嬰兒配方乳粉脂質母乳化的研究進展
2022-05-13 02:32:22李曉東周星宇陸欣宇唐雅倩李春梅冷友斌蔣士龍
食品科學 2022年7期
李曉東,林 爽,劉 璐,周星宇,陸欣宇,唐雅倩,李春梅,冷友斌,蔣士龍
(1.東北農業大學食品學院,乳品科學教育部重點實驗室,黑龍江 哈爾濱 150030;2.黑龍江省綠色食品科學研究院,黑龍江 哈爾濱 150028;3.黑龍江飛鶴乳業有限公司,黑龍江 哈爾濱 150030)
母乳作為滿足嬰兒生長發育的最佳能量來源,可以在嬰兒出生后前6個月為其提供所需的全部營養,但由于母親身體狀況、環境因素等限制,很大一部分嬰兒無法實現純母乳喂養,嬰兒配方乳粉應運而生。乳脂作為乳中的重要成分,約占母乳的3%~5%,為0~6個月嬰兒幾乎提供50%生長所需能量,此外還是許多必需營養物質的重要來源,如必需脂肪酸、脂溶性維生素和一些生物活性物質等[1]。研究顯示母乳和嬰兒配方乳粉中脂質組成和結構的差異會影響脂質的消化吸收,甚至可能通過影響脂質代謝來引起嬰兒生命早期“代謝程序化”的改變,進而對個體嬰兒期,甚至成年之后的健康發育產生深遠影響[2],目前對于脂質結構與“代謝程序化”的關系研究主要體現在嬰兒成長后期出現的肥胖問題上,在動物實驗研究中也已經初步證實了脂質的結構與后期脂肪組織堆積的相關性[3]。因此分析嬰兒配方乳粉與母乳中脂質存在的差異,努力將其縮小是嬰兒配方乳粉發展的必然趨勢。本文對脂質母乳化進展進行總結并提出展望,旨在為嬰兒配方乳粉的研發生產提供一定的理論依據。
1 母乳脂肪的組成及生物活性
1.1 乳脂肪的組成和結構
乳脂肪是一種最復雜的天然脂質混合物,其以脂肪球的形式穩定地存在于乳中。如圖1所示,乳脂肪球是以甘油三酯、膽甾醇酯等中性脂為核芯,并被特殊生物膜包裹的球狀組合體,粒徑分布在0.1~10.0 μm之間,平均粒徑為4 μm[4],比表面積約為2 m2/g[5]。甘油三酯約占總脂肪的98%,其以甘油為骨架,在3個位置分別酯化3個脂肪酸。目前已經在人乳中發現超過400種甘油三酯及200種脂肪酸。除甘油三酯外,乳脂中還含有約1%的磷脂、約0.3%的甾醇以及一些微量脂類,如甘油二酯、甘油一酯、游離脂肪酸等[6]。包裹在脂滴表面的特殊生物膜被稱為乳脂肪球膜(milk fat globule membrane,MFGM),它具有來自乳腺細胞內質網及頂端質膜的3 層膜結構,乳中絕大多數極性脂質位于MFGM上。研究顯示MFGM上的極性物質對嬰兒的神經發育、免疫功能、腸道功能及菌群的建立有特殊的有益作用[7-9],并且MFGM對脂滴的包裹也會對脂肪的消化吸收產生影響[10]。
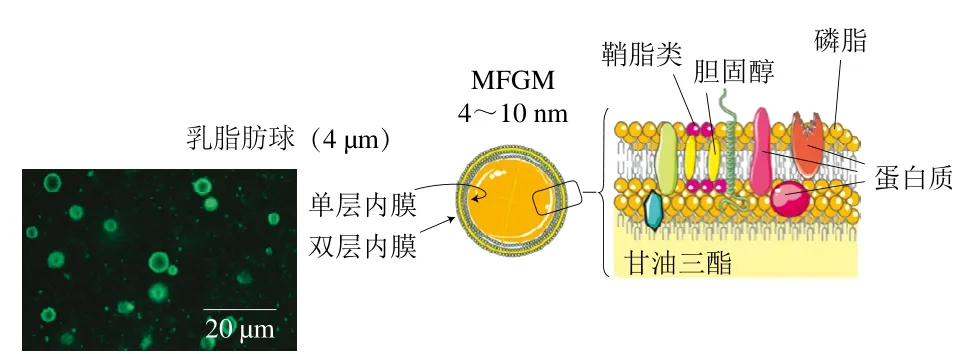
圖1 乳脂肪球結構示意圖[4]Fig. 1 Schematic drawing of the structure of the human milk fat globule[4]
1.1.1 脂肪酸的組成
母乳中脂肪酸主要以飽和脂肪酸為主[11],約占總量的44%~56%,其中含量最高的是棕櫚酸,其次是硬脂酸、月桂酸和肉豆蔻酸等,以油酸為主的單不飽和脂肪酸約占脂肪酸總量的31%~43%,與長鏈飽和脂肪酸相比,不飽和脂肪酸更容易被消化吸收。母乳中多不飽和脂肪酸的相對含量較低(11%~14%),且以亞油酸和亞麻酸為主。鏈長超過20個碳的多不飽和脂肪酸被稱為長鏈多不飽和脂肪酸(long chain polyunsaturated fatty acids,LC-PUFAs),根據最后一個雙鍵相對于分子末端的位置,LC-PUFAs又被分為n-3和n-6兩種類型[12]。母乳中最常見的n-6 LC-PUFAs為花生四烯酸(arachidonic acid,ARA),而常見的n-3 LC-PUFAs包括二十二碳六烯酸(docosahexaenoic acid,DHA)、二十二碳五烯酸(docosapentaenoic acid,DPA)和二十碳五烯酸(eicosapentaenoic acid,EPA)等,它們除了供能外還對嬰兒大腦、視力等方面起到有益作用,因而也被稱為功能性脂肪酸。乳中LC-PUFAs的含量雖然不高,但它們對嬰兒早期智力發育起著不可忽視的作用。
母乳中LC-PUFAs的含量受妊娠期、泌乳期、飲食習慣等多種因素的影響。初乳中LC-PUFAs含量高于成熟乳,早產母乳中DHA的含量也略高于足月母乳。母乳中的LC-PUFAs有兩個主要的來源,一是從飲食中直接獲取轉換,亞洲及一些沿海國家人民由于食用富含DHA的海魚較多,從而導致母乳有較高含量的DHA[13];二是內源性合成,即由體內的必需脂肪酸(亞油酸和亞麻酸)經延長酶和去飽和酶的作用合成相應的LC-PUFAs(圖2[1])。由于n-6 LC-PUFAs和n-3 LC-PUFAs的合成途徑存在競爭關系,因此嬰兒配方乳粉在設計脂肪配比時要特別注意亞油酸和亞麻酸的比例問題。
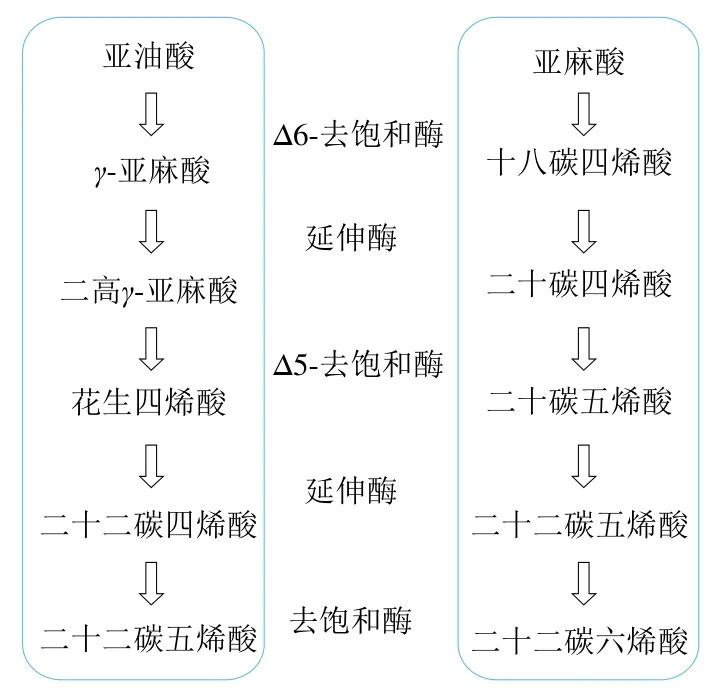
圖2 LC-PUFAs的內源性合成過程[1]Fig. 2 Endogenous synthesis process of long chain polyunsaturated fatty acids (LC-PUFAs)[1]
1.1.2 甘油三酯的組成
脂肪酸以與甘油骨架發生酯化反應形成甘油三酯的形式存在于母乳脂中,且其具有獨特的分布特點,以棕櫚酸為主的飽和脂肪酸大多酯化在甘油三酯的sn-2位[11],而不飽和脂肪酸主要分布在sn-1,3位,如母乳中含量最高的兩種甘油三酯——1,3-二油酸-2-棕櫚酸甘油三酯(1,3-dioleoyl-2-palmitoyl triglyceride,OPO)和1-油酸-2-棕櫚酸-3-亞油酸甘油三酯(1-oleoyl-2-palmitoyl-3-linoleoyl triglyceride,OPL)。研究表明具有sn-2棕櫚酸結構的甘油三酯更有益于嬰兒的消化吸收,而這種sn-2棕櫚酸很少存在于天然植物油中,其他動物乳脂中其含量也明顯低于人乳。同時LC-PUFAs也主要分布在甘油三酯的sn-2位[14](60% DHA和45% ARA),此位置的LCPUFAs具有更高的生物利用性[15]。此外,母乳脂中含有接近20%的中長鏈甘油三酯,即中鏈脂肪酸和長鏈脂肪酸酯化在同一個甘油骨架上,例如1,3-二油酸-2-月桂酸甘油酯(1,3-dioleicoyl-2-laurateoyl triglyceride,OLaO)(甘油三酯3個位置的脂肪酸組成:C18:1-C12:0-C18:1)、1-油酸-2-棕櫚酸-3-月桂酸甘油酯(1-oleoyl-2-palmitoyl-3-laurateoyl triglyceride,OPLa)(甘油三酯3個位置的脂肪酸組成:C18:1-C16:0-C12:0)和1,2-二肉豆蔻酸-3-油酸甘油酯(1,2-dimyristoyl-3-oleoyl triglyceride,MMO)(甘油三酯3個位置的脂肪酸組成:C14:0-C14:0-C18:1)等,這種甘油三酯兼具中鏈甘油三酯和長鏈甘油三酯的優點,可以快速供能,滿足早產兒及患有吸收不良疾病嬰兒的能量需求,同時還可避免有毒代謝物的產生[16]。
1.1.3 MFGM的組成及結構
MFGM的包裹使乳脂以脂肪球的形式穩定存在于母乳中,MFGM約占脂肪球總質量的2%~6%,是由極性脂質、膽固醇、膜特異性蛋白等組成的復雜體系。MFGM含有乳中絕大多數極性脂質,包括磷脂酰膽堿(phosphatidylcholine,PC)、磷脂酰乙醇胺(phosphatidylethanolamine,PE)、磷脂酰絲氨酸(phosphatidylserine,PS)、磷脂酰肌醇(phosphatidylinositol,PI)、鞘脂類以及微量的溶血磷脂和縮醛磷脂等[17]。鞘磷脂(sphingomyelin,SM)是含量最高的鞘脂,同時也是人乳中含量最高的極性脂質[18]。其次為PC和PE,而PS和PI的含量最低。MFGM蛋白約占MFGM總量的25%~70%,基于蛋白組學,目前已經檢測出數百種MFGM蛋白,較為常見的有嗜乳脂蛋白(butyrophilin,BTN)、氧化還原酶(xanthine oxidase,XO)、黃嘌呤脫氫酶(xanthine dehydrogenas,XDH)、黏蛋白等。蛋白質在MFGM上的分布是不對稱的,有些蛋白穿插在各層之間起到物質和信息傳遞的作用,而大部分蛋白質被糖基化分布在膜的表面,并發揮著功能性作用,如對有害菌的抑制等[19]。
在不破壞膜的情況下,利用激光共聚焦及原子力顯微鏡對膜的形態進行表征,能夠揭示SM的橫向分離性及膜的機械異質性,即極性脂在膜外表面是非均勻分布的,至少有兩相存在,包括由SM與膽固醇形成的有序相(又被稱為“脂筏”結構)和由甘油磷脂(PC、PE、PS、PI)形成的無序相。且有序相通常比環繞它的無序相厚度略大,這就導致酰基鏈之間的錯配和機械異質性相界的存在,可能對消化酶的吸附和插入產生影響[20]。SM中的脂肪酸大多是長鏈飽和脂肪酸,使得有序相具有維持MFGM穩定性和剛性結構的作用,同時保護MFGM中的活性物質不被消化。與之相反,無序相中多含不飽和脂肪酸,這使MFGM具有一定流動性的同時更易于與脂肪酶結合。極性脂質在膜整體上的分布也是不均勻的,帶負電荷的PI和PS多分布在最內層的單層膜上,PE主要分布在雙層膜的內層,而PC和SM是雙層膜外膜的主要極性脂[21],這種分布對MFGM的微觀結構有很大的影響,可以穩定脂肪球的脂質核心,保護其免受酶的分解[22]。
1.2 母乳脂質的生物活性
1.2.1 LC-PUFAs的生物功能
DHA、ARA及EPA這幾種LC-PUFAs以較高的濃度存在于人類的中樞神經組織,尤其是一些特定的大腦區域和視網膜細胞膜中[23],它們的快速積累主要發生在母親妊娠的最后3個月和嬰兒出生后的前兩年[24-25],即嬰兒腦部快速發育時期,這說明其對嬰兒神經及意識的發育可能具有重要作用。LC-PUFAs是神經細胞膜的重要組成部分,已經被證實其可以通過促進神經干細胞分化、促進突觸形成并改善突觸傳遞機制從而促進大腦的發育[26],DHA是視網膜脂質的重要結構成分,占視桿細胞和視錐細胞中總脂肪酸含量的50%以上,對嬰兒視力的發育至關重要[27]。產后哺乳時期是新生兒獲得免疫功能的關鍵時期,研究顯示母乳中的LC-PUFAs對嬰兒免疫系統的完善也具有重要作用[28]。
過去30 年間,LC-PUFAs對嬰兒大腦、視力、免疫發育的影響廣泛被人們所關注,并已經進行了許多臨床驗證試驗。本文選取部分有代表性的研究列于表1中。其中隨機雙盲對照實驗結果表明通過向哺乳期母親補充LC-PUFAs來提高母乳中DHA的濃度,經過一段時間的喂養,有利于嬰兒神經認知的發育,并且提高母乳中DHA的濃度還被證明可以改善嬰兒視力發育以及提高嬰兒的持久注意力和解決問題的能力[29-31]。有關嬰兒免疫發育的研究也表明LC-PUFAs的補充可以降低嬰兒出生后哮喘和過敏患病率,推遲患病時間[32],并可使嬰兒體內的免疫細胞(免疫T細胞和B細胞)和細胞因子(腫瘤壞死因子)濃度更接近于母乳喂養的嬰兒,有利于嬰兒免疫體系的完善[28]。另外通過掃描視覺誘發電位來測試嬰兒視力的臨床試驗,也表明長期LC-PUFAs的補充可以明顯改善嬰兒的視力發育[33]。

表1 LC-PUFAs對嬰兒發育產生影響的臨床試驗總結Table 1 Summary of clinical trials on the effects of LC-PUFAs on infant development
1.2.2sn-2棕櫚酸的生物益處
研究表明富含sn-2棕櫚酸的甘油三酯更加容易被嬰兒消化吸收,這得益于胃腸脂肪酶對甘油三酯特殊的水解選擇性,其可專一水解sn-1/3位形成兩分子游離脂肪酸和2-甘油一酯,相比于長鏈、飽和的脂肪酸,中短鏈、不飽和的脂肪酸更加容易被腸道直接吸收,而2-甘油一酯也以特殊的機制重新形成甘油三酯后被利用。這種形式的甘油三酯除了可以提高脂質的消化率,還可以避免鈣皂的形成(圖3),從而提高嬰兒的胃腸舒適度和鈣的利用率[34]。
目前已經有很多臨床研究證實了sn-2棕櫚酸的積極作用。最近一項大型(n=488)隨機、雙盲、多中心臨床試驗表明富含sn-2棕櫚酸的嬰兒配方乳粉可明顯改善嬰兒2周至2個月的糞便稠度,并增加嬰兒4個月時骨骼礦物質含量[34]。Padial-Jaudenes等[36]的綜述也進一步證實了sn-2棕櫚酸與鈣吸收和糞便硬度之間存在相關性。此外它還被證明與嬰兒哭泣次數的減少及腸道發育的成熟有關。Litmanovitz等[37]的隨機、雙盲臨床試驗結果表明,與普通乳粉相比,喂養富含sn-2棕櫚酸的嬰兒配方乳粉可以明顯降低嬰兒在下午和晚上哭鬧的頻率及縮短持續時間,這可能歸因于sn-2棕櫚酸提高了嬰兒胃腸的舒適度,也可能是由于sn-2棕櫚酸可以通過復雜的機制幫助兒童建立晝夜節律,并對神經內分泌介質和調節系統產生積極影響[38]。sn-2棕櫚酸還可以通過影響進入腸細胞的游離脂肪酸和sn-2甘油一酯的組成,來影響腸道成熟及菌群結構[38]。一項臨床研究發現喂養富含sn-2棕櫚酸的嬰兒配方乳粉會使嬰兒糞便中乳酸桿菌和雙歧桿菌數量增加,伴隨著對病原菌的拮抗作用和積極的免疫調節,其可以促進腸道成熟[39]。綜上,現有研究已一定程度上證實了sn-2棕櫚酸的益生作用,但仍然需要設計更全面、更細致的實驗來證明sn-2棕櫚酸對嬰兒發育的有益作用。
1.2.3 MFGM的營養功能
MFGM中的PI和PE被證明含有較高含量的LCPUFAs,可以為嬰兒提供生長所需的EPA和DHA,同時PC和SM是嬰兒主要的膽堿來源,膽堿是神經遞質乙酰膽堿的前體,它在代謝過程中可以作為甲基供體調節轉導信號,從而被認為是大腦發育所必需的營養物質[40]。由于脂筏區的存在,SM被證明不能被完全消化,可以部分抵達遠腸端,動物實驗結果顯示這可以加速幼鼠小腸的成熟[41]。另一項對大鼠的研究也表明,SM可以促進大鼠神經系統髓鞘的發育,其代謝產物鞘氨醇-1-磷酸也被證明對免疫功能起著重要的調節作用[42]。另外,SM可以與膽固醇作用形成復合體,從而減少腸道對膽固醇的吸收,這可能也是母乳喂養的嬰兒成年后心血管疾病患病率低的原因之一[43]。考慮到神經組織中神經節苷脂含量高,母乳中僅存在于MFGM上的神經節苷脂可能對嬰兒的神經意識發育起著無法替代的作用,此外它還被證明可以捕獲病原體,防止嬰兒感染,并調節免疫細胞行為,增強嬰兒的免疫力[44]。近年來,越來越多的隨機雙盲試驗旨在探究嬰兒飲食中MFGM的添加對嬰兒產生的具體影響。由表2可知,MFGM成分的補充會促進嬰兒神經及智力的發育,降低嬰兒腹瀉發生率,提高嬰兒免疫力,并且無明顯不良情況的發生,這說明嬰兒配方乳粉中MFGM的添加對嬰兒發育是有利且安全的。
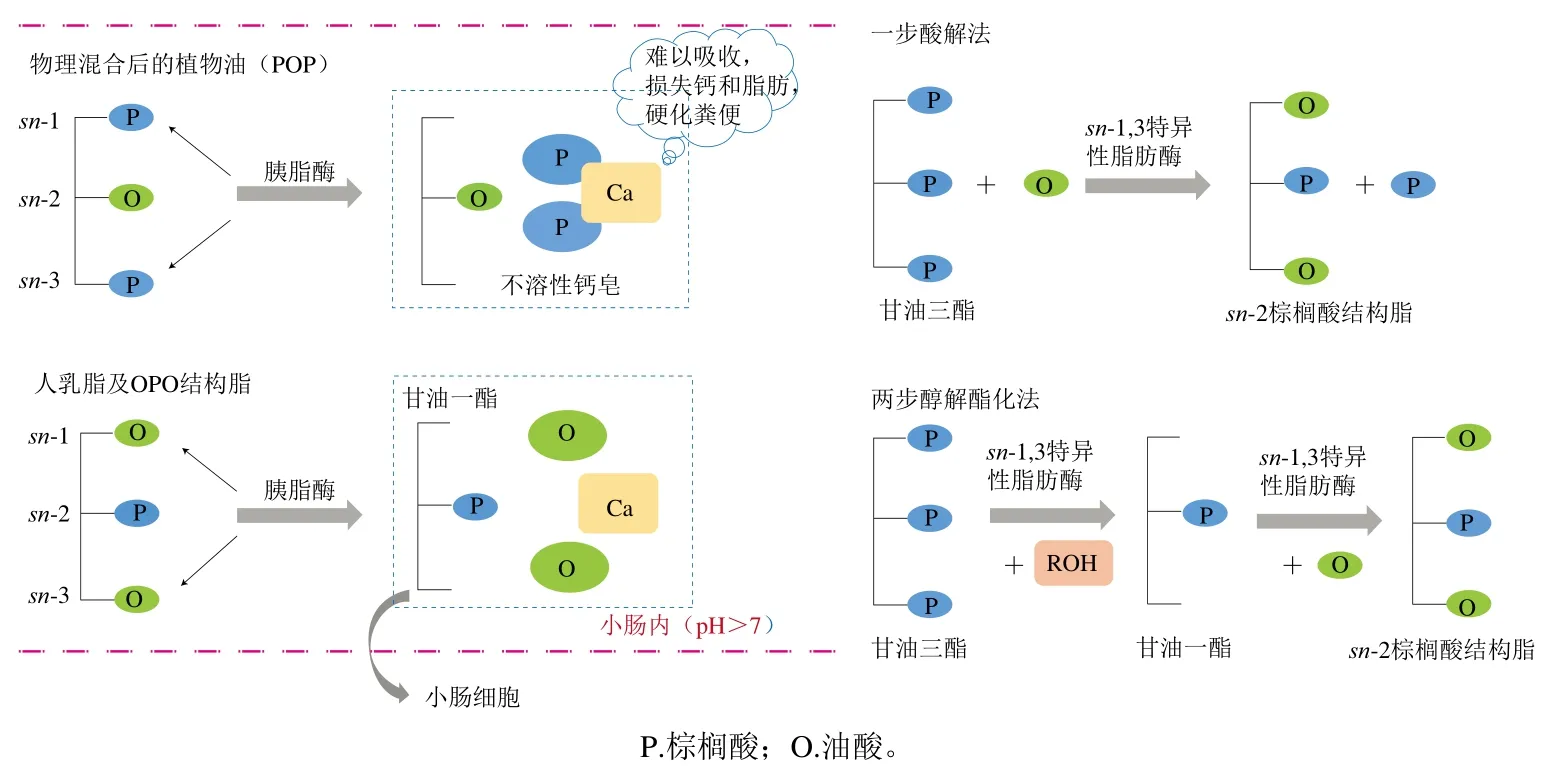
圖3 sn-2棕櫚酸結構脂促進鈣和脂肪吸收的原理及其酶法合成途徑[35]Fig. 3 Promoting effect of structured triacylglycerols rich in palmitic acid at sn-2 position on calcium and fat absorption and its enzymatic synthesis pathway[35]
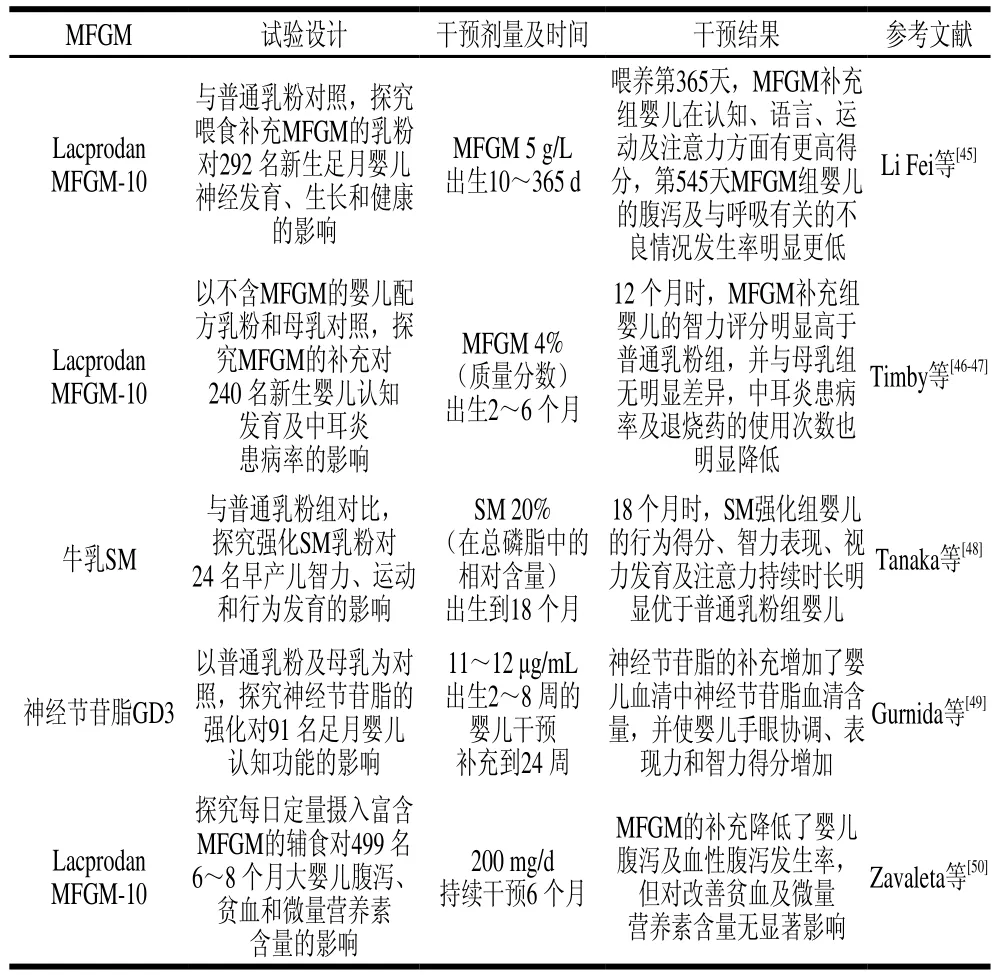
表2 嬰兒配方乳粉中添加MFGM對嬰兒發育影響的臨床研究Table 2 Clinical research on the effect of dietary supplementation of MFGM on infant development
研究表明脂滴的界面結構和有效表面積對脂肪消化代謝有很大的影響,新生兒還不具備成熟的消化系統,卻可以很好地吸收乳中的脂質,說明MFGM的包裹有利于嬰兒對脂質的消化,這可能是因為MFGM上存在胃腸脂肪酶特定的結合位點[51]。國外已經有制備MFGM包裹大粒徑脂滴的先例,達能集團利用MFGM中的磷脂優化包封工藝制備出含有與母乳脂肪球結構類似的新型嬰兒配方乳粉并進行臨床測試,發現這種新型乳粉更有利于嬰兒對脂質的消化吸收[52]。Baars等[3]以小鼠為研究對象,發現相比于普通乳粉,新型乳粉可以降低小鼠成年期的脂肪蓄積并改善代謝特征,從而說明其可能對嬰兒后天肥胖率產生影響。Breij等[53]通過隨機雙盲臨床試驗,發現食用新型乳粉組的嬰兒有更低的日增長體質量,排便情況也更加接近母乳喂養組,并且沒有明顯不良情況發生。
2 脂質原料與母乳脂的關鍵差異
通過對多種動物乳成分進行對比分析及受到產量的限制,目前牛乳和羊乳是嬰兒配方乳粉生產的主要原料,但在乳脂組成方面,它們與人乳脂還存在很大的差異。由表3可知,牛羊乳脂中的飽和脂肪酸含量明顯高于母乳,而母乳中含有更多的單不飽和脂肪酸和多不飽和脂肪酸[11]。同時,相比于母乳,牛羊乳脂中短鏈脂肪酸含量更高,這可能是反芻動物特殊的胃消化作用而造成的;但對脂質消化產生重要影響的sn-2棕櫚酸在牛羊乳中的含量卻明顯低于人乳[11]。另外,對于甘油三酯,母乳中含量最高且具有重要生理功能的兩種甘油三酯(OPO、OPL)在牛羊乳脂中含量很低,且牛羊乳中幾乎不含人乳中常見的中長鏈甘油三酯[54],這些乳脂分子水平的差異會對脂質的消化吸收產生影響,這也說明了天然的牛羊乳脂不是嬰兒最佳的脂質來源。
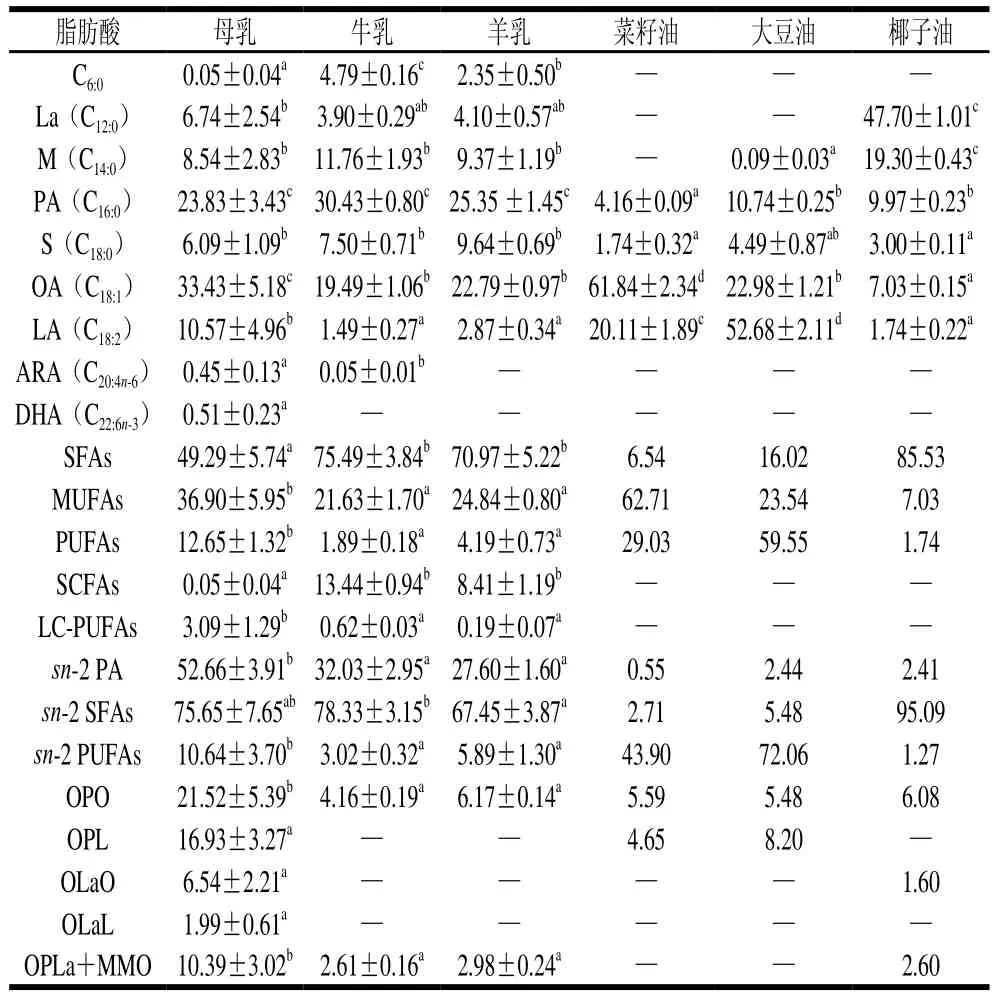
表3 人乳與嬰兒配方乳粉原料油脂質成分對比[11,55]Table 3 Comparison of lipid composition of human milk and raw oil used in infant formula[11,55]
植物油種類繁多,且幾乎所有母乳脂中的脂肪酸都能在植物油中找到,因此為了彌補動物乳脂的不足,目前市面上的嬰兒配方乳粉大多是以多種植物油經過物理混合來模擬母乳脂組成,也有部分嬰兒配方乳粉中添加了動物乳脂。調查顯示80%的進口乳粉采用純植物油,而75%國內乳粉則是將乳和植物油進行調配使用,主要的乳源為牛乳[56]。乳粉中所用植物油主要包括油酸型(如高油酸葵花籽油、菜籽油等)、油酸亞油酸型(如大豆油、玉米油、葵花籽油、花生油等)和中長鏈脂肪酸型(如棕櫚仁油、椰子油等)[55],植物油可以彌補牛羊乳不飽和脂肪酸含量的不足,使混合油脂在脂肪酸組成上更接近于母乳。還有一些產品會通過添加部分深海魚油、海藻油或微生物發酵產生的油脂來補充LC-PUFAs[56]。
嬰兒配方乳粉中的脂質是混合油脂經過高壓均質后,被乳中蛋白質包裹形成的小脂滴,因此與母乳脂肪球相比,具有更小的粒徑(0.3~0.5 μm),且缺少MFGM的包裹。在消化過程中,脂滴表面的蛋白質容易發生聚集,從而阻止脂肪酶與脂質的有效接觸,降低脂肪的消化率,使嬰兒出現脂質利用不足的現象[57]。MFGM的缺乏除了對消化產生影響,還會減少嬰兒對很多生物活性物質的獲取,如極性脂質和一些功能性膜蛋白。
混合油脂可以很好地模擬母乳中常量脂肪酸組成,卻無法對母乳中甘油三酯和脂肪球結構進行模擬。而嬰兒配方乳粉喂養的嬰兒胃腸舒適度、免疫力及早期表現能力與母乳喂養的嬰兒還存在差距,這也說明僅僅在脂肪酸層面對母乳脂肪進行模擬還不能完全滿足嬰兒對脂質的需求。功能性脂肪酸和MFGM成分的適量添加、甘油三酯及脂肪球結構的模擬可能是縮小嬰兒配方乳粉與母乳之間差異的突破點,也是目前嬰兒配方乳粉脂質母乳化的重點。因此,接下來本文就這幾方面的研究進展進行總結,并提出展望。
3 嬰兒配方乳粉脂質母乳化模擬途徑及策略
3.1 LC-PUFAs組成的模擬
嬰兒剛出生時體內的酶系發育不健全,自身不能合成足夠的LC-PUFAs,必須從飲食中獲取。從20世紀90年代開始,DHA和ARA陸續被添加到嬰兒配方乳粉中,2016年Fu Yuanqing等[58]對來自41個國家或地區的4 163 份母乳樣品數據進行總結,得出全球母乳DHA和ARA的平均水平分別為(0.37±0.11)%和(0.55±0.14)%(在總脂肪酸中的相對含量)。基于人乳中DHA和ARA的平均水平,嬰兒在6個月大時每天應攝入大約190 mg ARA和130 mg DHA[59]。兒童健康基金會也建議嬰幼兒每天應至少攝入140 mg ARA及100 mg DHA來維持大腦及視力的發育[60]。但多個國家的研究顯示絕大部分乳粉中LC-PUFAs的添加量沒有達到母乳水平,Loughrill等[61]對4個品牌嬰兒配方乳粉中LC-PUFAs的含量進行檢測,并結合6~9個月嬰兒每日乳粉的推薦食用量進行干預,發現嬰兒配方乳粉僅能為嬰兒提供19.18%每日所需的DHA以及21.56%每日所需的ARA。
LC-PUFAs最好的來源是海洋生物,尤其是多脂魚類。魚油中一般含有DHA、EPA,以及少量的ARA和DPA,并且它們的組成受魚類的飲食、位置、季節和生理條件(如物種的年齡和性別)的調節[62]。磷蝦油也被證明是n-3 LC-PUFAs良好的來源,磷蝦油由磷脂(占30%~65%)和甘油三酯組成,而磷脂中40%的脂肪酸為DHA和EPA[63]。近來研究發現海洋魚油高LC-PUFAs含量的原因是海魚以消耗微藻的浮游生物為食,而微藻具有合成和積累大量LC-PUFAs的能力,另外,一些細菌、真菌等微生物也被證明可以合成LC-PUFAs,因此人們逐漸把目光投向利用真菌和微藻開發新的細胞工程策略,以實現LC-PUFAs的可持續生產[64]。這些單細胞LC-PUFAs的提純,通常是先利用超聲、微波等手段將細胞破壞,再選用分子蒸餾、超臨界流體萃取等多種方法進行純化,并加入適量的抗氧化劑防止氧化[65]。該LC-PUFAs已經在歐洲接受了全面的安全評估,Morris等[66]的研究也證實了單細胞來源的LC-PUFAs對嬰兒的生長和總體健康沒有任何不良影響。
食品法典規定嬰兒配方乳粉中DHA的添加量不能超過總脂肪酸的0.5%,并且必須添加至少等量的ARA,同時要求EPA添加量不能超過DHA。LC-PUFAs由于其結構中含有多個不飽和雙鍵而容易被氧化,從而使其喪失活性功能并產生不良氣味,并且作為豐富LC-PUFAs來源的魚油脂本身就帶有海腥味,這限制了食品中LC-PUFAs的使用。嬰兒配方乳粉中LC-PUFAs的添加多采用微膠囊技術,即以天然或合成的高分子材料為壁材,通過化學法、物理法或物理化學法將核芯材料(活性物質)進行包埋,從而實現對內容物的保護,掩蓋內容物的不良風味并在特定條件下以受控的速度將其釋放。可以實現微膠囊化的方法有很多,常用于食品業的主要有噴霧干燥法、噴霧冷凝法、冷凍干燥法、復凝聚法等[67],其中應用最為廣泛的是噴霧干燥法,其原理是乳液霧化后,利用高溫氣流使溶劑迅速蒸發,最終使壁材在芯材表面形成膜結構,其具有成本低廉、工藝條件容易控制、產品顆粒均勻且溶解性好等特點[68]。對于嬰兒配方乳粉中LC-PUFAs的微膠囊化,壁材應該選用適合嬰兒的食品級成分,優選天然成分。乳蛋白、乳蛋白-碳水化合物混合物及乳蛋白和碳水化合物的美拉德產物都是用作LC-PUFAs微膠囊壁材的良好原料[69]。為防止微膠囊化過程中LC-PUFAs的氧化,可以在均質、乳化等步驟中添加一些天然抗氧化成分,如生育酚等抗氧化劑,這可以進一步提高嬰兒配方乳粉中LC-PUFAs的穩定性和利用率。
3.2 sn-2棕櫚酸結構脂的組成模擬
與70%的棕櫚酸位于母乳甘油三酯sn-2位這種特性不同,作為嬰兒配方乳粉脂質原料的植物油中棕櫚酸主要位于甘油骨架的sn-1,3位,這會影響嬰兒對脂質的消化吸收。因此選擇合適的脂肪酶對原料油進行酶法修飾,合成sn-2棕櫚酸結構脂并應用于嬰兒配方乳粉,是目前脂質在甘油三酯層面上母乳化的一個重點。Betapol?和Infat?即為兩種典型的sn-2棕櫚酸結構脂,已經應用于嬰兒配方乳粉的生產。與傳統的化學催化劑相比,結構脂酶法修飾所用到的脂肪酶具有特異選擇性強、操作條件溫和、催化效率高等優點,且酶催化過程對環境友好[70]。目前在合成中應用較為廣泛的商業脂肪酶是Novozymes公司生產的Lipozyme RM IM和Lipozyme TL IM。它們作為一種sn-1,3特異性脂肪酶,不僅能對甘油三酯sn-1,3脂肪酸進行專一水解,還能催化甘油三酯的酯化、酸解或醇解等合成反應[71]。在選擇合適的底物和酰基供體之后,反應可以在簡單的玻璃容器或反應釜中進行。
通常sn-2棕櫚酸結構脂的合成有兩種反應途徑:一步酶法反應和兩步酶法反應(圖3)。一步酶法反應又分為酸解法和酯交換法,酸解法即甘油三酯與一種或多種游離脂肪酸在脂肪酶的催化作用下直接進行酰基交換反應合成結構脂的過程[72],該法操作簡單、反應產物種類較少且易于分離,是目前最常用的合成方法。聯合利華的Betapol?即由sn-1,3特異性脂肪酶通過一步酶法酸解富含三棕櫚酸甘油酯的植物油與油酸,生成富含OPO的結構脂。酯交換反應是指兩種不同甘油三酯或簡單酰基酯與甘油三酯經脂肪酶催化,發生酶促轉酯反應合成結構脂[73]。由于該法專一性不強、合成效率不高且反應產物不易分離,因而不如酸解法應用廣泛。兩步酶解法又稱為醇解酯化法,其是將sn-2位富含棕櫚酸的甘油三酯與醇類(主要是乙醇)反應生成2-甘油一酯,接著將2-甘油一酯與脂肪酸經sn-1,3特異性脂肪酶催化合成結構脂[74],此法合成效率高,產物純度也更高,但由于進行了兩次酶法催化,提高了反應成本。sn-2棕櫚酸結構脂的合成,除了選擇合適的反應途徑,脂肪酶活性、特異性和載體、反應溫度、反應時間、底物、水分含量以及反應器類型也是提高生產率并降低生產成本需要考慮的因素。
3.3 MFGM及脂肪球結構的模擬
近年來,隨著對MFGM關注度的提高,已經有很多企業將牛乳來源的MFGM作為營養強化劑添加到嬰兒配方乳粉中,動物及臨床試驗都顯示出積極作用,且無明顯不良情況發生。目前商業化MFGM大多是從在牛乳或牛乳加工副產物中提取,經過乳脂肪球分離、脂肪球的洗滌、釋放MFGM、收集MFGM材料這4個步驟來獲取純度較高的MFGM成分[75]。酪乳、黃油乳清、β-乳清和干酪乳清這些牛奶加工的副產物都是MFGM材料的合適來源[3]。但牛乳MFGM在成分和結構上與人乳還存在一定的差異,Wang Lina等[76]研究發現,與牛乳相比,人乳中含有更高比例的SM、PS和PI,而牛乳中PC的比例更高,人乳磷脂酰基鏈的平均不飽和程度高于牛乳,特別是以DHA為代表的n-3 LC-PUFAs。MFGM的微觀結構與磷脂和脂肪酸組成有關,磷脂種類和比例的不同以及脂肪酸在磷脂上的分布都可能影響MFGM的微觀結構和營養價值。Bourlieu等[77]的研究也證明了人乳MFGM中SM含量高,且人乳MFGM中含有更高比例的PI和PS,這兩種磷脂是帶負電荷的陰離子磷脂,研究顯示較高含量的陰離子磷脂(PI和PS)可能通過靜電作用促進胃脂肪酶對MFGM的定向和吸附。通過對比人乳MFGM和牛乳MFGM的物理化學性質,研究人員發現人乳MFGM具有更高的可壓縮性,這與其較高的不飽和脂肪酸含量有關,進一步可能對脂肪酶在MFGM表面的插入產生影響[78]。因此,MFGM的模擬在整體成分添加的基礎上,也應注意單個極性脂的比例,使其更加接近母乳MFGM。目前不同生產企業所用原料及加工技術有所不同,導致不同MFGM產品組成存在較大差異[79],且單一極性脂的提純還很難達到。因此,更加全面地分析各種乳中MFGM組成及差異,提高MFGM的提純工藝,按照母乳MFGM中各極性脂的比例進行調配,是MFGM成分方面模擬的重點。
MFGM在確保乳脂肪穩定的同時也會影響嬰兒對脂質的消化代謝。因此,目前除了對MFGM生物功能的研究,脂肪球結構的模擬也成為研究人員關注的重點。近年來,被MFGM包裹的大脂滴已經可以通過將MFGM片段與甘油三酯混合均質而成功制備。與乳粉中被蛋白質包被的脂滴相比,它們在結構上更接近人乳脂肪球。Lopez等[80]利用黃油乳清制備的MFGM涂層脂滴物理性質穩定,1個月后沒有觀察到聚集,這可以解釋為界面MFGM組分在脂滴之間的靜電和空間排斥力中起到了有益作用。由此可見,制備MFGM涂層脂滴模擬乳脂球是可行的,且脂滴的大小可以通過制備過程中施加的均質壓力進行調節。通過顯微技術觀察發現,在制備過程中添加的MFGM碎片和MFGM組分(如極性脂質、蛋白質)能夠在均質過程中移動到脂滴表面并吸附上去,且在一定的壓力范圍內,MFGM碎片不會被破壞。MFGM成分中的蛋白質也會吸附在脂滴表面,因此在制備乳液的過程中,需要考慮原料的引入順序和極性脂與蛋白質的比例,因為它們會強烈地影響脂滴表面的組成和結構,以及脂滴的功能特性[81]。目前還沒有模擬出母乳MFGM的3 層膜結構,且尚不清楚單層膜脂滴會在多大程度上對嬰兒的成長產生影響,以及是否會轉化為長期效應。因此,對母乳脂肪球結構的進一步模擬及對嬰兒成長的影響結果還需進行深入研究。
4 結 語
過去的20 年,在確定母乳成分和對嬰兒的營養作用方面已經取得了巨大成就,對于嬰兒配方乳粉脂質的母乳化也取得了一定的進展。目前以牛羊乳為乳基料并選擇多種植物油復配可以很好地實現母乳脂肪酸層面的模擬,并且可以通過添加部分深海魚油或海藻油來滿足嬰兒對LC-PUFAs的需求。對于甘油三酯層面的模擬也在逐步向前推進,OPO結構脂的添加使嬰兒可以更好地吸收鈣和脂肪,同時提高嬰兒的胃腸舒適度。但由于母乳脂肪甘油三酯的種類繁多、結構復雜,除OPO結構脂的使用之外,在母乳甘油三酯方面上更深層次的模擬也必須重視起來,例如中長鏈甘油三酯、sn-2 LC-PUFAs甘油三酯種類和含量的模擬,以及進一步提高嬰兒配方乳粉中sn-2棕櫚酸的含量。此外,母乳中MFGM的存在也被證明有利于嬰兒大腦及免疫的發育,脂肪球的結構還對嬰兒脂肪吸收和代謝產生影響,而這兩者正是目前大部分嬰兒配方乳粉所欠缺的。因此,優化MFGM的制備工藝,尋找最佳的人乳MFGM替代品,利用MFGM包裹脂滴來達到對乳脂肪球結構的模擬,是未來新型嬰兒配方乳粉研究和開發的重點。
