低溫脅迫對柳杉不同無性系的影響及抗寒性評價
2022-05-10 12:21:10崔潔冰張萌張瑩婷徐進
生物技術通報 2022年3期
關鍵詞:植物
崔潔冰 張萌 張瑩婷 徐進
(南京林業大學林木遺傳與生物技術省部共建教育部重點實驗室 南京林業大學南方現代林業協同創新中心 南京林業大學林學院,南京 210037)
柳杉(Cryptomeria fortunei)為杉科(Taxodiaceae)柳杉屬(Cryptomeria)常綠高大喬木,喜山地氣候及條件良好的酸性土壤,具有側根發達、生長迅速、適應性強、樹形優美且能凈化空氣、改善環境等特性[1-2],是中國南方優良的用材和園林樹種。天然種群集中分布于中國東南的天目山、武夷山、日本本州島及九州島等地,長江以南省區也多有栽培[3]。但其生長和分布常受到干旱、寒冷等極端環境的制約,隨著全球氣候變化以及極端天氣頻繁發生,柳杉的生長發育受到了極大的影響。
低溫是影響自然界植物生存的主要環境因子之一,也是限制植物地理分布的關鍵因素[4]。一般來說,溫度降低時會導致植物的各種生理活動減慢,生長發育變緩,當低溫造成的傷害超過一定程度時,環境脅迫就會對植株造成不可逆的傷害,甚至死亡[5-6]。植物為了抵御低溫脅迫,其根莖葉在形態上存在著相應的響應機制,如葉形、葉片厚度、葉色和解剖結構等方面的變化[7-8]。其次,植物體內多種滲透調節物以及保護酶等在維持細胞結構及生理功能、抵抗惡劣氣候條件等方面具有重要作用[9],并通過一系列生理生化代謝反應提高對環境的適應能力。
柳杉的抗寒性研究可為其品種選育提供理論依據,對于提高林木生產力與地區經濟效益、促進我國生態環境的建設具有重要意義[10]。但是目前柳杉的研究主要集中在育苗、良種選育、病蟲害防治與生理生態等方面,關于柳杉不同無性系的抗寒性研究相對較少。本研究測定了10個無性系在低溫脅迫下的丙二醛(malondialdehyde,MDA)含量、超氧化物歧化酶(superoxide dismutase,SOD)活性及葉綠素熒光參數(Fv/Fm、Y(II)、qP)等生理生化指標,采用主成分分析結合隸屬函數計算對柳杉各無性系抗寒性差異進行綜合評價,以期進一步探尋柳杉無性系抗寒能力評價體系,為柳杉抗寒無性系的評價篩選及造林栽培提供理論依據。
1 材料與方法
1.1 材料
10個柳杉無性系主要來源于我國東部柳杉分布區(表1),穗條均采自南京林業大學白馬實驗基地柳杉優良無性系收集區,2018年6月進行扦插,2020年用于本研究。

表1 材料及來源Table 1 Materials and sources
1.2 方法
1.2.1 低溫處理 取生長一致的10個柳杉無性系的2年生幼苗枝條,將每個無性系的枝條分為7組,分別放入海爾程控冰箱中進行4℃、0℃、-4℃、-8℃、-12℃、-16℃和-20℃低溫處理,從4℃開始,以4℃/2 h降溫,到達每個目標溫度時處理12 h。零上溫度處理后直接進行生理指標的測定,零下溫度處理結束后放入4℃中解凍12 h后進行測定,每個處理設置3個重復。
1.2.2 測定方法 取針葉0.5 g,通過0.05 mol/L磷酸緩沖液(pH7.8)獲得粗酶提取液。采用硫代巴比妥酸法[11]和氮藍四唑比色法[11]分別測定MDA含量和SOD活性。柳杉枝條經低溫處理后,使用便攜式光纖型雙通道調制葉綠素熒光儀DUAL-PAM/F(德國Walz公司)測定葉綠素熒光參數PS II最大光化學效率(Fv/Fm)、PS II的實際光合量子產量(Y(II))和光化學淬滅系數(qP),每個處理3次重復。
1.2.3 數據分析 (1)各項指標數據采用Excel 2010和SPSS 22進行統計與分析,使用Duncan檢驗法進行差異顯著性分析。
(2)權重計算:本課題組Zhang等[12]在分析半致死溫度時發現-4℃是一個關鍵的轉折溫度,因此以-4℃下各指標的值作主成分分析。綜合相對電導率(relative conductivity,REC)、MDA含量、SOD活性及Fv/Fm 4個指標(REC數據見Zhang等[12]),使用SPSS 22軟件將數據進行標準化處理,通過主成分分析確定其特征值、負荷量與貢獻值,計算權重(weight,W)。
(3)隸屬函數值計算:根據各無性系測定的指標數據,計算其隸屬函數值并進行排序,對負向指標采用反隸屬函數計算[13-14]。
隸屬函數 :f(xij)=(xij-xjmin)/(xjmax-xjmin)
反隸屬函數 :f(xij)=(xjmax-xij)/(xjmax-xjmin)
式中,f(xij)表示i無性系j指標的抗寒隸屬函數值;xij表示i無性系j指標的測定值;xjmax表示所有無性系j指標的最大值;xjmin表示所有無性系j指標的最小值。
(4)抗寒性評價:根據各指標權重及隸屬函數值,通過公式Y=Wij×f(xij)計算各無性系抗寒性綜合指數Y(Wij為各抗寒指標權重;f(xij)為各指標隸屬函數值)[13-14]。再結合相對電導率(REC)、MDA含量、SOD活性以及葉綠素熒光參數(Fv/Fm)等指標進行綜合分析,建立抗寒性評價體系。
2 結果
2.1 低溫脅迫對柳杉各無性系生理生化指標的影響
2.1.1 低溫脅迫對柳杉各無性系中MDA含量的影響 由圖1和附表1可知,柳杉不同無性系的MDA含量總體呈上升-下降-上升的趨勢。但柳杉各無性系在不同溫度時,MDA含量的變化不太一致,在一定溫度范圍內具有波動性。在0℃、-8℃、-16℃出現了峰值,在0℃和-16℃后呈明顯的下降趨勢,其中在-4℃、-8℃、-12℃時,MDA含量相較于0℃呈現下降趨勢,但各無性系變化無明顯規律,說明柳杉不同無性系對低溫的應答反應不同,且MDA含量受多種因素影響。在整個低溫處理過程中,66#無性系的MDA含量偏低,74#無性系的MDA含量偏高,說明66#無性系在低溫脅迫下膜功能受損害的程度較低,抗性較強。

圖1 不同低溫脅迫對柳杉各無性系MDA含量的影響Fig.1 Effects of different low temperature stress on MDA content in different clones of C. fortunei
2.1.2 低溫脅迫對柳杉各無性系超氧化物歧化酶(SOD)含量的影響 由圖2和附表2可知,在低溫脅迫下,多數柳杉無性系的SOD活性變化呈下降-上升-下降的趨勢。在4℃降到0℃時,由于植株對低溫的不適應,SOD活性呈現下降趨勢,隨后植株經過短暫的抗寒鍛煉,抗寒能力逐漸增強,SOD活性出現短暫的上升,-8℃后,SOD活性逐漸下降,說明過低的溫度超過了SOD酶作用的范圍,植株自身抗氧化系統受到破壞,抗寒能力減弱。特別的是25#和74#無性系,呈現先上升后下降的趨勢。在整個低溫處理過程中,68#無性系的SOD活性值偏高,而且波動較小,抗性較強。42#無性系的SOD活性值下降較快且降幅較大,說明在低溫處理過程中,42#無性系的超氧化物歧化酶受溫度的影響比較大,抗性較弱。

圖2 不同低溫脅迫對柳杉各無性系SOD含量的影響Fig.2 Effects of different low temperature stress on SOD content in different clones of C. fortunei
2.1.3 低溫脅迫對柳杉各無性系葉綠素熒光參數的影響
2.1.3.1 低溫脅迫對柳杉各無性系Fv/Fm的影響 由圖3和附表3可知,隨著處理溫度的降低,柳杉各無性系的Fv/Fm值呈上升-下降的變化趨勢。在0℃時,Fv/Fm開始下降,-4℃時急劇降低,到-20℃逐漸平緩,說明柳杉在經歷低溫初期時有抵御能力,能夠在一段時間內維持光合效率,超過抵御的臨界點,其光合效率大幅度降低。其中,42#無性系的Fv/Fm變化幅度較大,且顯著低于其他無性系,說明其PS II反應中心光能轉化效率較低,對低溫脅迫較為敏感,抗寒性較差。68#無性系的Fv/Fm值在4℃至-4℃時相對較高,而57#無性系在低溫初期變化平緩且隨溫度的降低整體水平降幅較小,說明它們在低溫脅迫下PS II損傷較小,光能轉化效率較高。

圖3 不同低溫脅迫對柳杉各無性系Fv/Fm的影響Fig.3 Effects of different low temperature stress on Fv/Fm in different clones of C. fortunei
2.1.3.2 低溫脅迫對柳杉各無性系Y(II)的影響 由圖4和附表4可知,在低溫處理下柳杉各無性系Y(II)與Fv/Fm的變化趨勢相似,隨著溫度的降低整體呈上升-下降的變化趨勢。Y(II)在低溫初期出現小幅上升,在0℃后呈下降趨勢,隨著溫度的降低,在-4℃以后急劇下降,-8℃后變化逐漸平緩,-20℃時各無性系Y(II)值趨近于0。其中,68#無性系在0℃達到峰值,在-4℃之后急劇下降,且變化幅度較大,說明68#在低溫初期抗性較強,卻對溫度較為敏感。57#無性系的Y(II)值隨溫度降低也明顯下降,但整體上比其他無性系下降幅度小。由圖可以看出,42#無性系在-8℃和-16℃時Y(II)值趨近于0,各無性系在-20℃時的Y(II)值也幾乎為0。Y(II)是PS II實際光合量子產量,一段時間內持續地對光能的有效轉換能力[15]。Y(II)為0,說明柳杉PS II反應中心的實際光能轉換能力在低溫脅迫下受到抑制,柳杉的光合結構受到破壞。

圖4 不同低溫脅迫對柳杉各無性系Y(II)的影響Fig.4 Effects of different low temperature stress on Y(II)in different clones of C. fortunei
2.1.3.3 低溫脅迫對柳杉各無性系qP的影響 由圖5和附表5可知,柳杉各無性系qP值在4℃-0℃無顯著變化,從0℃開始緩慢下降,-4℃時qP值急劇降低,在-8℃后變化平緩,直至-20℃ qP值趨近于0,這與Y(II)值的變化相似。由圖5可以看出,qP值隨溫度的降低呈現下降趨勢。其中,74#無性系的qP值在低溫初期變化不明顯且處于較高水平,57#和32#無性系隨溫度的變化qP值降幅較小,在低溫后期,qP值高于其他無性系,說明74#、57#和32#無性系光合電子傳遞能力較強,PS II受害程度較輕,抗性較好。42#無性系qP值變化與Y(II)值變化相似,顯著低于其他無性系且趨近于0,說明對光能的利用能力較弱,抗性較差。

圖5 不同低溫脅迫對柳杉各無性系qP的影響Fig.5 Effects of different low temperature stress on qP in different clones of C. fortunei
2.2 主成分分析
結果如表2所示,遵循特征值大于1的原則,從4項生理指標中共提取出2個主成分,特征值分別為2.217和1.056,貢獻率分別為55.419%和26.389%,累計貢獻率達81.809%,因此,結果理想,提取的2個主成分概括了4個生理指標的絕大部分信息。運用2個主成分對4個生理指標進行負荷量分析和權重的計算,由表3可以看出,各指標對柳杉抗寒性影響由強到弱分別是相對電導率(REC)、MDA、Fv/Fm、SOD。

表2 主成分分析結果Table 2 Results by principal component analysis
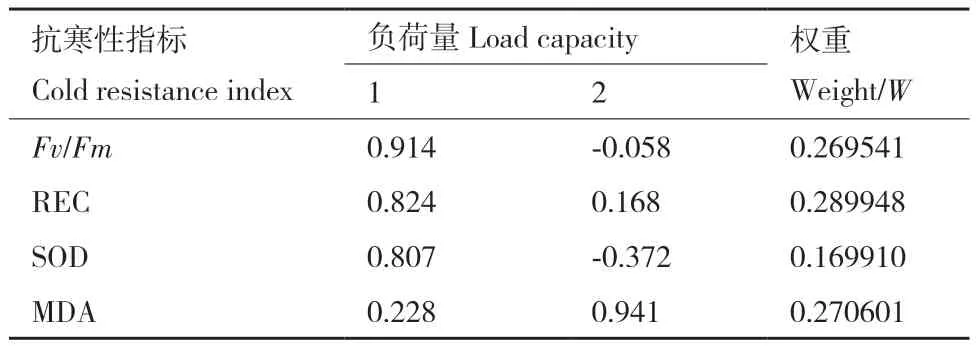
表3 柳杉各無性系抗寒性的負荷量及權重Table 3 Load capacities and weights of cold resistance indicators of C. fortunei clones
2.3 抗寒指標隸屬函數值
以相對電導率(REC)、MDA含量、SOD活性及葉綠素熒光參數(Fv/Fm)為評級指標,計算隸屬函數值并進行排序。結果(表4)表明,無性系57#、68#和32#的隸屬函數值較大,表明其抗寒評分較高,抗寒性較強,3#、25#和42#無性系隸屬函數值較小,表明其抗寒評分較低,抗寒性相對較弱。
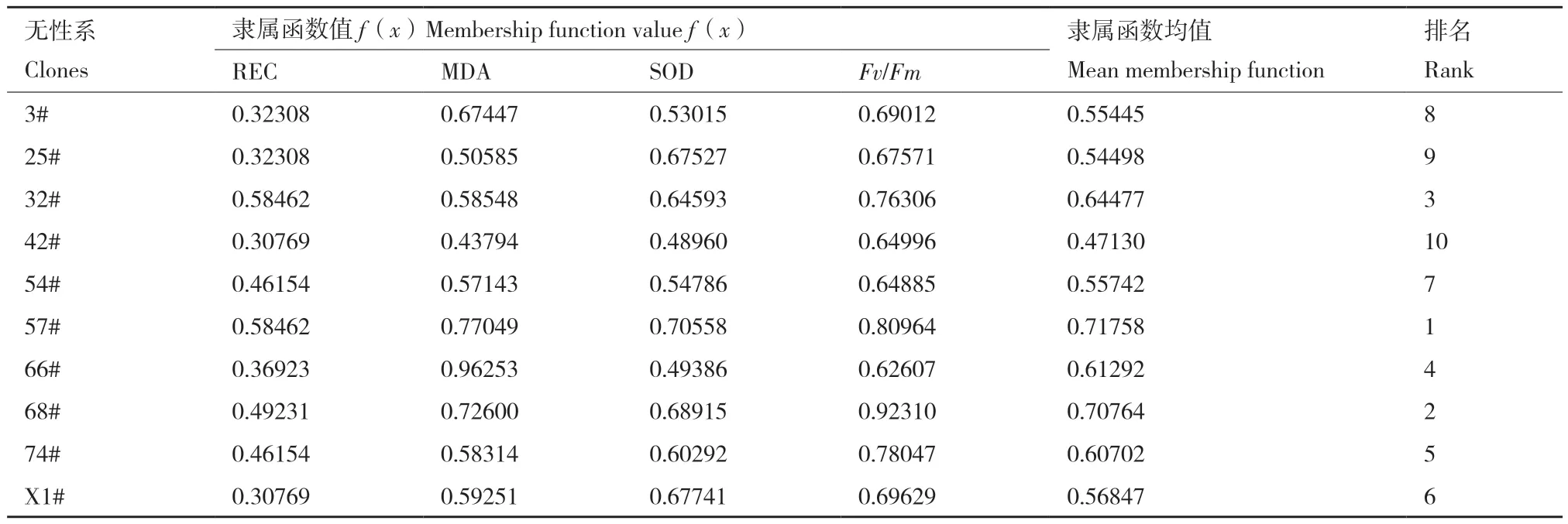
表4 柳杉各無性系抗寒性指標的隸屬函數值Table 4 Membership function values of cold resistance index of C. fortunei clones
2.4 柳杉各無性系抗寒性綜合評價
以相對電導率(REC)、MDA含量、SOD活性及葉綠素熒光參數(Fv/Fm)為評級指標,依據權重與隸屬函數值,計算柳杉各無性系抗寒性綜合指數Y,進而評價柳杉各無性系的抗寒性(表5)。結果表明,57#、68#和32#為抗寒性較強的無性系,3#、25#和42#無性系抗寒性較弱。各無性系的綜合指數有一定的差異,其綜合指數為0.46-0.72,抗寒性最好的無性系為57#,其次為68#、32#、66#和74#,抗寒性最弱的無性系為42#,其余幾個無性系的綜合指數相對來說差距不大,為0.52-0.56。

表5 柳杉各無性系抗寒性綜合評價Table 5 Comprehensive evaluation of cold resistance in C. fortunei clones
3 討論
細胞膜是細胞感受環境脅迫最敏感的部位,當植物受到低溫脅迫時,細胞膜首先受到傷害,透性增大,使細胞內的電解質大量外滲。外滲液電導值的變化,可反映材料的細胞膜傷害程度,進而判斷植物抗寒性的大小[16-18]。Zhang等[12]在研究柳杉不同無性系低溫脅迫時相對電導率發現,電導率整體呈“S”型曲線,呈現先上升后基本不變的趨勢。10個柳杉無性系的半致死溫度在-4.7℃--8℃。其中57#和32#在整個過程中相對電導率相對于其他無性系較低,抗寒能力較強,42#相對電導率增幅最大,說明受低溫的影響較大,抗寒能力較弱。
此外,植物在逆境條件下往往發生膜脂過氧化作用,其產物MDA會嚴重損傷生物膜,通常利用它作為膜脂過氧化指標,表示細胞膜脂過氧化程度及對逆境反映的強弱[19]。本研究表明在低溫脅迫下,MDA含量隨著溫度的降低在一定溫度范圍內存在波動性,但總體來說呈現先上升后下降再上升的趨勢,表明在低溫脅迫下,柳杉細胞膜功能受到抑制,隨著溫度的降低MDA在細胞內積累,膜功能受阻或喪失,造成細胞代謝紊亂,從而對細胞產生傷害。柳杉各無性系MDA的含量沒有明顯的變化規律,其原因可能有兩個方面原因,一是MDA含量與某些植物的抗寒性并無太大關系,如王微[20]對紫花苜蓿幾個抗寒生理指標的研究中也出現MDA含量無變化規律的現象,她認為抗寒反應的生理機制比較復雜,單從某一個生理指標來鑒定抗寒性是比較困難的;二是試驗中采用離體枝條針葉進行測定,可能與整株植株生理指標變化有差異;三是整個低溫處理過程在黑暗的冰箱中進行,沒有考慮光照對生理指標的影響。
過氧化氫(H2O2)和超氧陰離子(O2-)是逆境情況下產生的活性氧(ROS)物質,低溫脅迫下植物會增加細胞內活性氧物質的含量[21]。超氧化物歧化酶(SOD)是生物防御系統的關鍵酶,在植物體內主要是用于清除超氧陰離子自由基等活性氧物質,維持植物體內活性氧代謝平衡,從而對機體起到保護作用[22-23]。因此,SOD酶活性的強弱能直接反映出植物在低溫脅迫下的自我調節能力,常被作為評價植物抗寒性的重要指標[7,24]。低溫脅迫下柳杉各無性系SOD酶活性整體上表現出先短暫的下降后逐漸上升而后又下降的趨勢。低溫初期SOD活性的降低與陳政等[19]對日本無刺花椒的研究結果一致。章錦濤等[25]在研究山茶的抗寒能力時發現隨著脅迫溫度的降低,植株體內SOD活性先升高后降低。在低溫脅迫下為了維持植物體內活性氧和自由基的動態平衡,植物體內的保護酶體系會發揮作用清除這些有害物質,使SOD等保護酶的活性增強,但超過保護酶清除作用范圍時其酶活性會下降[26-27]。郭祥泉[28]認為隨著低溫脅迫時溫度的降低,SOD酶活性有一個溫度拐點,在拐點前其活性急劇上升,在拐點后又下降,以此來定量研究不同樹種及不同個體的低溫臨界值。
光合作用是植物對逆境較為敏感的過程,低溫脅迫會抑制PS II的活性,使其光能捕捉能力與光化學轉化效率降低,反應中心的開放程度降低,從而引起葉綠素熒光參數的變化[29-30]。植物可變熒光Fv與最大熒光Fm的比值Fv/Fm能夠良好地反映光合色素把捕獲的光能轉化為化學能的速度和效率[31-32]。Fv/Fm下降越多表明PS II受損傷越大,Fv/Fm的變化可反映低溫下PS II受傷害的程度[33]。柳杉各無性系Fv/Fm值在4℃-0℃出現短暫的上升,0℃后隨著低溫脅迫的加劇而逐漸降低。Fv/Fm的降低說明PS II反應中心受到傷害,抑制了光合作用的原初反應,不利于光能的轉換。Y(II)值代表光系統 II的實際量子產量,它反映了植物的實際光合效率[29]。qP是衡量開放的光系統 II反應中心光抑制的程度,一定程度上表示PS II電子傳遞活性[34]。高qP值可以加速推動光合電子傳遞能力,促進光合效率增長[35]。Y(II)的變化趨勢與Fv/Fm的變化大致相同,隨著溫度的降低柳杉各無性系Y(II)值呈現先升高再降低的趨勢,而qP在低溫初期變化不明顯,0℃后與Y(II)值變化相同,隨著溫度的變化逐漸降低,甚至趨近于0,說明PS II反應中心的開放受到影響,實際光合效率和光合活性均受到低溫的抑制。Fv/Fm、Y(II)和qP值在4℃-0℃的回升及不變說明柳杉在經歷低溫初期有抵御能力,能夠在一定時間內維持光合效率,當超過臨界值其光合效率會大幅降低,嚴重時導致反應中心受到損傷甚至失活。
植物是一個統一的有機整體,逆境脅迫下引起植物生理生化指標變化的因素錯綜復雜,因此,單一指標很難準確評價植物的抗逆性[36-37]。主成分分析能有效地簡化數據,分析各指標間的關系,起到了濃縮數據的作用[14,38]。利用主成分分析及隸屬函數法,可以綜合評價柳杉各無性系的抗寒能力,在一定程度上可以真實的反映其抗寒能力強弱[39-40]。陳新華[41]運用綜合評價的方法,對不同品種甜櫻桃的抗寒性進行了評價,蘇李維等[14]用主成分分析法結合隸屬函數值建立了葡萄抗寒性綜合評價方法。本研究以相對電導率(REC)、MDA含量、SOD活性及葉綠素熒光參數(Fv/Fm)為評級指標,依據權重與隸屬函數計算柳杉各無性系抗寒性綜合指數,最終結果表明57#、68#和32#為抗寒性較強的無性系,3#、25#和42#無性系抗寒性較弱。
4 結論
低溫脅迫下,測定10個柳杉無性系針葉的丙二醛含量、超氧化物歧化酶活性及葉綠素熒光參數,其變化均具有顯著差異。結合主成分分析與隸屬函數法對10個柳杉無性系的抗寒性進行綜合評價,其中57#、68#和32#無性系對低溫脅迫具有較強的適應性,抗寒能力強,而3#、25#和42#無性系抗寒性較弱。
文章所有附圖附表數據請到本刊官網下載(http://biotech.aiijournal.com/CN/1002-5464/home.shtml)。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
