根際微生物對植物重金屬脅迫的緩解作用及其機理研究進展
2022-05-10 12:21:30楊露辛建攀田如男
生物技術通報 2022年3期
楊露 辛建攀 田如男
(南京林業大學風景園林學院,南京 210037)
由于農藥化肥的大量施用、污水污泥的不當處理、尾礦堆存和溶淋、大氣沉降及工業廢水的不合理排放等,重金屬污染已成為備受全球關注的生態環境問題之一[1]。重金屬廣泛分布于水體、土壤及空氣等介質,不但能夠經由根系吸收、葉面吸附等途徑進入植物體內,誘發細胞氧化應激反應,干擾多種生理代謝活動,包括水分與營養物質吸收、光合作用及內源植物激素穩態等,從而抑制植物生長與發育[2-7],而且可以通過食物鏈的富集作用對人類和其他生物健康構成嚴重威脅[8]。因此,重金屬污染環境的修復工作迫在眉睫。
在生態文明建設的國家重大需求下,重金屬污染環境的綠色修復是環境治理與修復領域的技術前沿和重點發展方向[9]。植物修復技術具有低成本、低維護、二次污染易控制及環境友好等優點,能夠通過植物根系吸收、揮發、過濾及穩定等作用有效降低環境中重金屬濃度,已成為重金屬污染環境綠色修復的重要手段。近年來,隨著重金屬污染環境植物修復研究的逐步深入,根際區域受到諸多學者的關注。研究表明,植物根系可以通過向根際環境釋放多種化學物質,改變土壤理化性質和微生物種類組成,調節其與微生物之間的關系,從而改善微生物的生存環境[10-11]。這種作用能夠反向促進寄主植物生長,并增強其對逆境脅迫的適應能力[12-13]。
目前,有關微生物緩解植物重金屬脅迫的研究陸續被報道,多集中于根際促生菌和菌根真菌等。鑒于此,本文就根際微生物組成、根系-微生物相互作用、根際微生物對植物重金屬抗逆性及其抵御重金屬脅迫的相關機制等方面進行綜述,以明確植物與微生物的聯結作用對重金屬脅迫下植物生長及其脅迫耐性的調控作用,同時為探索微生物輔助植物修復重金屬污染環境研究提供科學依據。
1 根際微生物
微生物是土壤生態系統中重要組成部分之一,在食物鏈中扮演著分解者的角色。它不但能夠通過硝化、氨化、反硝化及生物固氮等生化過程增加土壤中氮(N)、磷(P)濃度,從而提高土壤肥力[14-15],而且可以分解有機質消除污染物,分泌化學信號物質(如酚類、萜類化合物等)及抑制病原菌等,改善植物養分供應和環境適應能力[16-17]。其中,與植物相關的微生物被稱為植物微生物組,也被認為是植物的第二基因組。它包括附著在植物外表面的微生物(葉際)、寄生在組織內的微生物和與土壤根際密切相關的微生物群[18-19]。這些微生物群的許多成員對植物生長、土壤生態修復、水質修復及氣候變化等均有較大益處[20-22]。
根際區域是鄰接植物根系的土壤區域[23],含有高度多樣化的微生物群落,其基因數量遠超植物基因數量[24],被認為是最復雜的生態區域之一。根際區域是生物界的研究熱點之一,相關學者在根際發現了細菌、真菌、卵菌、線蟲、原生動物、藻類、病毒及古細菌等眾多微生物[25-26]。在根際區域中,一方面植物根系釋放分泌物吸引各類微生物構建根際微生物組;另一方面微生物通過各種活動調節植物與環境間的相互作用,促進植物生長,加速植物修復進程,從而改善生態環境[27]。某些微生物因其對植物和環境的有益作用而被廣泛關注,包括固氮細菌、菌根真菌、植物生長促進菌、生防微生物及真菌寄生菌等[24]。植物生長促進菌包括刺激植物生長或賦予植物脅迫抗性的有益真菌(plant growth-promoting fungi,PGPF)和細菌(plant growthpromoting bacteria,PGPB)[28]。其中,PGPF 主要與植物抗病能力有關[29-30],PGPB的部分成員(如植物根際促生菌,plant growth-promoting rhizobacteria,PGPR)多與植物重金屬脅迫耐性有關,在植物修復重金屬污染環境中具有重要作用[31-32]。
2 植物根系分泌物-根際微生物之間的相互作用
2.1 植物根系分泌物對根際微生物的影響
植物根系能夠釋放化學物質與根際微生物進行交流,并迅速建立有效共生關系,被認為是維持土壤環境穩態、促進植物生長的關鍵環節[33-34]。Akiyama等[35]發現,AM真菌在感應到宿主根系釋放的獨腳金內酯(SLs)后,菌絲會穿透并定殖于寄主根皮層細胞,并分化形成叢枝結構與植物進行營養交換。
植物根系可以分泌各種初級代謝產物(糖類、氨基酸等)和次級代謝產物(有機酸、萜類等),并通過傳遞、重塑等作用改變根際微生物區組。其中,某些分泌物可以調控微生物體內級聯反應,一方面促使微生物相關基因座的轉錄和特定化合物的生物合成,促進微生物的擴繁和定殖;一方面招募多種有益微生物以抵抗有害微生物,對植物根際微生物群落的建立和穩態有較大影響[36]。研究表明,銀杏(Ginkgo biloba)根系分泌的倍半萜內酯類化合物能夠誘導叢枝菌根真菌(arbuscular mycorrhizal fungi,AMF)孢子萌發和菌絲分枝形成,促進菌根定殖和宿主對水分、養分的吸收[35]。同時,宿主根系分泌水楊酸(SA)不但可以促進根際有益細菌的定殖,而且能夠通過驅動植物免疫信號在根際可用微生物群落中篩選并塑造優勢微生物群落[37]。此外,根系分泌物可能是驅動微生物群落交叉定殖的重要因素。研究發現,木豆(Cajanus cajan)-玉米(Zea mays)間作可以促進根系分泌類黃酮含量的增加,并改變有機酸(包括富馬酸、蘋果酸和琥珀酸等)種類與含量,從而誘導微生物對根系分泌物表現出顯著的趨化作用[38]。
此外,植物根系還能夠與某些真菌以互惠互利關系建立菌根共生體,通過釋放相關分泌物為真菌提供能量,而真菌則為宿主提供養分與水分,二者協調生長。龐大的菌根體系在地下網絡中連接同種或異種植物個體,介導植物養分和水分轉移、化學信號傳導等,從而增強宿主植物對逆境的耐受性[39]。
2.2 根際微生物對植物根系分泌物的反饋作用
微生物感知植物根系釋放的信號后會釋放相關物質,對宿主植物生長及其抗逆性等產生影響,由此形成土壤-根系-微生物的信號閉環。微生物可以分泌結瘤因子(nodulation factors,Nod factors)、菌根因子(mycorrhizal factors,Myc factors)、微生物相關分子模式(microbial-associated molecular patterns,MAMPs)和揮發性有機化合物(volatile organic compounds,VOCs)等物質,并通過改變植物激素網絡對宿主產生影響[40]。研究表明,植物體內水楊酸水平的增高會延遲AMF菌根定殖[41],而根瘤菌和菌根真菌釋放的Nod和Myc因子可以抑制宿主體內水楊酸介導的防御反應,并結合LysM類受體激酶啟動共生信號通路,促進微生物與宿主的共生定殖,從而促進植物生長[42-43]。微生物的MAMPs(包括脂多糖、冷休克蛋白、鞭毛蛋白和幾丁質等)可以促進植物系統獲得性抗性的形成,激發植物激素網絡(如水楊酸、茉莉酸、乙烯)級聯反應,從而提高植物應對生物或非生物脅迫的抗逆性[44-46]。
植物根系還能夠響應微生物釋放的VOCs(醇類、苯類、醛類及酸類等),其與光合相關信號共同作用可以激活擬南芥(Arabidopsis thaliana)根系中參與根際細菌介導的誘導系統抗性(rhizobacteriainduced systemic resistance,ISR)和鐵(Fe)吸收相關的MYB72,從而增強植物根系對微量元素Fe的吸收能力[47]。假單胞菌(Pseudomonas simiae)產生的揮發性VOCs能夠誘導編碼營養貯藏蛋白(vegetative storage protein,VSP)、γ-谷氨酰水解酶(gamma-glutamyl hydrolase,GGH)和RuBisCo大鏈蛋白基因的上調表達來促進大豆(Glycine max)生長并緩解逆境傷害[48]。微生物與植物產生并傳遞的相關化合物影響著植物基因表達、根形態構成和植物系統抗性等,間接證明了微生物向植物釋放環境脅迫信號的現象[49]。
3 根際微生物對植物重金屬脅迫的緩解作用
3.1 植物根際促生菌
接種植物根際促生菌(PGPR)可以明顯減輕重金屬對植物的毒害作用。Saleem等[50]發現,接種熱氏假單胞菌(Pseudomonas gessardii)或熒光假單胞菌(Pseudomonas fluorescens)能夠顯著促進向日葵(Helianthus annuus)葉片光合作用,并增加株高、根長及干重,從而減弱鉛(Pb)脅迫毒害程度,相似的結果也被發現于重金屬脅迫下的紫色苜蓿(Medicago sativa)[51]、小白菜(Brassica campestris ssp. chinensis)[52]及水稻(Oryza sativa)[53]等多種植物。
首先,PGPR可以誘導寄主植物根系表面形成生物膜并定殖,其能夠有效緩解植物遭受的氧化損傷。研究發現,接種耐重金屬菌團(如Proteus sp.、Pseudomonas sp.及 Ensifer meliloti) 能 夠 促 進紫花苜蓿合成非酶類抗氧化劑(抗壞血酸、谷胱甘肽和多酚)和酶類抗氧化劑(CAT、SOD、GPX和APX),有效清除細胞中的ROS,降低氧化應激指數(oxidative stress index,OSI),從而緩解重金屬誘導的氧化損傷[54]。同時,PGPR可以降低植物呼吸碳消耗,并分泌生長素、赤霉素及細胞分裂素等植物激素,從而促進宿主根系生物量增加以固定更多的重金屬[55-56]。Kotoky等[57]發現,黏質沙雷氏菌(Serratia marcescens S2I7)能夠分泌大量生長素(IAA),促進水稻根長、莖長及干重的增加,從而促進植株生長并增強其對Cd脅迫的抗性。其次,PGPR能夠溶解礦質養分為植物生長發育提供能量。研究發現,根際溶磷菌可以通過溶磷機制溶解無機磷酸鹽為宿主植物提供磷元素,從而促進植株分蘗數、根系體積及干重的增加[58]。此外,在重金屬脅迫下,PGPR可以與植物根系結合產生胞外聚合物以絡合重金屬,或通過誘導脅迫相關基因和蛋白表達,增強植物生理代謝作用,從而促進植物生長,并增強其抗逆性[59-60]。Kong等[61]發現,接種Sinorhizobium meliloti可以誘導天藍苜蓿(Medicago lupulina)CuZnSODc、CuZnSODp和APXc等抗氧化基因的上調表達,促進抗氧化酶活性升高,從而提高植物應對過量銅脅迫的抗氧化防御能力,相似的結果也被發現于鎘脅迫下的番茄[62]。
3.2 菌根真菌
菌根包括外生菌根、叢枝菌根和內生菌根[63],其中外生菌根、叢枝菌根與植物重金屬脅迫耐性密切相關。菌根結合和菌根類型可以促進植物對營養物質的吸收和病原體拮抗物的產生,從而改變生物多樣性,同時菌根結合可以有效降低土壤中重金屬離子的遷移和生物可利用性,有利于改善植物抗逆性[39,64]。
在重金屬脅迫下,外生菌根可以誘導氮代謝、乙醇及脂肪酸代謝等相關基因的表達,從而有效緩解重金屬毒害作用。Shi等[65]等發現,外生菌根一方面通過硝酸還原酶家族基因(nidD,niiA)調節反硝化和還原六價鉻(Cr6+)以緩解其誘發的氧化脅迫,一方面可以通過上調乙醇脫氫酶Ⅲ啟動子(alcC)、乙醛脫氫酶(aldA)等基因,保護巰基、促進次生代謝產物及長鏈不飽和脂肪酸生物合成,相似的結果也被發現于重金屬脅迫下的Saccharomyces cerevisiae[66]。Khullar等[67]發現,外生菌根可以合成金屬硫蛋白、谷胱甘肽等非蛋白巰基肽化合物,其可以通過螯合作用降低胞內重金屬離子濃度。同時,外生菌根可以有效保證宿主植物對養分與水分的吸收、運輸和利用,從而有利于植株生長及其抗逆性[68]。研究表明,接種外生菌根(Pisolithus tinctorius)可以明顯降低巨桉(Eucalyptus grandis)降低葉片錳含量,促進葉片碳同化、水分利用效率及養分吸收速率等[69]。Szuba等[70]發現,耐鉛真菌(Paxillus involutus)可以改善Pb2+脅迫下楊樹(Populus canescens)葉片光合作用,并能夠將Pb2+固定在真菌細胞中,從而緩解Pb2+植物毒性。
接種叢枝菌根不但能夠改變介質中重金屬形態,而且可以促進宿主植物水分養分的吸收、光合過程中碳水化合物的交換等,從而促進植物生長及其對重金屬脅迫的耐受性。研究表明,接種AMF可以提高重金屬脅迫下玉米葉片葉綠素含量、光合速率、氣孔導度和蒸騰速率,提升植株產量和品質,并有效降低土壤有效態Cd2+、Pb2+含量[71],這可能是因為過量重金屬可以誘導AMF菌絲合成和積累不溶性糖蛋白(球囊素),其能夠通過螯合作用改變根際重金屬化學形態,從而降低其生物可利用性[72]。同時,AMF還可以促進菌根內特定基因上調,誘導金屬硫蛋白、分泌蛋白及轉運蛋白生物合成,從而增強宿主對重金屬脅迫的抗性[73]。Meier等[74]發現,GrosMT1、鋅轉運體GinZnT1、GmarMT1 和GintABC1參與維持AMF菌體胞內重金屬穩態過程。過量鋅(Zn)能夠促進GinZnT1的表達并將Zn2+區隔于液泡中,同時上調GmarMT1以促進金屬硫蛋白的生物合成,并調節細胞氧化還原電位,保護植物免受重金屬誘發的氧化損傷等。此外,AMF能夠促進植物體內特異性磷酸鹽轉運蛋白家族基因的表達,促進細胞中P濃度的增加,從而提高葉片光合作用、植株生長量、單株鈴數和纖維成熟度[75](表1)。
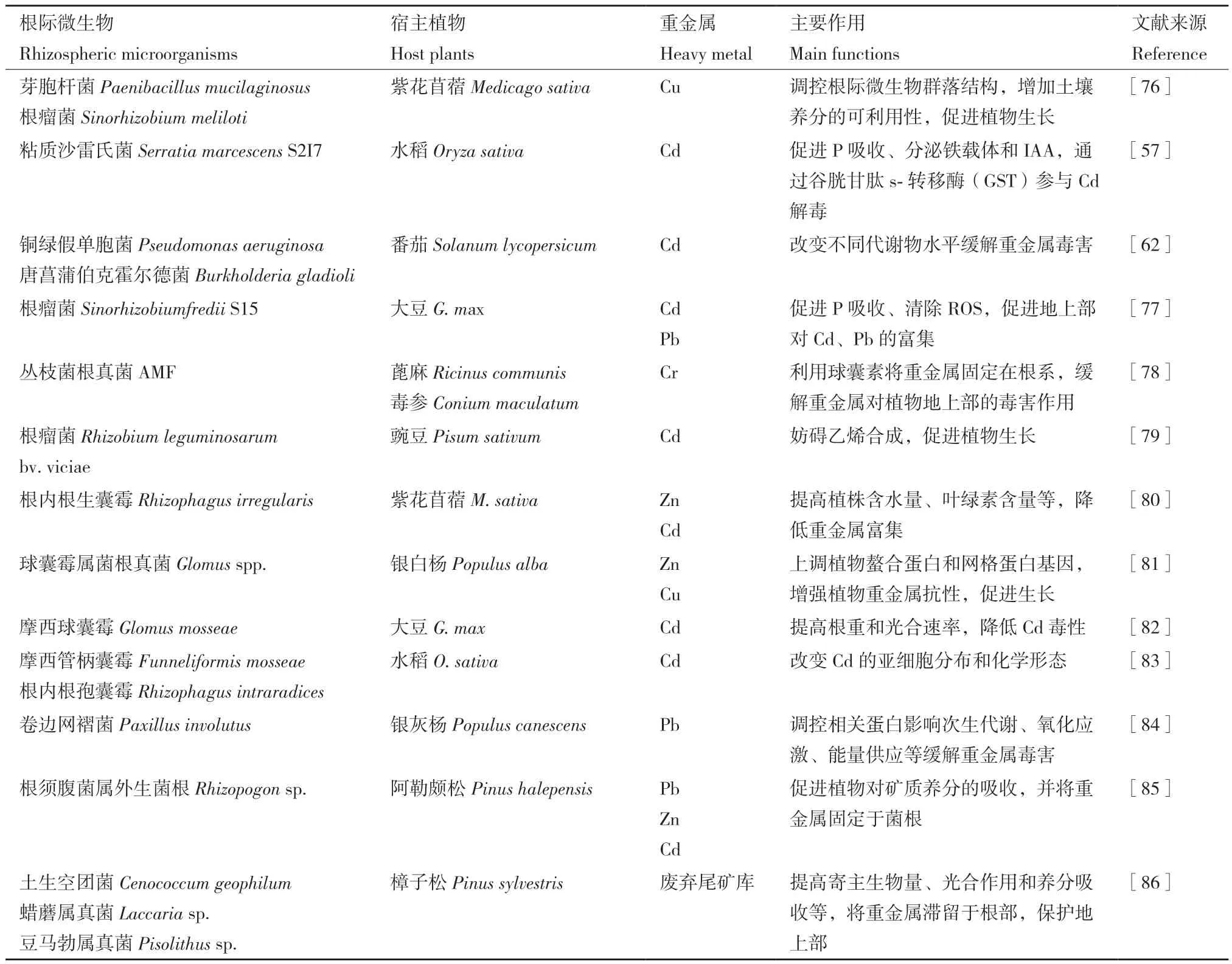
表1 根際微生物對重金屬脅迫下植物生長與生理代謝促進作用Table 1 Effects of rhizosphere microorganisms on plant growth and physiological metabolism under heavy metal stress
4 根際微生物緩解植物重金屬脅迫作用機制
4.1 微生物自身防御機制
在重金屬脅迫條件下,微生物可以通過胞外吸附與沉淀、胞內區隔以緩解重金屬生物毒性。過量重金屬脅迫不但可以誘導細菌形成胞外多糖,為金屬陽離子提供吸附位點[87],而且可以促使抗性基因(如copA、copB、copC和copD)編碼蛋白形成重金屬離子的胞外結合位點,避免其進入胞內破壞組織結構[88]。同時,微生物可以促使類金屬硫蛋白基因(smtA)編碼金屬硫蛋白,與重金屬結合進行胞內區隔[89],并通過分泌大量的谷胱甘肽、草酸等化合物形成胞外沉淀[90-91]。其次,Misra等[92]發現,細菌可以進化出一組具有汞(Hg)抗性的操縱子基因(mer)以適應Hg(II)脅迫,并誘導有機汞裂解酶和汞離子還原酶的生物合成,從而降低Hg毒性。此外,菌絲體是菌根植物的一道天然屏障,其可以通過吸附作用將重金屬固定在菌絲、叢枝和小泡結構中,增強植物根系固定重金屬的能力,從而抑制重金屬從根系到地上部的遷移能力[93-94]。
4.2 促進植物根系分泌物釋放
逆境會刺激根系分泌物在根際區域釋放,其能夠與重金屬離子結合形成穩定復合物,從而影響重金屬在介質中的遷移率、溶解性和生物有效性等[95]。其中,鐵載體、有機酸及生物表面活性劑能與重金屬離子發生絡合或螯合反應,增加土壤中重金屬離子的有效性和溶解性,從而提高植物對有毒離子的吸收能力[96]。Shi等[97]發現,接種耐Cd銅綠假單胞菌(Pseudomonas aeruginosa)可誘導龍葵(Solanum nigrum)根系分泌鐵載體,其能夠螯合Cu2+、Cd2+、Zn2+及Ni+,促進幼苗對Cd2+的吸收,并改善植株長勢。同時,小米(Setaria italica)根系分泌的低分子量有機酸(LMWOA)能夠與Cd2+螯合形成Cd-LMWOA,促進根系對Cd2+的吸收[98]。此外,根際微生物可以合成胞外聚合物、糖蛋白,其能夠通過固定作用降低重金屬在土壤中的遷移能力[72]。
4.3 抑制植物乙烯合成
植物受到重金屬脅迫時會產生大量乙烯,誘導葉片早衰脫落、根系生長抑制等,從而妨礙植物生長發育,并降低其抗逆性[99]。植物根際微生物能通過酶解乙烯合成的直接前體1-氨基環丙烷-1-羧酸(ACC)來減少乙烯生物合成,從而緩解乙烯對植株產生的傷害[100]。目前,ACC脫氨酶活性是植物根際有益微生物提高宿主脅迫耐性的研究熱點之一[101]。研究表明,在Cd脅迫下,接種含ACC脫氨酶的根瘤菌不但能夠顯著提高豌豆(Pisum sativum)結瘤儲量、地上部生物量、共生固氮能力,而且能夠改善植株水分利用效率和促進礦質營養吸收,還能夠增強植株對Cd2+的吸收能力[79]。接種含ACC脫氨酶的邊緣假單胞菌(Pseudomonas marginalis)和紅球菌屬菌株(Rhodococcus sp.)能刺激宿主植物根系生長和養分吸收,由此改善Cd對植物生長的抑制效應[102]。
4.4 促進植物光合作用
過量重金屬會破壞植物葉片葉綠素和相關酶的生物合成,進而抑制光合作用。接種具有重金屬耐性的促生細菌(包括根瘤菌、熒光假單胞菌和假單胞菌等)能夠顯著增加植株葉片光合速率、蒸騰速率及水分利用效率等氣體交換參數,從而改善宿主光合性能,并促進其生長[103]。同時,接種菌根真菌能通過誘導植物光合相關蛋白的生物合成與表達,促進植株光合作用并改善其生長。研究發現,接種AMF能夠誘導植物葉綠素合成蛋白表達,提高葉綠素中必需元素(如Mg、Fe)的攝取,從而促進葉綠素生物合成[104-105];接種卷邊網褶菌(Paxillus involutus)能夠顯著提高白楊(Populus canescens)體內與蛋白質轉換、脅迫反應、碳水化合物代謝等相關的葉片蛋白豐度,促進植株光合作用與生長長勢[106]。此外,菌根真菌還能夠提高宿主植物氣孔導度和光合相關基因的轉錄水平,從而促進植物光合氣體交換能力和光合效率[107]。Wu等[108]發現,接種AMF能夠誘導蘆葦(Phragmites australis)體內與光合電子傳遞相關的petH基因編碼,上調鐵氧還原蛋白-NADP+氧化還原酶表達,從而促進光合電子傳遞,增強植株光合活性。
4.5 激活植物抗氧化防御系統
重金屬脅迫會誘導植物細胞活性氧(ROS)過量積累,從而擾亂植物體內活性氧產生和清除之間的動態平衡。因此,提高抗氧化防御系統活性是維持重金屬脅迫下植物體內氧化還原狀態平衡的關鍵環節之一[109]。接種根際有益微生物可以明顯提高植物體內超氧化物氣化酶(SOD)、過氧化氫酶(CAT)、過氧化物酶(POD)、乙二醛酶 I(GlyI)、乙二醛酶II(GlyII)活性及谷胱甘肽含量,從而緩解ROS誘發的氧化損傷[110-111]。研究發現,植物可以上調GlyI、GlyII活性并協同GSH實現對胞內甲基乙二醛的解毒作用[112]。Zhang等[111]發現,接種AM菌能夠通過下調蒺藜苜蓿(Medicago truncatula)根內MtRbohC-G的轉錄水平,降低NADPH氧化酶介導的H2O2積累,從而清除多余的ROS來維持細胞氧化還原穩態。同時,接種PGPR能夠調控宿主體內不同抗氧化酶、應激蛋白和乙烯編碼蛋白的基因表達以提高自由基清除劑水平,從而緩解脅迫對植物產生的傷害。Khanna等[113]發現,在Cd脅迫下,接種銅綠假單胞菌可以上調番茄SOD、POD和PPO基因表達量,并下調CAT、GR、GST、GPOX和APOX基因表達量,通過調節抗氧化防御蛋白表達來降低Cd誘導的番茄氧化應激。
4.6 促進植物水分與養分的吸收
菌根定殖后可形成發達的菌套和菌絲體,其可以顯著增加根系表面積,促進根系對水分的吸收,從而緩解由非生物脅迫引起的生理性干旱。研究發現,接種外生菌根(Hebeloma crustuliniforme)能夠有效促進顫楊(Populus tremuloides)幼苗胞間水分運輸,使根系導水率顯著高于未接種植株[114]。同時,有益微生物可以釋放生長素(IAA)、細胞分裂素(CTK)及赤霉素(GA)等植物激素以溶解礦質元素,并改善菌根定殖率和密度,從而提高宿主植物地上部對N、P及Fe等礦質元素的吸收和利用[115]。此外,某些微生物體內含有新輔酶吡咯喹啉醌基因(pqqABCDE),其參與葡萄糖脫氫酶(Gdh)輔助因子——吡咯并喹啉醌的生物合成過程,而Gdh蛋白能產生有機酸(葡萄糖酸)溶解無機磷酸鹽,為植物提供養分。研究表明,接種含該基因的促生菌能夠顯著提高宿主植物體內P含量,促進植株根系生長和生物量增加,從而改善其生長[116-117]。
4.7 改善根際土壤微環境
根際微生物是根際土壤物質循環及轉換的主要驅動者。微生物在爭奪生態位及定殖過程中,通過轉化難溶有機質、固氮、溶磷等方式,促進土壤中N、P、C的循環,從而改善農田土壤的養分環境[118]。根際有益微生物區組能夠極大程度地改良土壤質量,包括土壤團粒結構、含水量、pH、土壤酶活性及微生物豐富度等。研究發現,微生物可以通過分泌胞外聚合物(EPS),改善土壤物理結構并促進土壤水分保持及傳導,甚至有些微生物可以將自身嵌入其中,從而在惡劣環境中生存[119-120]。劉暢等[121]發現,接種PGPR復合菌劑(假單胞菌屬Pseudomonas hunanensis,芽孢桿菌屬Bacillus flexus,束村氏屬Tsukamurella sp.,伯克霍爾德氏菌屬Burkholderia sp.)能夠有效增加根際土壤氮循環功能菌群、溶磷菌及解鉀菌的數量,并顯著提高土壤脲酶、過氧化氫酶和中性磷酸酶活性,促進農田土壤環境良性循環,從而緩解重金屬對植物的毒害作用。同時,根際微生物還可以通過改變土壤中重金屬化學形態,降低重金屬離子可交換性及生物利用性,從而間接地促進作物生長。任曉斌等[122]發現,球形紅細菌(Rhodobacter sphaeroides)和生物炭的聯合使用能夠降低土壤中可交換態Cr含量,增加殘渣態和有機結合態Cr含量,從而促進Cr污染土壤的植物修復。
5 總結與展望
土壤-植物-微生物是一個緊密聯合的生物網絡,微生物群落、植物根系和土壤環境之間發生的各種生理生化反應可以促進土壤生態系統中能量流動、物質循環和信號傳遞的順利進行,從而使其在植物修復中具備巨大潛力[27]。在逆境脅迫下,植物根系-微生物通過釋放相關信號相互作用,促使根際微生物組重塑,從而改善土壤生態環境,促進宿主植物生長并增強其脅迫耐性,為土壤-植物-微生物網絡構建良好的生態平衡。
根際有益微生物促進植物根系分泌物釋放、水分與養分吸收、葉片光合作用,清除細胞ROS、抑制乙烯生物合成和改善根際土壤微環境,從而緩解重金屬植物脅迫。今后可針對根際微生物緩解植物重金屬脅迫的研究繼續開展以下研究:
(1)目前,有關微生物增強植物重金屬脅迫耐性的研究多集中于芽孢桿菌屬(Bacillus)、鏈霉菌屬假單胞菌類(Streptomyces pseudomonas)、寡養單胞菌屬(Stenotrophomonas)、沙雷氏菌屬(Serratia)、球囊霉屬(Glomus)及根生囊霉屬(Rhizophagus)等菌株[123-124]。微生物組作為植物的第二基因組,其復雜程度不亞于植物體基因,故今后可以從不同重金屬污染地區分離、篩選和培養新菌株,進一步挖掘在逆境脅迫下對植物生長有益的新菌種。
(2)有關微生物緩解植物重金屬脅迫的多數研究為控制實驗,注重某一類微生物對單一重金屬脅迫下植物生長及其脅迫耐性的改善[57,65,67,70],這與野外自然環境之間存在較大差別,忽略了不同微生物之間的相互作用。因此,今后可以通過礦區廢棄地復綠、濕地重金屬污染污染物修復等實踐考察根際微生物間相互作用及其對宿主植物逆境耐性的影響,并篩選出較優的植物-微生物組合。
(3)根際微生物是重金屬與植物接觸的第一道防線,在增強植物重金屬脅迫耐性中發揮了重要作用。Smith[125]認為,與植物育種和遺傳改良方法相比,基于微生物的植物生物技術在改善植物非生物脅迫耐性中具有成本低、時間短和效率高等特點。以往研究多關注某一類微生物改善植物重金屬脅迫耐性機制,忽略了重金屬脅迫下根際微生物與植物如何協同增強植物逆境耐性的相關機制。因此,有必要明確重金屬脅迫下植物根系-微生物互作體系的生理分子機制,從而為該互作體系在重金屬污染環境中的實際應用提供科學依據。
猜你喜歡
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
