病視網膜病變(diabetic retinopathy,DR)是糖尿病最常見的慢性微血管并發癥之一。DR導致視力下降甚至失明最主要的階段是增殖性糖尿病視網膜病變(proliferative diabetic retinopathy,PDR)。流行病學調查表明,DR診斷25a后,PDR的患病率接近50%,且大多數1型糖尿病(T1DM)患者將在大約10a后發展為PDR[1],其病理學特征主要為晚期組織缺血缺氧,誘導多種促血管生成因子和炎癥因子的釋放增加,導致新生血管的形成,從而繼發玻璃體積血和視網膜脫離引起患者視力喪失[2]。在DR的發生發展進程中,VEGF和NF-κB起著非常關鍵的作用。其中,血管內皮生長因子(vascular endothelial growth factor,VEGF)被認為是促進血管生成的最主要因子,也是抗血管生成的重要靶點[3]。研究發現,PDR患者玻璃體和纖維血管組織中的VEGF水平顯著高于正常眼[4]。核因子-κB(nuclear factor-κB,NF-κB)是一種多向性核轉錄因子,廣泛參與機體的免疫、炎癥等生理病理過程[5]。研究發現,DR高血糖狀態可引起活性氧(reactive oxygen species,ROS)生成增加,從而介導大量的炎癥因子釋放,而炎癥因子和ROS水平上調又可進一步激活NF-κB,導致視網膜低度炎癥[6]。姜黃素(curcumin,Cur)是一種從姜科植物姜黃中提取的主要的多酚類活性成分,具有抗氧化、抗炎、抗腫瘤等多種藥理作用,尤其在抗腫瘤方面具有較強的功效,能通過下調VEGF的表達抑制腫瘤新生血管的形成[7-8]。研究還發現,姜黃素顯著降低脂質過氧化,增加細胞內抗氧化劑含量,調節抗氧化酶,并清除高血糖誘導的ROS產生[9]。但其對于高糖誘導的人視網膜血管內皮細胞(human retinal capillary endothelial cells,HRCECs)是否也能通過降低VEGF、NF-κB的表達而發揮治療作用尚不明確。因此,本研究通過CCK-8細胞增殖實驗、Western-blot及免疫細胞化學法,探討姜黃素對于高糖誘導的HRCECs作用的潛在機制。
1材料和方法
1.1材料HRCECs(上海中科院細胞庫);低糖/高糖DMEM培養基(美國Gibco公司);姜黃素、胰酶(美國Sigma公司);CCK-8細胞增殖試劑盒(上海碧云天生物公司);兔抗人VEGF多克隆抗體,兔抗人NF-κB多克隆抗體,兔抗人血管內皮生長因子IgG,蛋白預染Marker,BCA蛋白濃度測定試劑盒(武漢博士德生物公司);倒置相差顯微鏡(日本Olympus公司);全自動酶標儀(美國Bio-Rad公司)。本研究經襄陽市第一人民醫院醫學倫理委員會審核通過。
1.2方法
1.2.1姜黃素溶液的配置姜黃素,純度>95%,稱取14.2mg姜黃素粉末溶解于1mL 100%的二甲基亞砜(DMSO)溶劑中,根據C=m/M.V,配制成濃度為40mmol/L的姜黃素母液,除菌分裝后保存在-20℃冰箱中。臨用前,用DMEM高糖培養基稀釋至實驗所需濃度,DMSO終濃度<0.1%。
1.2.2細胞處理及分組將HRCECs置于含有青/鏈雙抗、胎牛血清的DMEM培養基中,放入培養箱中培養。取處于對數生長期的細胞進行實驗分組:空白組(G0組,只含高糖DMEM培養基的調零組),正常對照組(NG組,低糖DMEM培養基+細胞),高糖對照組(HG組,高糖DMEM培養基+細胞),G1組(高糖DMEM培養基+細胞+20μmol/L姜黃素),G2組(高糖DMEM培養基+細胞+40μmol/L姜黃素),G3組(高糖DMEM培養基+細胞+80μmol/L姜黃素)。正常糖濃度的DMEM培養基葡萄糖濃度為5.5mmol/L,統稱為低糖培養基;高糖DMEM培養基中葡萄糖濃度為25mmol/L;G0組僅設于CCK-8實驗。
1.2.3CCK-8法檢測HRCECs增殖取對數生長期且長勢良好的HRCECs用于實驗,將細胞經PBS洗滌、胰酶消化、離心后,用DMEM培養基制成濃度為5×104cell/mL的單細胞懸液,以每孔200μL將懸液接種于96孔板內,每組設5個復孔,培養至細胞貼壁。按照上述分組分別培養12、24、48h,向每孔加入CCK-8溶液10μL,37℃孵育4h后,用酶標儀在450nm波長下測定各孔吸光度值(A值),計算每組濃度復孔平均值及細胞增殖抑制率,上述實驗重復3次。細胞抑制率(%)=[(對照組平均A值-加藥組平均A值)/(對照組平均A值-空白組平均A值)]×100%。
1.2.4Western-blot法檢測HRCECs中蛋白的表達收集各組處理24h后的HRCECs用PBS洗滌,加入適量的RIPA裂解液4℃下反應20min以提取細胞總蛋白。加入1/5體積的5×蛋白上樣緩沖液,置于100℃沸水中10min。BCA法檢測蛋白濃度。按照每孔50μg上樣量進行SDS-PAGE電泳。恒流300mA電轉90min后,用5%的脫脂牛奶室溫下封閉60min。然后分別加入經TBST液稀釋的VEGF、NF-κB和β-actin一抗4℃孵育過夜,洗膜后加入辣根過氧化物酶標記的二抗室溫下孵育1h。采用Image J軟件測定灰度值,計算A值,以β-actin為內參計算蛋白相對表達量。
1.2.5免疫細胞化學法檢測HRCECs中蛋白的表達將預先消毒滅菌的蓋玻片放入6孔板內,制備濃度為5.0×104cell/mL的單細胞懸液,每孔內滴入2mL懸液,培養48h后棄液,按照分組加入不同的條件培養液,共5組:NG組、HG組、G1~G3組,每組設2個復孔,繼續培養24h后取出蓋玻片,用PBS洗滌、4%多聚甲醛室溫固定15min。再經PBS洗滌,0.1%Triton-X100室溫通透化處理15min作染色前處理。然后在3%過氧化氫-甲醇溶液中浸泡30min,以滅活內源性過氧化物酶,用PBS液洗滌后,再將其置于二抗來源的非免疫兔血清濕盒中封閉30min,去除血清后甩干;加入一抗(兔抗人VEGF 1∶200),同時用PBS代替一抗設為陰性對照組,4℃濕盒中過夜,室溫下復溫30min,PBS液洗滌,加入生物素標記的二抗37℃孵育1h。最后,經SABC法染色、DAB顯色、蘇木素復染、乙醇梯度脫水、二甲苯透明、中性樹膠封片,干燥后顯微鏡下觀察。上述實驗重復3次。計算各組標本灰度值:陽性灰度值與背景灰度值之差。
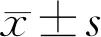
2結果
2.1CCK-8法檢測姜黃素對高糖誘導的HRCECs增殖能力的影響干預不同時間各組A450比較,差異有統計學意義(F組間=687.00,F時間=179.50,F交互=26.74,均P<0.001)。與NG組相比,HG組中HRCECs增殖能力顯著增強(P<0.01),隨著干預時間的延長而增強。增殖抑制率隨著藥物的濃度增加而上升,具有濃度依賴性。在相同藥物濃度作用下,隨著干預時間的延長,細胞增殖能力減弱(P<0.01),細胞增殖抑制率逐漸上升,具有時間依賴性。結果表明,高糖環境下的HRCECs增殖能力顯著增強,而姜黃素對高糖誘導的HRCECs增殖有明顯抑制作用,且隨著濃度的增加、干預時間的延長抑制作用增強,見表1,圖1。

表1 不同濃度姜黃素作用不同時間后對高糖誘導的HRCECs增殖的影響

圖1 不同濃度姜黃素作用不同時間后對高糖誘導的HRCECs增殖抑制率的影響。
2.2Western-blot法檢測姜黃素對高糖誘導的HRCECs中VEGF-A和NF-κBp65表達的影響實驗結果顯示:與NG組比較,HG組中VEGF-A、NF-κB p65的表達顯著升高(P<0.01);與HG組比較,G1~G3組中VEGF-A、NF-κB p65的表達均顯著下降(P<0.01);在相同藥物濃度作用下,隨著干預時間延長,HRCECs中VEGF-A、NF-κB p65的表達呈下降趨勢(P<0.01);在相同干預時間下,HG組與G1~G3組之間兩兩比較,隨著姜黃素濃度增加,HRCECs中VEGF-A、NF-κB p65的表達呈下降趨勢(P<0.01)。結果表明,姜黃素對高糖誘導的HRCECs中VEGF-A、NF-κB p65的表達具有下調作用,且在藥物濃度范圍內呈劑量-時間依賴性,見表2、3,圖2~4。

表2 高糖誘導的HRCECs經不同濃度姜黃素干預12、24、48h后VEGF-A的表達

圖2 姜黃素作用12h后對高糖誘導的HRCECs中VEGF-A和NF-κB p65表達的影響。

圖3 姜黃素作用24h后對高糖誘導的HRCECs中VEGF-A和NF-κB p65表達的影響。

圖4 姜黃素作用48h后對高糖誘導的HRCECs中VEGF-A和NF-κB p65表達的影響。
2.3免疫細胞化學法檢測姜黃素對高糖誘導的HRCECs中VEGF表達的影響VEGF的表達主要存在于HRCECs的細胞膜及細胞漿中,陽性表現為棕褐色顆粒著染。灰度值結果示,與NG組比較,HG組HRCECs中VEGF的表達明顯增多(P<0.01)。不同濃度姜黃素干預24h后與HG組比較,HRCECs中VEGF表達明顯減少(P<0.01),隨著藥物濃度增加,細胞膜及細胞漿中的棕褐色陽性著染逐漸減少,顏色變淡,不同濃度加藥組之間兩兩比較,差異均有統計學意義(P<0.01)。表明姜黃素能有效抑制HRCECs中VEGF的表達,且隨著藥物濃度增加抑制作用增強,上述結果與Western-blot結果相符,見表4、圖5。

圖5 免疫細胞化學法檢測姜黃素對高糖誘導的HRCECs中VEGF表達的影響。
3討論
視網膜新生血管(retinal neovascularization ,RNV)是DR患者病情惡化、視力喪失的主要原因,其發生發展與HRCECs的增殖、VEGF及NF-κB表達上調之間關系密切。
表3 高糖誘導的HRCECs經不同濃度姜黃素干預12、24、48h后NF-κB p65的表達

表4 姜黃素作用24h后各組HRCECs中VEGF的表達
新生血管的形成過程十分復雜,主要包括血管基底膜降解、內皮細胞增殖、遷移、成管四個階段。研究發現,長期高糖環境對不同組織的血管內皮細胞影響不同,對大血管內皮細胞主要表現為抑制其生長,而對微血管內皮細胞反而表現為促進其增殖[10]。本實驗結果也提示,高糖環境下,HRCECs的增殖能力顯著增強,與既往文獻報道相符[11];在給予不用濃度姜黃素干預后,可明顯降低HRCECs的增殖能力,且抑制作用隨著藥物濃度增加、干預時間延長而增強。表明高糖誘導的HRCECs增殖能力增強,而姜黃素對其可有效抑制。
參與調控RNV形成的細胞因子眾多,血管生成因子(如VEGF)起刺激作用,血管抑素和色素上皮衍生因子(PEDF)起抑制作用[12]。正常環境下,各種因子保持動態平衡。但在高血糖、氧化應激及非感染性炎癥反應等病理狀態時,平衡狀態會被破壞,促使血管生成的主要因子VEGF表達上調[13]。VEGF與其受體結合后,可激活下游的PI3K/Akt、p38及Raf等通路,來調控內皮細胞的增殖。本實驗中Western-blot和免疫細胞化學法結果均顯示,HG組細胞中VEGF的表達量較NG組顯著增加,表明一定濃度范圍內高糖環境可上調VEGF的表達,與既往文獻報道相符[11]。姜黃素在抗新生血管形成方面的作用已經得到了證實。研究發現,姜黃素能通過調控VEGF信號通路誘導氧化應激和抑制血管生成,在結直腸癌的治療上發揮作用[8]。李勇等[7]研究發現,姜黃素可通過降低VEGF的表達水平,抑制肝癌組織缺氧誘導的血管形成和肝癌細胞生長轉移。本實驗采用不同濃度姜黃素處理高糖誘導的HRCECs后發現,加藥組細胞VEGF的表達顯著下降,且呈濃度與時間依賴性。結合CCK-8結果,表明姜黃素抑制體外高糖誘導的HRCECs增殖可能與VEGF表達下調有關。
NF-κB是一種關鍵的促炎轉錄因子,其信號通路的激活是DR炎癥反應發生發展過程中的關鍵一步。在PDR患者中,高血糖介導的ROS生成增加、NF-κB激活與VEGF上調之間關系顯著[14]。研究發現,高糖環境下氧化應激誘導的ROS上調和非酶糖基化產物與其受體的結合,均能引起NF-κB的持續激活[6,15]。正常情況下,NF-κB在胞漿中與抑制蛋白(I-κB)結合處于失活狀態。當受到刺激時,I-κB激酶發生磷酸化,導致游離的NF-κB易位到細胞核,與特定的基因序列結合,誘導多種促炎因子、黏附因子、炎性酶、趨化因子等表達,從而促進炎癥發生、細胞增殖和血管生成[5],表明NF-κB的激活是始動事件,隨后其他炎癥通路的激活逐步導致了DR微血管損傷。研究還發現NF-κB通路的激活能誘導VEGF的表達[16]。姜黃素對NF-κB通路的抑制作用已在多種細胞中被報道。研究發現,姜黃素能通過抑制I-κB激酶和NF-κB活性誘導慢性粒細胞白血病細胞的KCL-22細胞凋亡[17],其機制可能是由姜黃素自身的抗炎、抗氧化等藥理特性改變了細胞的微環境和對ROS水平的調控。本研究中Western-blot結果示HG組細胞中NF-κB p65的表達量較NG組顯著增加,表明NF-κB通路在DR進展過程發揮著重要作用,與既往文獻報道相符[11]。在給予不用濃度姜黃素干預后,加藥組細胞NF-κB p65的表達顯著下降,且呈濃度與時間依賴性。結合CCK-8結果,表明姜黃素抑制體外高糖誘導的HRCECs增殖可能與NF-κB p65表達下調有關。
綜上所述,本實驗證實姜黃素能抑制高糖誘導的HRCECs增殖,其機制可能與其下調HRCECs中VEGF-A、NF-κB p65的表達有關。但本實驗僅是體外實驗,而且關于RNV的生成因素只考慮了VEGF和NF-κB,實際上DR的發病機制復雜,參與的細胞因子眾多,是多種因素相互作用的結果。本實驗尚未明確姜黃素抑制VEGF、NF-κB表達的具體機制,及它們間的相互作用關系。姜黃素對高糖誘導的HRCECs增殖及VEGF、NF-κB p65表達的抑制作用的具體機制等問題有待進一步實驗進行深入研究。
猜你喜歡
作文·小學低年級(2025年2期)2025-02-13 00:00:00
小雪花·小學生快樂作文(2024年11期)2024-12-31 00:00:00
作文·小學低年級(2024年2期)2024-04-29 00:00:00
作文·小學低年級(2023年3期)2023-04-29 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
小主人報(2022年4期)2022-08-09 08:52:06
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
