小飛鼠微衛星位點的篩選及遺傳特征分析
2022-05-16 10:26:20田新民楊孟平宋雅祺劉小慧廉明棟
野生動物學報 2022年2期
關鍵詞:物種
田新民 楊孟平 宋雅祺 劉小慧 廉明棟 陳 紅
(牡丹江師范學院生命科學與技術學院,牡丹江,157011)
小飛鼠(Pteromysvolans)隸屬哺乳綱(Mammalia)嚙齒目(Rodentia)鼯鼠科(Pteromyidae)飛鼠屬,為樹棲夜行滑行類物種,主要分布于歐亞大陸北部,國內主要分布在東北、華北、西北等北方林區[1-3]。由于不合理的森林采伐,森林生境趨于破碎化甚至喪失,已導致全球許多地區的小飛鼠種群處于瀕危狀態[4-5]。作為森林可持續經營的重要指示和傘護物種[6],我國也將其列為“易危”與“三有”重點保護的野生動物[7-8]。目前,國外在小飛鼠巢址選擇、家域、擴散、遺傳多樣性和分子系統進化等領域已開展系列研究[9-13],為該物種的科學保護提供了參考,而我國對小飛鼠生態學領域的研究基本空白。
保護遺傳學研究在揭示瀕危物種遺傳適應與瀕危機制、實現物種的精準保護與管理方面發揮了重要的作用,微衛星分子標記以選擇中性、多態性高及共顯性等優點,在保護遺傳學領域有廣泛應用[14]。由于微衛星側翼序列在物種間的高度保守性,表現出近緣物種間的引物通用性,從而可實現從近緣物種微衛星中篩選出目標物種的位點,實現該物種的群體遺傳學研究[15-16],相比其他篩選策略,這是一種低成本與高效率的方法。本研究對小飛鼠其他亞種、近緣物種的微衛星位點進行測試篩選,構建小飛鼠東北亞種(P.v.arsenjevi)的微衛星分析系統,藉以為該亞種的保護遺傳學領域深入研究提供參考。
1 材料與方法
1.1 樣本與DNA提取
在伊春市五營區龍江松鼠養殖場共收集30只自然死亡的小飛鼠,此樣本均為東北亞種野外個體的繁殖后代。從這些個體的腿部取肌肉組織,采用血液/細胞/組織基因組DNA試劑盒(天根,北京)提取基因組DNA。用ND-1000紫外分光光度計(Nanodrop,USA)測定DNA濃度,稀釋至10 ng/μL,4 ℃存放備用。
1.2 PCR擴增
根據GenBank中已報道的微衛星位點,結合文獻[17-22],選取鼯鼠科小飛鼠指名亞種(P.v.volans)、北美飛鼠(Glaucomyssabrinus)、海南小飛鼠(Hylopetesphayrei)和赤頰林飛鼠(H.sagitta),以及松鼠科(Sciuridae)北松鼠(Sciurusvulgaris)5個物種中擴增效果較好的34對引物(表1),以小飛鼠東北亞種的DNA為模板進行擴增。采用不同退火梯度,對微衛星位點初步篩選;成功擴增的位點,上游引物5′端進行熒光標記(Fam、Hex、Tamra或Rox)。擴增體系20.0 μL:5 U/μL ExTaqDNA聚合酶(TaKaRa,Japan)0.1 μL,10×Buffer 2 μL,2.5 mmol/L dNTPs 1.6 μL,10 μmol/L上下游引物各0.5 μL,模板DNA 1.5 μL,超純水13.8 μL。PCR反應條件均為:94 ℃,4 min;(94 ℃,30 s;50~59 ℃,30 s;72 ℃,30 s)× 35 個循環;72 ℃,10 min。擴增產物經過2.0%瓊脂糖凝膠電泳檢測,其中熒光標記引物成功擴增的PCR產物用ABI 3730XL測序儀(Applied Biosystems Inc.,America)進行毛細管電泳檢測,判讀等位基因大小。所有的引物合成和基因分型均由上海生工完成。

表1 鼯鼠科和松鼠科物種的微衛星位點信息
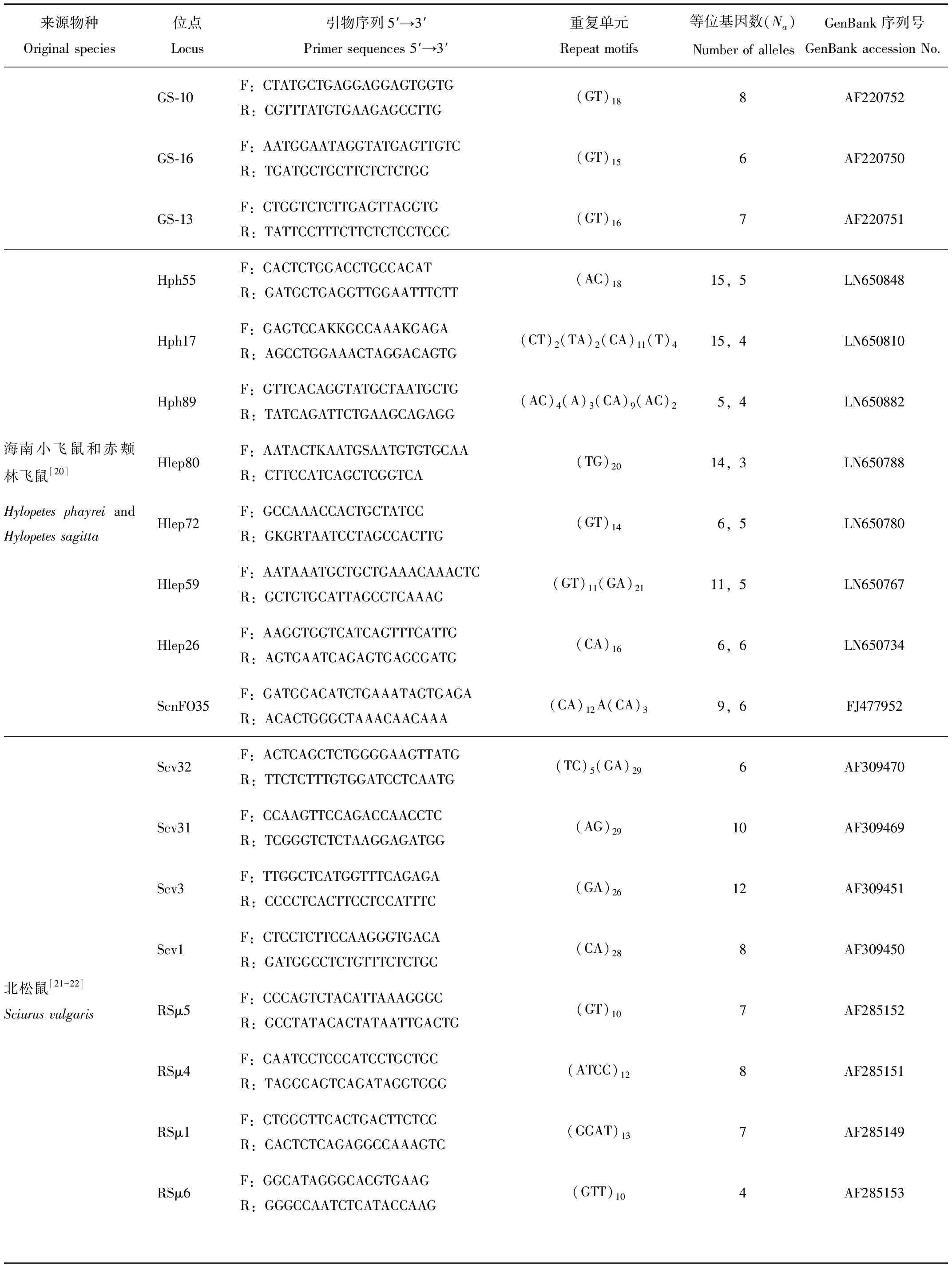
續表1
1.3 數據處理
采用軟件GENALEX 6[23]計算各位點的等位基因數、觀測雜合度和期望雜合度;Excel microsatellite tool kit[24]計算各位點的多態信息含量(polymorphic information content,PIC)。采用軟件GENEPOP 4.0[25]測算各位點是否符合Hardy-Weinberg平衡,位點間的連鎖不平衡情況和概率檢驗使用馬爾可夫鏈法(Markov chain method),參數設置:dememorization=10 000;batch=20;iteration=5 000,以Bonferroni法對檢驗的顯著性進行修正。對于多態性位點,采用軟件GIMLET 1.3.3[26]計算位點的個體聯合判別率(probability of identity,PID),PID指無親緣關系或同胞個體之間具有相同基因型的概率。
2 結果與分析
PCR產物經瓊脂糖凝膠電泳檢測發現,共有14個位點有明確的符合微衛星片段大小的擴增產物,占篩選位點總數的41.2%。毛細管電泳(表2)顯示,上述14個位點均能進行準確的基因型判斷,但是位點Hlep26和GLSA48只有1個等位基因。其余12個位點Pvol10、Pvol41、PvolE1、PvolE5、PvolE6、PvolE10、Scv3、Hlep59、Hlep72、Hlep80、Hph17和ScnFO35的等位基因8~16個,多態信息含量為0.757~0.902,均為高度多態性位點,占可擴增位點的85.7%(12/14)。
12個高度多態性位點的觀測雜合度為0.500~0.950,期望雜合度為0.775~0.909,其中,有6個位點(Pvol41、PvolE6、Hlep59、Hlep72、Hph17和ScnFO35)符合Hardy-Weinberg平衡(P=0.123~0.998);其余6個位點(Pvol10、PvolE1、PvolE5、PvolE10、Scv3和Hlep80)都偏離Hardy-Weinberg平衡(P<0.05),皆呈現不同程度的雜合度不足(表2)。在12個位點中,均未檢測到連鎖不平衡現象。Gimlet分析顯示,12個微衛星位點的個體聯合判別率很高,2只個體具有相同基因型的無偏概率為3.49×10-22;即使全同胞個體,錯誤判別概率Prod(sibs)也只有1.44×10-6。多態性最高的前6個位點(Hlep59、PvolE1、PvolE5、Pvol10、Pvol41和Hlep72)擴增失敗時,全同胞錯誤判別概率也只上升到0.16%(圖1)。

表2 14個微衛星位點在小飛鼠種群中的遺傳特征

圖1 12個微衛星位點按照個體聯合判別率順序的個體基因型相似概率曲線Fig.1 Probability of identity(PID)curve generated 12 microsatellite loci
3 討論
微衛星側翼序列的保守性,是近緣物種間位點交叉擴增的基礎,為許多物種的遺傳管理提供了極大的便利[14-16,27-29]。本研究選擇了鼯鼠科與松鼠科中5個物種(小飛鼠指名亞種、北美飛鼠、海南小飛鼠、赤頰林飛鼠和北松鼠)內高多態性的34個微衛星位點,在小飛鼠東北亞種內共獲得12個具有多態性的位點,成功率為35.3%,其中,同物種小飛鼠指名亞種的位點在東北亞種內多態性擴增成功率最高,為85.7%(6/7);同為鼯鼠科不同屬的海南小飛鼠和赤頰林飛鼠的成功率次之,為75.0%(7/8);松鼠科的北松鼠最低,為12.5%(1/8)。由此可以看出,近緣種基因組序列相似度高,交叉擴增成功概率高,而遠源種之間情況呈相反的趨勢[15]。例外的是,在鼯鼠科的北美飛鼠11個微衛星位點內,僅有1個位點能夠擴增成功,并無多態性。
本研究檢測到12個多態性微衛星位點,其中有6個位點偏離Hardy-Weinberg平衡。相關研究認為,導致位點和群體偏離Hardy-Weinberg平衡的原因有很多,如種群亞結構、近親交配、遷入和遷出等[29-30]。在這6個位點中,均呈現一定程度的雜合度不足。本研究樣本來自籠養的繁殖個體,可能受到近親交配的影響,導致了位點偏離Hardy-Weinberg平衡。得到的12個位點均為高度多態性位點(PIC值>0.5),其個體聯合判別率很高;僅使用相比多態性略低的6個位點(PvolE6、PvolE10、Scv3、Hlep80、Hph17和ScnFO35),全同胞錯誤判別概率也小于1%(0.16%)。因此,本研究篩選的12個多態性位點,可作為非損傷性遺傳取樣個體識別的參考位點,也可為小飛鼠東北亞種群體遺傳學研究與管理提供重要的分子標記。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
