細胞焦亡調控腫瘤免疫微環境的分子機制研究進展
2022-05-16 14:00:06黃小俠
中國農業大學學報 2022年6期
董 婧 王 耀 黃小俠 李 林
(沈陽農業大學 動物科學與醫學學院,沈陽 110866)
犬貓作為伴侶動物,深受大眾喜愛,隨著飼養條件和寵物診療技術的進步,壽命逐漸延長,但寵物腫瘤的發病率逐年增加,給寵主帶來巨大的經濟壓力和心理壓力。我國犬的腫瘤病例數量呈逐年增長趨勢,數據顯示在全國范圍內,1983—2000年有536例,2000—2012年有1 163例,2018—2020年沈陽市寵物醫院收治犬腫瘤病例數量達1 225例。癌癥是犬最常見的死因之一,美國的一項犬尸檢統計報告顯示,2 002只犬中,23%死于癌癥,而10歲以上高達45%。我國寵物臨床上,犬乳腺腫瘤的發生概率最高,平均發病年齡9歲左右,是寵物腫瘤的主要類型,位居第二位的是皮膚腫瘤。犬乳腺腫瘤病例中,約有41%~53%為惡性,并有轉移傾向,而貓乳腺腫瘤中85%以上為惡性或已轉移。良性腫瘤可采用手術切除,惡性腫瘤即使采用手術或化療藥物治療,也會伴隨預后不良及高復發率。因此,尋找更優質的腫瘤防治方案成為獸醫工作者面臨的急需解決的難題。近年來對患癌犬的治療中,腫瘤免疫調節成為一種有效的治療策略,能有效延長患犬的生存期。
在腫瘤免疫微環境中,腫瘤細胞會受到間質細胞、間質外細胞、細胞因子、趨化因子和代謝產物等的保護,使得腫瘤細胞能夠得以生存。細胞焦亡可以在腫瘤免疫微環境中對腫瘤細胞產生抑制和促進兩種作用,既可以通過細胞滲透性裂解后釋放出來的損傷相關分子模式(DAMPs)將腫瘤免疫微環境重新編程為免疫刺激狀態,從而抑制腫瘤細胞生長和轉移;又可以在炎癥因子的作用下,促進腫瘤細胞生長。細胞焦亡可能會對腫瘤免疫微環境產生特定影響并促進免疫監視。越來越多的研究表明,可以通過化學藥物治療誘導使腫瘤細胞發生焦亡,如臨床常用的紫杉醇、順鉑和阿霉素等。誘導細胞焦亡及其調控腫瘤免疫微環境的研究為抗腫瘤治療提供了新思路。
1 細胞焦亡概述
細胞焦亡是由各種炎性體觸發的溶解性及程序性細胞死亡,通過誘導癌細胞死亡或引發抗癌免疫在癌癥進展中起重要作用。細胞發生焦亡時會釋放大量的細胞內容物,如促炎因子和溶酶體,從而產生炎癥級聯反應,其特點是細胞形態逐漸扁平,線粒體和溶酶體受損,細胞核濃縮,DNA斷裂降解,染色質凝聚,細胞膜形成膜孔,細胞逐漸膨脹至質膜破裂。細胞焦亡可以通過典型的含半胱氨酸的天冬氨酸蛋白水解酶-1(Caspase-1)炎性途徑和非經典的Caspase-4、5(人)和Caspase-11(鼠)途徑完成。同時,Wang等證明Caspase-3通過裂解Gasdermin E(GSDME)參與細胞焦亡的調控。細胞焦亡失調可能會導致病原體清除效率降低和功能紊亂,從而無法刺激適應性免疫防御系統,導致組織受損。焦亡通路的關鍵GSDMs在許多癌癥組織中處于沉默狀態,即無法發揮其功能,因此對細胞焦亡分子生物學機制的深入研究有望為寵物臨床提供一種前景廣闊的新型治療方法。
2 細胞焦亡的分子機制
2.1 介導細胞焦亡的關鍵因素
2
.1
.1
炎性小體炎性小體是由胞漿內識別受體(PRRs)介導組裝的多聚體蛋白復合物,為天然免疫系統的主要組成成分。炎性小體包含傳感器、適配器和酶原3個結構,種類多樣,主要有核苷酸寡聚化結構域(NOD)和富含亮氨酸重復序列(LRR)的受體NLR、黑色素瘤2(AIM2)樣受體ALR以及三聯基序(TRIM)家族的成員Pyrin/TRIM20。炎性小體能夠識別病原相關分子模式(PAMPs)和DAMPs,并由包括NLR、AIM2或Pyrin在內的傳感器來招募含有胱天蛋白酶募集結構域(CARD)的凋亡相關斑點樣蛋白(ASC),形成多聚體復合物,將Pro-caspase-1募集到復合物中形成活化的Caspase-1,誘導其切割GSDMD并促進白介素等細胞因子的成熟。有研究顯示,NLRP1b可被炭疽致死因子直接切割,誘導N末端降解激活Caspase-1。NOD樣受體(NLR)家族CARD結構域3(NLRC3)通過p38信號通路負調控樹突狀細胞(DCs)的抗原呈遞功能,可作為治療誘導過敏和自身免疫的誤導性適應性免疫反應的潛在靶標。NLRC4對細胞質鞭毛蛋白以及細菌3型分泌系統的蛋白有反應,防御腸道內外病原體的入侵。NLRP4被證實可以抑制腫瘤壞死因子-α(TNF-α)和白細胞介素1β(IL-1β)介導的核因子-κB(NF-κB)激活,是病毒感染過程中誘導免疫反應的關鍵介質。AIM2被證明有助于監控和清除DNA損傷,通過藥物靶向AIM2可能在放射治療期間具有益處。
2
.1
.2
Caspase家族Caspase家族是一組存在于細胞質溶膠中的半胱氨酸蛋白水解酶。Caspase以無活性的酶原狀態存在,在哺乳動物體內至少發現16種,其活化狀態特異性地裂解天冬氨酸殘基后的肽鍵,切割不同的底物來實現細胞凋亡和細胞焦亡等不同的細胞死亡類型。其中研究較為廣泛的Caspase可以分為介導細胞凋亡的啟動者Caspase-2、8、9、10和細胞凋亡的執行者Caspase-3、6、7以及介導細胞焦亡的Caspase-1、3、4、5、8、11。研究表明,介導細胞焦亡的Caspase成員可以被炎性小體激活,使細胞膜形成孔膜,進而實現對靶細胞的殺傷作用。
2
.1
.3
Gasdermin家族Gasdermins(GSDMs)是一類功能多樣、結構保守的蛋白家族,在胃腸道、皮膚和免疫細胞等多種組織細胞中表達。在人體內,GSDMs包含GSDMA-E和PJVK(或常染色體隱性耳聾59型,DFNB59)。在小鼠體內,GSDMs包括GSDMA的3個同源物(GSDMA1-3)、GSDMC的4個同源物(GSDMC1-4)、GSDMD-E和PJVK。GSDMs蛋白全長處于自抑制狀態,除了PJVK,均可由Caspases切割成N末端(GSDM-NT)和C末端(GSDM-CT)2個部分。GSDM-NT與細胞膜內小葉結合,形成內徑10~14 nm 的膜孔,破環細胞離子梯度,使直徑為4.5 nm 的IL-1β和7.5 nm的Caspase-1通過,誘導細胞焦亡。GSDMA主要在人的胃腸道、皮膚、食道和胃中表達。小鼠GSDMA3-NT與細胞膜上的心磷脂和磷酸肌醇相互作用形成孔,觸發細胞焦亡。GSDMB主要在胃腸道、肺、淋巴細胞、肝臟、結腸和食管上皮等組織中有高表達。顆粒酶A(GZMA)能切割GSDMB,釋放GSDMB-NT誘導細胞焦亡。Caspase-1在D236位點切割GSDMB也可釋放GSDMB-NT誘導細胞焦亡。另一項研究顯示,GSDMB可促進Caspase-4活性,與Caspase-4的CARD結構域結合,促進GSDMD釋放GSDMD-NT,誘導細胞焦亡。Chao等研究表明,凋亡執行者Caspase-3、6、7能夠切割GSDMB,釋放GSDMB-NT誘導細胞焦亡。GSDMC主要在人的胃腸道、氣管、脾臟、食管和胃上皮細胞中表達,在鼠的胃、小腸、結腸和盲腸中也有表達。Hou等研究表明,Caspase-8可以特異性切割GSDMC,將TNF-α誘導的乳腺癌細胞凋亡轉變為細胞焦亡。GSDMD主要在皮膚、食道、胃和免疫細胞中表達,主要用于治療細菌感染。在炎性小體的激活下,Caspase-1會切割高表達的GSDMD,釋放GSDMD-NT誘導巨噬細胞、單核細胞和樹突狀細胞等發生焦亡。另外,胞質脂多糖(LPS)通過激活Caspase-4、5、11切割GSDMD也會誘導焦亡發生。GSDMD成孔釋放危險信號和炎癥因子,募集免疫細胞消除病原體。GSDME常見于癌癥靶向療法,可在大腦、心臟、腎臟和胎盤中檢測出來,一般在正常細胞中表達,而不同癌細胞中由于受其啟動子甲基化表達水平較低或不表達。GSDME可以被Caspase-3切割,誘導細胞焦亡。此外,顆粒酶B(GZMB)在接頭區域切割GSDME也會誘導細胞焦亡。目前文獻報道較多的GSDMs家族蛋白有GSDMB-E如表1所示。GSDMs作為致癌基因或抑癌基因參與癌癥的發生發展,可作為癌癥患者早期篩查、療效評價和臨床預后預測的前瞻性生物標志物,特別是與免疫治療相結合,在腫瘤治療方面具有廣闊的前景。
2.2 激活細胞焦亡的重要途徑
2
.2
.1
經典的細胞焦亡途徑經典細胞焦亡作為最早被研究的焦亡途徑,已經被人們所熟知,由炎性小體激活Caspase-1介導是其主要特征。當細胞受到PAMPs和DAMPs刺激時,PRRs接收危險信號分子,并通過Pyrin結構域(PYD)募集含有CARD的ASC,將炎性體傳感器與Pro-caspase-1橋接,從而激活Caspase-1。活化的Caspase-1將GSDMD切割成N末端和C末端,N末端片段易位到質膜,與質膜上的心磷脂和磷酸肌醇相結合,使質膜出現寡聚化蛋白孔洞,進一步促進炎性因子的釋放,隨之產生細胞腫脹、破裂,最終導致細胞焦亡。激活的Caspase-1也會促進細胞內白細胞介素18(IL-18)、IL-1β前體的成熟,觸發炎癥反應。
2
.2
.2
非經典的細胞焦亡途徑非經典的細胞焦亡途徑與經典細胞焦亡途徑不同,GSDMD的切割是由LPS激活Caspase-4、5(人)和Caspase-11(鼠)來完成。LPS可以直接與 Caspase-4、5、11的CARD結構域結合并使其活化,蛋白水解GSDMD,形成GSDMD-N端p30片段。同時,活化的Caspase-4、5、11可激活細胞膜上的泛連接蛋白-1(Pannexin-1)通道,釋放出單價離子和ATP,激活細胞膜上的P2X7通道,形成孔隙,進而導致細胞腫脹破裂,內容物滲出,IL-1β和IL-18 釋放,誘導細胞焦亡。
表1 GSDMs家族成員在機體內的功能
Table 1 The function of GSDMs family members in the organism
GSDM蛋白家族GSDMs活化酶Activating enzyme 功能Function參與焦亡途徑Pathways involved in pyroptosisGSDMBCaspase-1、3、4、6、7細胞焦亡Caspase-1、3、4、6、7/GSDMB介導的細胞焦亡GZMA顆粒酶A介導的細胞焦亡GSDMCCaspase-8細胞焦亡Caspase-8/GSDMC 介導的細胞焦亡GSDMDCaspase-1焦亡、炎癥、宿主防御經典的細胞焦亡Caspase-4、5、11非經典的細胞焦亡GSDMECaspase-3焦亡、抗腫瘤免疫Caspase-3/GSDME 介導的細胞焦亡GZMB顆粒酶B介導的細胞焦亡
2
.2
.3
Caspase-3/GSDME 介導的細胞焦亡途徑GSDME是一種重要且新穎的線粒體成孔蛋白,可由Caspase-3切割活化,普遍用于腫瘤的治療。當GSDME低表達或者不表達時,會發生細胞凋亡現象;當細胞內GSDME高表達時,Caspase-3能通過切割、活化GSDME蛋白導致膜孔形成,將細胞凋亡轉變為細胞焦亡。GSDME在NCI-H522、SH-SY5Y和SGC-7901等腫瘤細胞系中高表達,在腫瘤治療過程中,化療藥物使GSDME被活化的Caspase-3特異性切割成GSDME-NT和GSDME-CT兩部分,分離出來的GSDME-NT識別并結合膜上的4,5-二磷酸磷脂酰肌醇,造成質膜穿孔,促進乳酸脫氫酶(LDH)和炎性物質的釋放,誘導焦亡的發生。Zhang等研究證明順鉑和紫杉醇均能夠促進Caspase-3切割GSDME,誘導肺癌細胞焦亡。Rogers等研究證明GSDME-NT作用后形成的線粒體孔道能夠釋放細胞色素C,激活凋亡小體,正反饋調節Caspase-3和GSDME的裂解,進一步促進細胞焦亡。
2
.2
.4
顆粒酶介導的細胞焦亡途徑顆粒酶(GZM)是外源性絲氨酸蛋白酶,來自細胞毒性淋巴細胞(CTLs)和自然殺傷細胞(NK)釋放的細胞漿顆粒。CTLs與靶細胞結合時,釋放GZM和穿孔蛋白,穿孔蛋白在細胞膜上聚集,使細胞膜穿孔,為GZM進入靶細胞創造有利條件。已知GZMA/B可引起細胞焦亡,提供了觸發炎性死亡的簡單機制,比經典途徑和非經典途徑簡單得多,是CTLs在抗腫瘤免疫中清除惡性細胞的關鍵機制。其中,活化的CTLs會釋放干擾素,干擾素-g(IFN-g)可以促進GZMA在Lys處將GSDMB切割,NT成孔進而誘導細胞焦亡。GZMB可以在不需要Caspase-3參與的情況下,于D270后直接切割GSDME,誘導細胞焦亡。GZMB也可以通過激活Caspase-3間接誘導GSDME依賴性細胞焦亡。因此,GZMA/B對抗腫瘤免疫的研究具有重要意義。細胞焦亡重要途徑如圖1所示。
3 細胞焦亡對腫瘤免疫微環境的影響
腫瘤免疫微環境(TIME),是指在腫瘤微環境中,與T細胞、腫瘤相關巨噬細胞(TAMs)、髓源性抑制細胞(MDSCs)、NK細胞等免疫細胞相關的微環境。TIME可以分為三種類型,分別是浸潤排斥的腫瘤免疫微環境(I-E TIME)、炎性浸潤的腫瘤免疫微環境(I-I TIME)和三級淋巴樣結構的腫瘤免疫微環境(TLS-TIME)。TIME是腫瘤微環境的重要組成部分,大多數腫瘤的形成過程都經歷了由可控性炎性反應到不可控炎性反應的轉變,從而產生適合其生長的微環境,促進腫瘤的增殖、侵襲、轉移、血管生成及免疫逃逸。在TIME中,腫瘤細胞可以借助TIME來逃避免疫監視。通過誘導細胞焦亡可以將腫瘤微環境改變為免疫刺激狀態,對于腫瘤的免疫治療具有重要研究價值。GSDM在TIME中的作用機制如圖2所示。

1)炎性小體識別病原相關分子模式(PAMPs)和損傷相關分子模式(DAMPs),由包括NLR、AIM2或Pyrin在內的傳感器來招募含有CARD的凋亡相關斑點樣蛋白(ASC),上調Caspase-1,激活GSDMD,釋放IL-1β/18。2)脂多糖(LPS)作用于Caspase-4、5、11,激活GSDMD,促進IL-1β/18的成熟和釋放。3)化療藥物誘導Caspase-3將GSDME切割成GSDME-NT,并在細胞膜上打孔,將凋亡轉化為焦亡。4)細胞毒性淋巴細胞釋放GZMA/B,GZMA在Lys244處將GSDMB切割,N末端成孔進而誘導細胞焦亡。GZMB可以在D270后代替Caspase-3直接切割GSDME誘導細胞焦亡,或者通過激活Caspase-3間接誘導GSDME依賴性細胞焦亡。 1) Inflammasomes recognize pathogen-associated molecular patterns (PAMPs) and damage associated molecular pattern molecules (DAMPs). Sensors including NLR, AIM2 or pyrin recruit apoptosis-associated speck-like protein containing a CARD (ASC), up-regulate caspase-1, activate GSDMD and release IL-1β/18. 2) Lipopolysaccharide (LPS) acts on caspase-4,5 and 11 to activate GSDMD and promote the maturation and release of IL-1β/18.3) Chemotherapeutic drugs induce caspase-3 to cleave GSDME into the N-terminal and perforate the cell membrane to transform cell apoptosis into pyroptosis. 4) Cytotoxic lymphocytes release GZMA/B. GZMA cleaves GSDMB at Lys244, and N-terminal form spores and induces pyroptosis. GZMB can replace caspase-3 after D270 to directly cleave GSDME to induce pyroptosis, or indirectly induce GSDME-dependent pyrolysis by activating caspase-3.圖1 4種主要焦亡途徑示意圖Fig.1 Schematic diagram of four main pyroptosis pathways
3.1 細胞焦亡對TIME中T細胞的影響
使用化療藥物誘導GSDME激活細胞焦亡,可以增強抗腫瘤免疫。TIME中的CD8+T淋巴細胞僅有較少的穿孔素,以至于其無法長時間穿透靶細胞。腫瘤中的GSDME可以避開CD8+T淋巴細胞,在遇到靶細胞時釋放GSDME-NT結構域,插入細胞膜的內小葉并形成廣泛的孔,將CD8+T細胞的毒性顆粒釋放到靶細胞中,進而增加CD8+T淋巴細胞的抗腫瘤功能,將腫瘤細胞殺滅。隨著毒性顆粒進入腫瘤細胞,GZMA/B直接或間接誘導腫瘤靶標中的GSDM依賴性細胞焦亡,進而形成一種免疫激活狀態。可以將TIME分為T細胞缺失或排斥的“冷腫瘤”環境和T細胞浸潤、免疫激活的“熱腫瘤”環境。促炎細胞因子通過細胞焦亡成孔釋放到TIME中,招募更多T細胞浸潤,增強細胞毒性T細胞的免疫作用,使TIME由“冷”轉“熱”,導致激活狀態的TIME能夠有效地殺滅腫瘤細胞。然而,在嵌合抗原受體T(CAR-T)/GSDME免疫療法介導靶腫瘤細胞焦亡的過程中,高遷移率族蛋白1(HMGB1)促進巨噬細胞中Caspase-1和GSDMD的激活,釋放大量的IL-6和IL-1β,產生細胞因子釋放綜合征(CRS),降低T細胞療法的有效性。因此,尋找最佳的細胞焦亡程度對TIME治療是至關重要的。
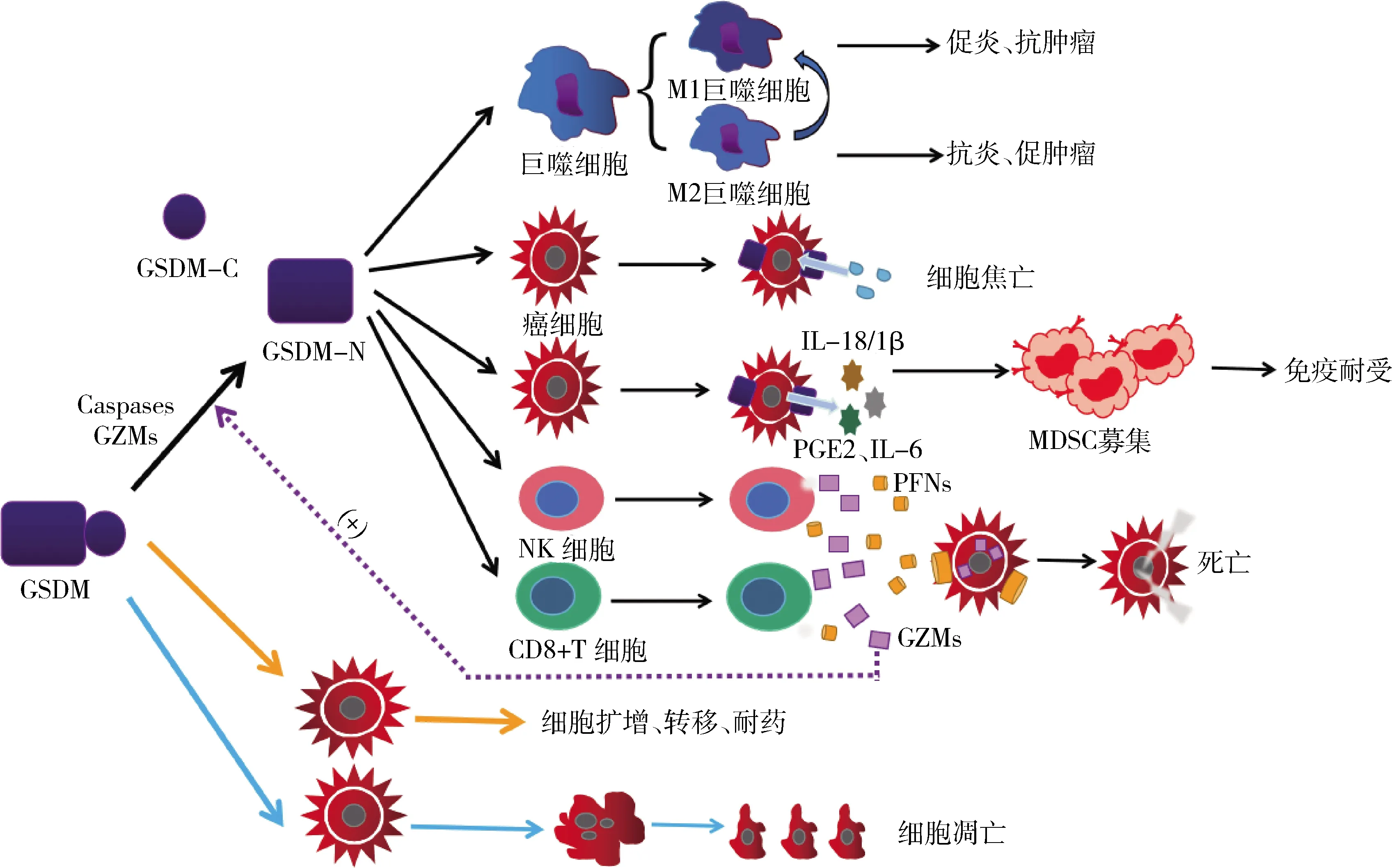
GSDM可以影響癌細胞的增殖、凋亡、侵襲、轉移和耐藥性。GSDM可以被Caspases和GZMs等蛋白酶切割,釋放GSDM-NT。GSDM-NT能在細胞膜上打孔,誘導細胞焦亡,并作用于巨噬細胞、NK細胞、CD8+T細胞等免疫細胞協調抗癌免疫。GSDM-NT增強CD8+T細胞和NK細胞的浸潤能力,并促進M2巨噬細胞向M1巨噬細胞轉變。細胞焦亡產生大量的炎性因子募集MDSC,造成免疫耐受。 GSDM can affect the proliferation, apoptosis, invasion, metastasis and drug resistance of cancer cells.GSDM can be cleaved by proteases such as Caspases and GZMs to release GSDM-NT. GSDM-NT can punch holes in the cell membrane, induce pyroptosis and act on immune cells such as macrophages, NK cells, and CD8+T cells to coordinate anti-cancer immunity. GSDM-NT enhances the infiltration capacity of CD8+T cells and NK cells, and promotes the conversion of M2 macrophages to M1 macrophages. Pyroptosis produces a large number of inflammatory factors to recruit MDSC, resulting in immune tolerance.圖2 GSDM在TIME中的作用機制Fig.2 The mechanism of GSDM in TIME
3.2 細胞焦亡對TIME中巨噬細胞的影響
TIME中含氧量較少,刺激嗜中性粒細胞釋放各種趨化因子在炎癥部位激活并募集巨噬細胞,這些巨噬細胞稱為TAMs,占某些實體瘤的50%左右。TAMs分為M1和M2兩種,M1與M2會以相反的方式影響免疫反應,既可以介導吞噬和氧化等抗腫瘤作用又可以為惡性細胞提供營養,兩者可相互轉化。在正常情況下,巨噬細胞能發揮其吞噬作用將病原體吞噬,隨著巨噬細胞的生長,Caspase-3會切割GSDME產生GSDME-NT片段,靶向質膜以誘導細胞焦亡。但由于巨噬細胞的壽命較長,很多腫瘤細胞在巨噬細胞死亡前進行快速復制,將其變成為更易感染的靶標,就算是中性粒細胞也無法阻止巨噬細胞中病原體的復制,從而產生更多的病原體并釋放出來。而當被炎性體激活或使用藥物誘導細胞焦亡時,可以使巨噬細胞提前溶解,快速釋放病原體,在其還沒有復制時就暴露在清除機制中,被免疫細胞消滅。在TIME這種缺氧狀態中,細胞程序性死亡配體1(PD-L1)可以與巨噬細胞產生的TNF-α相互作用,誘導Caspase-8激活GSDMC,進而促進細胞焦亡,使巨噬細胞產生誘導腫瘤細胞壞死的功能。Tan等研究表明,使用D2多巴胺受體可調節TIME,上調M1并限制NF-κB 信號通路,觸發GSDME介導的乳腺癌細胞焦亡。因此,在臨床治療過程中,巧妙的運用巨噬細胞的免疫特性并聯合用藥,為實現靶向治療腫瘤提供了新思路。
3.3 焦亡對TIME中髓源性抑制細胞的影響
MDSC是粒細胞、巨噬細胞和樹突狀細胞的前體,在病理條件下MDSC抑制免疫反應,促進腫瘤的生長、侵襲。在一些TIME中,由DAMPs觸發的炎性小體NLRP3的激活促進NF-κB轉錄因子的易位,上調Pro-IL-18和Pro-IL-1β表達,Caspase-1介導細胞焦亡將腫瘤激活為免疫狀態,釋放IL-18和IL-1β等炎性因子,激活前列腺素E2(PGE2)回路誘導IL-6生成,募集MDSC,如S100A8/A9 誘導血清淀粉樣蛋白A3直接在肺中募集MDSC并促進轉移,HMGB1在腹部手術后大量釋放,招募大量MDSC促進結腸癌腹膜轉移,可見,過度的細胞焦亡產生大量炎性因子會促進MDSC的募集,導致免疫耐受。然而,Erkes等使用BRAF抑制劑和MEK抑制劑(BRAFi+MEKi)體內聯合泊苷治療中發現,通過BRAFi+MEKi處理,可誘導Caspase-3活化并切割GSDME促進細胞焦亡,在上調CD4+T和CD8+T細胞的同時,降低了MDSC的含量,減弱腫瘤的耐藥性,使腫瘤細胞無處可逃。
3.4 細胞焦亡對TIME中NK細胞的影響
NK細胞是一種天然免疫細胞,可以殺傷腫瘤細胞。在癌細胞中,GSDME高表達顯著增加了腫瘤內NK細胞數量,并促進NK細胞殺死腫瘤細胞,為持續的炎癥微環境提供各種活性氮、活性氧、活性醛類、細胞因子、趨化因子和生長因子,將腫瘤微環境激活為免疫刺激狀態,使細胞內促炎因子從焦亡細胞中快速有效地釋放出來,從而引發腫瘤的炎癥。同時,NK細胞對GSDME引導的焦亡也具有促進作用。在TIME中,腫瘤會逃避NK細胞所產生的凋亡,在凋亡信號通路受阻時,NK細胞常聯合細胞毒性T細胞與靶細胞相互作用,產生特異性殺傷功能,并釋放GZMB,代替或激活Caspase-3在D270位點后激活GSDME,促進腫瘤細胞焦亡,使毒性顆粒進入腫瘤細胞,發揮NK細胞的自然殺傷作用,抑制腫瘤生長。因此,細胞焦亡和NK細胞在TIME中的功能是互相促進的,在抗癌免疫中形成正反饋回路,將腫瘤的免疫微環境從“冷”改變為“熱”。
4 展 望
細胞焦亡在腫瘤中的作用是復雜的,是一把雙刃劍,影響到腫瘤發生的所有階段。一方面,炎癥小體介導的細胞焦亡和焦亡釋放的細胞因子改變了腫瘤免疫微環境,且通過逃避免疫監視促進了腫瘤的發展。另一方面,細胞焦亡產生的細胞因子可以募集免疫細胞并激活免疫系統,從而提高腫瘤免疫治療的效率。為了開發靶向細胞焦亡的腫瘤免疫療法,需要進一步探索細胞焦亡對免疫系統的潛在功能。未來將對以下問題深入研究,①探索焦亡發揮抗腫瘤作用的其他途徑及分子機制;②研究原發腫瘤發展中誘導細胞焦亡激活的因素,從而探尋避免腫瘤發生的策略;③明確細胞焦亡對免疫細胞的調節和免疫反應的功能,提高癌癥治療的效率以及探索更多的抗腫瘤療法。
此外,細胞焦亡是由GSDMs介導的程序性細胞死亡,其在激活免疫系統方面的最新進展越來越被關注。因為GSDMs 在細胞焦亡和癌癥進展中的重要功能,所以藥物對其蛋白活性的正負調節成為焦亡發生后對TIME影響的關鍵。臨床中化療藥物的副作用常限制了這些藥物的使用劑量,而GSDME是抑瘤因子同時也是化療副作用的關鍵分子。藥物誘導不同組織細胞焦亡的廣泛研究及GSDMs蛋白家族的每位成員在TIME中扮演的不同角色的深入研究將為動物癌癥早期篩查、療效評價、臨床預后預測及尋找安全有效抗癌藥物提供科學依據。
