458份小麥品種(系)抗病性狀功能基因的KASP標(biāo)記檢測(cè)
2022-05-12 13:06:20張宏芝王立紅李劍峰樊哲儒張躍強(qiáng)
新疆農(nóng)業(yè)科學(xué) 2022年4期
關(guān)鍵詞:檢測(cè)
王 重,張宏芝,高 新,時(shí) 佳,王立紅,李劍峰,樊哲儒,趙 奇,張躍強(qiáng)
(1.新疆農(nóng)業(yè)科學(xué)院核技術(shù)生物技術(shù)研究所,烏魯木齊 830091;2.農(nóng)業(yè)部荒漠綠洲作物生理生態(tài)與耕作重點(diǎn)實(shí)驗(yàn)室,烏魯木齊 830091;3.新疆農(nóng)作物生物技術(shù)重點(diǎn)實(shí)驗(yàn)室,烏魯木齊 830091)
0 引 言
【研究意義】新疆地處亞洲大陸中心,降雨量少,蒸騰量大,氣候干燥,小麥病害發(fā)生較少[1],近年來(lái)由于氣候變化、栽培方式改變、致病菌種變異以及抗病性喪失等因素,小麥種植區(qū)白粉病和銹病發(fā)生頻率和病害有所加重[2]。目前防治小麥白粉病和銹病病害的主要措施是農(nóng)藥,將常規(guī)育種手段與分子輔助選擇育種技術(shù)相結(jié)合,選育抗病分子標(biāo)記聚合的育種材料或品種,對(duì)于新疆的小麥生產(chǎn)有實(shí)際意義。【前人研究進(jìn)展】競(jìng)爭(zhēng)性等位基因特異性PCR(kompetitive allele specific PCR,KASP)是一種對(duì)特定位點(diǎn)處SNP以及插入和缺失(insertion-deletion,InDels)進(jìn)行雙等位基因分型技術(shù),可針對(duì)絕大多數(shù)基因組DNA的SNPs和特定位點(diǎn)InDels,準(zhǔn)確分析雙等位基因,具有穩(wěn)定性好、準(zhǔn)確性高、無(wú)需合成特異熒光探針、檢測(cè)成本較低和高通量等優(yōu)點(diǎn)[3]。李瑋等[4]對(duì)KASP對(duì)94份小麥育種親本的功能基因進(jìn)行分子檢測(cè)表明,與抗性相關(guān)的Lr68優(yōu)異等位變異占比為15%,未檢測(cè)到Lr34和Pm21陽(yáng)性材料。利用KASP標(biāo)記技術(shù)對(duì)120份小麥品種(系)的重要性狀功能基因進(jìn)行檢測(cè),69份材料含有Lr14a,3份材料含Lr68基因,未發(fā)現(xiàn)含有Yr15基因材料[5]。高潔等[6]對(duì)小麥4個(gè)多效抗病基因分子標(biāo)記進(jìn)行了轉(zhuǎn)化和再開(kāi)發(fā),并對(duì)其進(jìn)行了驗(yàn)證。【本研究切入點(diǎn)】目前通過(guò)KASP檢測(cè)的材料多以當(dāng)?shù)刂髟云贩N、育種骨干親本及衍生品系為主,針對(duì)新疆小麥品種(系)及育種親本的抗白粉病、銹病基因分子檢測(cè)少見(jiàn)報(bào)道。需采用KASP標(biāo)記技術(shù)檢測(cè)抗白粉病基因Pm21、抗條銹病基因Yr15和抗葉銹病基因Lr14a、Lr68。【擬解決的關(guān)鍵問(wèn)題】以56份新疆小麥品種、304份小麥品系和98份引進(jìn)種質(zhì)資源為試驗(yàn)材料,研究不同小麥品種(系)抗病性狀功能基因的分布狀態(tài),分析小麥品種(系)的抗病功能基因,為小麥抗病育種提供理論依據(jù)。
1 材料與方法
1.1 材 料
1.1.1 小麥品種(系)
參試小麥品種(系)總計(jì)458份材料,其中新疆審定小麥品種56份,高代穩(wěn)定品系304份,引進(jìn)種質(zhì)資源98份。
1.1.2 KASP標(biāo)記
抗白粉病標(biāo)記1個(gè),Pm21;條銹病抗性標(biāo)記1個(gè),Yr15;葉銹病抗性標(biāo)記2個(gè),Lr14a和Lr68。引物參照中國(guó)農(nóng)科院作物科學(xué)研究所夏先春老師提供序列由上海生工合成。
1.2 方 法
1.2.1 小麥DNA提取
參試小麥品種(系)播種于育苗盤中,待其生長(zhǎng)至3葉1心取葉片置于1.5 mL離心管中,鋼珠震蕩破碎,采用PVP40(聚乙烯吡咯烷酮40)法[7]提取小麥基因組DNA。DNA提取完成后使用Nanodrop one對(duì)DNA濃度測(cè)定,并定容至100 ng/μL。
1.2.2 抗性標(biāo)記檢測(cè)
以參試小麥品種(系)DNA為模板,選擇中國(guó)春等5個(gè)已知分子標(biāo)記品種作為陽(yáng)性對(duì)照,采用擴(kuò)增子拯救多重PCR(amplicon rescued multiplex PCR,arm-PCR)技術(shù)對(duì)小麥基因組DNA的抗性標(biāo)記檢測(cè)[8]。
1.2.2.1 PCR體系
PCR反應(yīng)體系10 μL,包括DNA模板2 μL,2×KASP Mix 5 μL,引物混合液0.1 μL,ddH2O 2.9 μL。引物混合液由合成的FAM引物(100 μM)、HEX引物(100 μM)、通用引物(100 μM)和ddH2O按照12∶12∶30∶46的比例進(jìn)行混合制成。
1.2.2.2 PCR程序
預(yù)變性94℃ 5 min;變性94℃ 20 sec,退火65~56℃1 min,每個(gè)循環(huán)降低1℃,10個(gè)循環(huán);變性94℃ 20 sec,退火56℃ 1 min,30個(gè)循環(huán)。
2 結(jié)果與分析
2.1 抗白粉病基因Pm21檢測(cè)
研究表明,僅有2份材料檢測(cè)到Pm21標(biāo)記,約占群體的0.44%,且這2份材料均為審定品種。抗白粉病分子標(biāo)記Pm21在審定品種中有極少量分布,但是在目前的高代穩(wěn)定品系中,以及引進(jìn)種質(zhì)資源中,均未發(fā)現(xiàn)白粉病抗性標(biāo)記Pm21。圖1
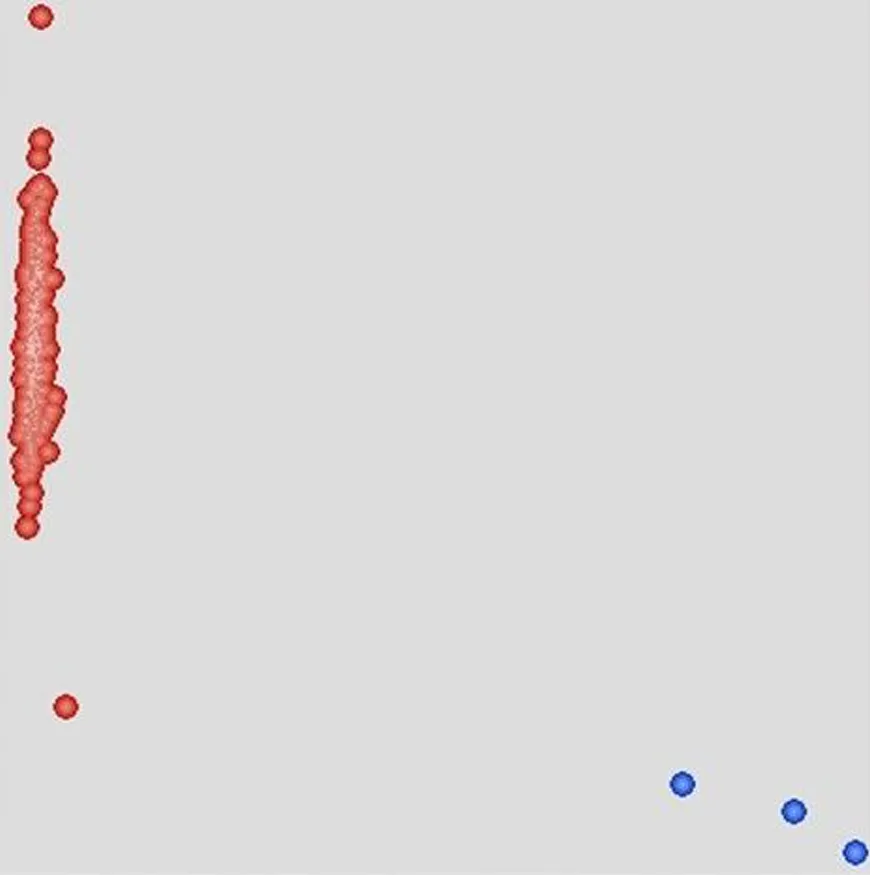
注:紅色:無(wú)Pm21;藍(lán)色:攜帶Pm21
2.2 抗條銹病基因Yr15檢測(cè)
研究表明,14份材料檢測(cè)到Y(jié)r15標(biāo)記,約占群體的3.06%,其中,審定品種7份,高代穩(wěn)定材料6份,引進(jìn)種質(zhì)資源1份。雖然條銹病抗性標(biāo)記Yr15在參試品種(系)中的分布頻率較低,但是無(wú)論在審定品種或者高代品系中均存在較為穩(wěn)定的分布。圖2
2.3 葉銹病抗性基因檢測(cè)
2.3.1 抗性標(biāo)記Lr14a檢測(cè)
研究表明,22份材料檢測(cè)到Lr14a標(biāo)記,約占參試材料的4.80%。其中,審定品種21份,引進(jìn)種質(zhì)資源1份。Lr14a較為集中的分布于冬小麥審定品種中,共有20份材料,其余2份材料分布于春小麥品種和引進(jìn)種質(zhì)資源中。圖3
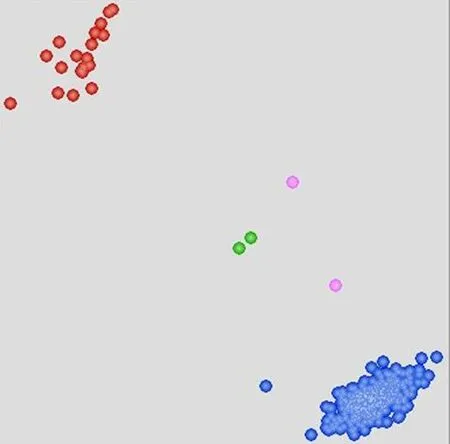
注:紅色:攜帶Yr15;藍(lán)色:無(wú)Yr15

注:紅色:攜帶Lr14a;藍(lán)色:無(wú)Lr14a
2.3.2 抗性標(biāo)記Lr68檢測(cè)
研究表明,有48份材料檢測(cè)到Lr68標(biāo)記,約占供試材料的10.48%。其中,審定品種4份,高代品系15份,引進(jìn)種質(zhì)資源29份。雖然新疆小麥審定品種中Lr68分布較為少見(jiàn),但是在目前的高代品系和引進(jìn)種質(zhì)資源中Lr68的分布頻率顯著增加。圖4
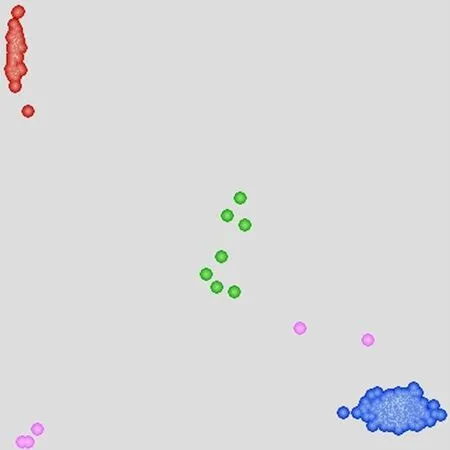
注:紅色:攜帶Lr68;藍(lán)色:無(wú)Lr68
2.4 小麥抗病功能基因分布
研究表明,抗葉銹病功能基因Lr68的分布頻率最高,達(dá)到了10.48%,其次是葉銹病抗性基因Lr14a,分布頻率為4.80%,抗條銹病基因Yr15分布頻率為3.06%,抗白粉病功能基因Pm21分布頻率最低,僅為0.44%。在不同小麥類型中,冬小麥抗葉銹病功能基因Lr14a的分布頻率較高,抗白粉病功能基因Pm21僅有的2份材料均為冬小麥,而春小麥中,抗條銹病功能基因Yr15和抗葉銹病基因Lr68的分布頻率較高且多于冬小麥。根據(jù)參試小麥品種(系)不同來(lái)源分析,在審定品種中4個(gè)抗病基因均有分布,引進(jìn)種質(zhì)資源中未見(jiàn)白粉病抗性基因Pm21,條銹病抗性基因Yr15及葉銹病抗性基因Lr14a僅發(fā)現(xiàn)1份材料,而高代品系中Pm21和Lr14a均未檢測(cè)到。表1
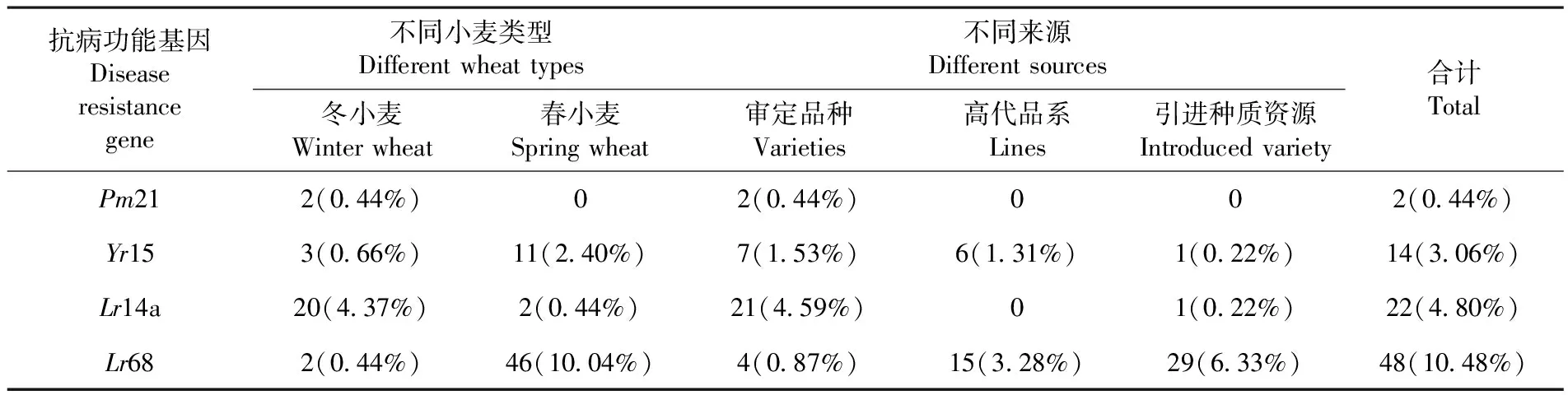
表1 小麥抗病功能基因分布狀態(tài)Table 1 Distribution of disease-resistance genes in wheat
3 討 論
小麥抗白粉病基因Pm21位于小麥6A染色體上,來(lái)源于簇毛麥[9-10]。由于小麥白粉病菌變異性較強(qiáng),部分小麥抗白粉病基因抗性逐漸減弱甚至喪失,而Pm21在近幾年的鑒定中仍然保持了很好的抗性,且具有較廣的抗譜,可以作為小麥抗白粉病育種的重要抗性來(lái)源[11]。但Pm21在國(guó)內(nèi)小麥品種中分布較少,李瑋等[4]對(duì)94份小麥品種或高代品系進(jìn)行檢測(cè),未發(fā)現(xiàn)Pm21陽(yáng)性材料,王鑫等[12]對(duì)305份國(guó)內(nèi)外小麥種質(zhì)進(jìn)行鑒定,僅發(fā)現(xiàn)1份材料含有Pm21。研究中在458份小麥品種(系)中僅檢測(cè)到2份材料攜帶Pm21,在新疆小麥品種(系)中,Pm21的分布極少,與前人研究結(jié)果基本一致。
小麥抗條銹病功能基因Yr15位于小麥1B染色體上,來(lái)源于野生二粒小麥[13]。目前Yr15是少數(shù)幾個(gè)對(duì)于中國(guó)條銹菌流行小種仍然保持較好抗性的基因[14],對(duì)于抗條銹病小麥育種而言,Yr15是極其重要的抗性來(lái)源。鄒景偉等[5]對(duì)120份小麥品種(系)進(jìn)行了檢測(cè),未發(fā)現(xiàn)含有Yr15材料,王鑫等[12]在305份國(guó)內(nèi)外小麥種質(zhì)中發(fā)現(xiàn)10份攜帶Yr15材料。研究在458份小麥品種(系)中檢測(cè)到含有Yr15材料14份,這與王鑫等[12]的結(jié)果基本一致,Yr15分布頻率均為3%左右。在高代品系中有6份材料含有Yr15,Yr15通過(guò)雜交等手段成功導(dǎo)入后代材料中,進(jìn)一步工作中可以加強(qiáng)分子標(biāo)記輔助選擇,篩選攜帶Yr15后代材料,提升新品種條銹病抗性。
小麥抗葉銹病功能基因Lr14a位于小麥7B染色體上,來(lái)源于野生二粒小麥[15-16];Lr68也位于小麥7B染色體上,來(lái)源于野生二粒小麥,屬于慢銹病基因,即反應(yīng)型表現(xiàn)出感病,但嚴(yán)重度較低[17]。Lr14a和Lr68在小麥中的分布較為廣泛,李瑋等[4]對(duì)94份小麥品種或高代品系進(jìn)行鑒定,發(fā)現(xiàn)Lr14a陽(yáng)性率約為50%,Lr68為15%,鄒景偉等[5]在120份小麥種質(zhì)中發(fā)現(xiàn)69份材料含有Lr14a,3份材料含有Lr68。研究在458份小麥材料中檢測(cè)到含有Lr14a材料22份,Lr68材料48份,這與前人結(jié)果并不一致。可能是由于春小麥材料中Lr14a的分布頻率太少,導(dǎo)致整體分布頻率偏低,而Lr68在引進(jìn)種質(zhì)資源中分布頻率較高,并成功導(dǎo)入高代品系中,在高代品系中分布頻率也提高。
4 結(jié) 論
小麥抗病基因Pm21、Yr15和Lr14a的分布頻率仍然較低,均低于5%,而Lr68的分布頻率雖然超過(guò)10%,但是在審定品種中分布頻率也并不高。
新疆小麥品種(系)中抗病基因的分布頻率較低,約為5%。從458份小麥品種(系)中檢測(cè)到含有Pm21材料2份,含有Yr15材料14份,含有Lr14a材料22份,含有Lr68材料48份。
猜你喜歡
中國(guó)設(shè)備工程(2022年12期)2022-07-11 04:33:00
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年9期)2019-11-25 07:34:36
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年9期)2019-11-25 07:34:34
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年12期)2019-05-21 02:53:50
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年12期)2019-05-21 02:53:48
