甘薯近緣二倍體野生種TPS家族全基因組鑒定及表達分析
2022-05-17 07:46:26賈小云王文斌
山西農業科學 2022年5期
關鍵詞:分析
梁 璇,李 鵬,楊 哲,賈小云,王文斌
(山西農業大學 生命科學學院,山西 太谷 030801)
甘 薯(Ipomoea batatas(L.)Lam)屬 旋 花 科(Convolvuaceae)番薯屬(Ipomoea)雙子葉植物,是一種典型的短日照作物,喜光耐旱且根系發達,但對寒冷的耐受性較低。甘薯具有高產、適應性廣、抗逆性強、利用價值高等特點,是世界上種植最廣泛的作物之一,2019年全球甘薯產量為9 200萬t,以我國為首,占世界總量的56%。自1950年以來,人們一直在研究番薯屬植物化學,一些番薯屬物種顯示出抗菌、鎮痛、解痙、降壓和抗癌活性[1]。甘薯憑借其食用價值、藥用價值、飼養價值和經濟價值成為我國重要的經濟作物,在我國不同區域均有廣泛種植,但其產量受到干旱、高溫、鹽堿等環境因素的嚴重影響,因此,研究甘薯抗逆相關基因,對于降低非生物脅迫對甘薯產量的影響、提高甘薯產量都有著重要作用,同時可為甘薯的分子遺傳改良提供優質的種質資源。
海藻糖(Trehalose)是天然雙糖中最穩定且具有非還原性的糖類,通常存在于真菌、細菌和無脊椎動物中,可取代水分子與生物膜及蛋白質等生物大分子的羥基連接以維持其結構與功能的穩定,在抗逆反應中對生物體有至關重要的保護作用[2]。海藻糖-6-磷酸合成酶(Trehalose-6-phosphate synthase,TPS)作為植物海藻糖合成途徑的關鍵酶,在植物受到脅迫時表達增強,使海藻糖含量升高從而提高植物體的抗逆性。海藻糖在植物體內的合成是基于UDP-葡萄糖和6-磷酸葡萄糖在TPS的催化下合成6-磷酸海藻糖(Trehalose-6-phosphate,Tre6P),再由海藻糖-6-磷酸酯酶(Trehalose-6-phosphate phosphate,TPP)催化合成海藻糖[3]。但由于細胞內存在許多可以將Tre6P脫磷酸化為海藻糖的非特異性磷酸酯酶,可以不經過TPP直接催化Tre6P脫磷酸化生成海藻糖,由此可見,指導編碼TPS蛋白的TPS基因能否被成功轉錄表達對植物體能否合成海藻糖有決定性作用[4-5]。
近年來,已有許多研究證實,TPS基因在不同植物如擬南芥[6]、番茄[7]、水稻[8]等體內的過表達會增強植物抵御非生物脅迫的能力,但到目前為止,僅對甘薯IbTPS1基因進行了生物信息學分析并驗證了其在酵母細胞中的異源過表達會提高酵母細胞的抗逆能力[2,9],而甘薯近緣野生種TPS基因的研究分析還未見報道。但是栽培甘薯是六倍體物種,不僅基因組龐大,結構復雜,且具有無性繁殖、自交不親和等特性,使得對甘薯的遺傳組成和基因的研究進展緩慢,遠落后于其他作物[10]。有研究表明,甘薯的二倍體近緣野生種三淺裂野牽牛(Ipomoea trifida)和三裂葉薯(Ipomoea triloba)的基因組測序均已完成[11-12]。
本研究通過對甘薯的二倍體近緣野生種——三淺裂野牽牛和三裂葉薯的TPS家族全基因組進行分析,以探討TPS基因在植物生長發育及非生物脅迫中功能及應用,為六倍體甘薯TPS功能的進一步研究鑒定奠定基礎,同時也為甘薯的遺傳改良提供優良的候選基因。
1 材料和方法
1.1 三淺裂野牽牛ItfTPS和三裂葉薯Itb TPS家族成員的鑒定及系統進化分析
在密歇根州立大學甘薯資源庫(https://sweetpotato.plantbiology.msu.edu/)中獲得三淺裂野牽牛和三裂葉薯的全基因組序列和注釋文件,并建立本地數據庫。用TBtools[13]從Gtf/Gff3文件中提取基因結構的注釋信息,基于基因結構注釋信息從基因組中提取出基因的CDS序列,簡化后批量翻譯為蛋白序列。在Pfam數據庫(https://pfam.janelia.org/)中下載GT 20_TPS保守結構域的隱馬爾可夫模型(hidden markov model,HMM)PF00982,同時使用HMMER3.0[14]軟件從三淺裂野牽牛和三裂葉薯的蛋白序列中進行檢索,將檢索出的蛋白序列用在線網站NCBI CDD(https://www.ncbi.nlm.nih.gov/cdd/)預測其結構域,排除特征匹配度低及重復的序列,最終確定ItfTPS和ItbTPS家族成員。在NCBI(https://www.ncbi.nlm.nih.gov/)中下載擬南芥及水稻的TPS蛋白序列,使用MEGA 7.0[15]對ItfTPS、ItbTPS、AtTPS及OsTPS的家族成員進行系統進化分析,采用鄰接法構建系統進化樹,校驗參數Bootstrap=1 000,其余參數為默認值。使用Evolview[16](http://evolgeniuus.info//evolview-v2/#login)將獲得的nwk文件進行美化。
1.2 Itf TPS和Itb TPS家族蛋白理化性質分析
使用在線網站Ex PASy[17](http://web.expasy.org/protparam/)預測ItfTPS和ItbTPS基因家族成員編碼蛋白質的氨基酸長度、相對分子量、理論等電點等理化性質。使用在線網站PSORT(https://psort/.hgc.jp/form.html)預測蛋白的亞細胞定位。
1.3 Itf TPS和ItbTPS基因家族成員motif和基因結構分析
使用在線網站MEME[18](https://meme-suite.org/tools/meme)預測ItfTPS和ItbTPS基因家族成員的保守基序,并利用TBtools對其基因結構和保守基序進行可視化。
1.4 Itf TPS和Itb TPS家族成員保守結構域分析
通過生物結構序列NCBI-CDD和Pfam工具對ItfTPS和ItbTPS基因家族成員進行結構域的保守型分析,并利用TBtools將其可視化。
1.5 Itf TPS和Itb TPS基因家族染色體定位
利用密歇根州立大學甘薯資源庫中下載的三淺裂葉牽牛和三裂葉薯的Gff3文件,用TBtools對篩選到的ItfTPS和ItbTPS基因家族成員進行位置信息的提取并使其可視化。
1.6 Itf TPS和Itb TPS基因家族成員的組織表達分析及不同脅迫下的表達模式分析
在密歇根州立大學甘薯資源庫中下載三淺裂野牽牛和三裂葉薯的組織表達轉錄組數據和不同脅迫下的轉錄組數據,分別對ItfTPS和ItbTPS家族成員在花、花蕾、葉、根和莖中的表達量及其組培幼苗全株在不同脅迫下的表達量進行以2為底的對數處理,用TBtools繪制ItfTPS和ItbTPS基因家族成員組織表達熱圖和不同脅迫處理下的表達熱圖。
2 結果與分析
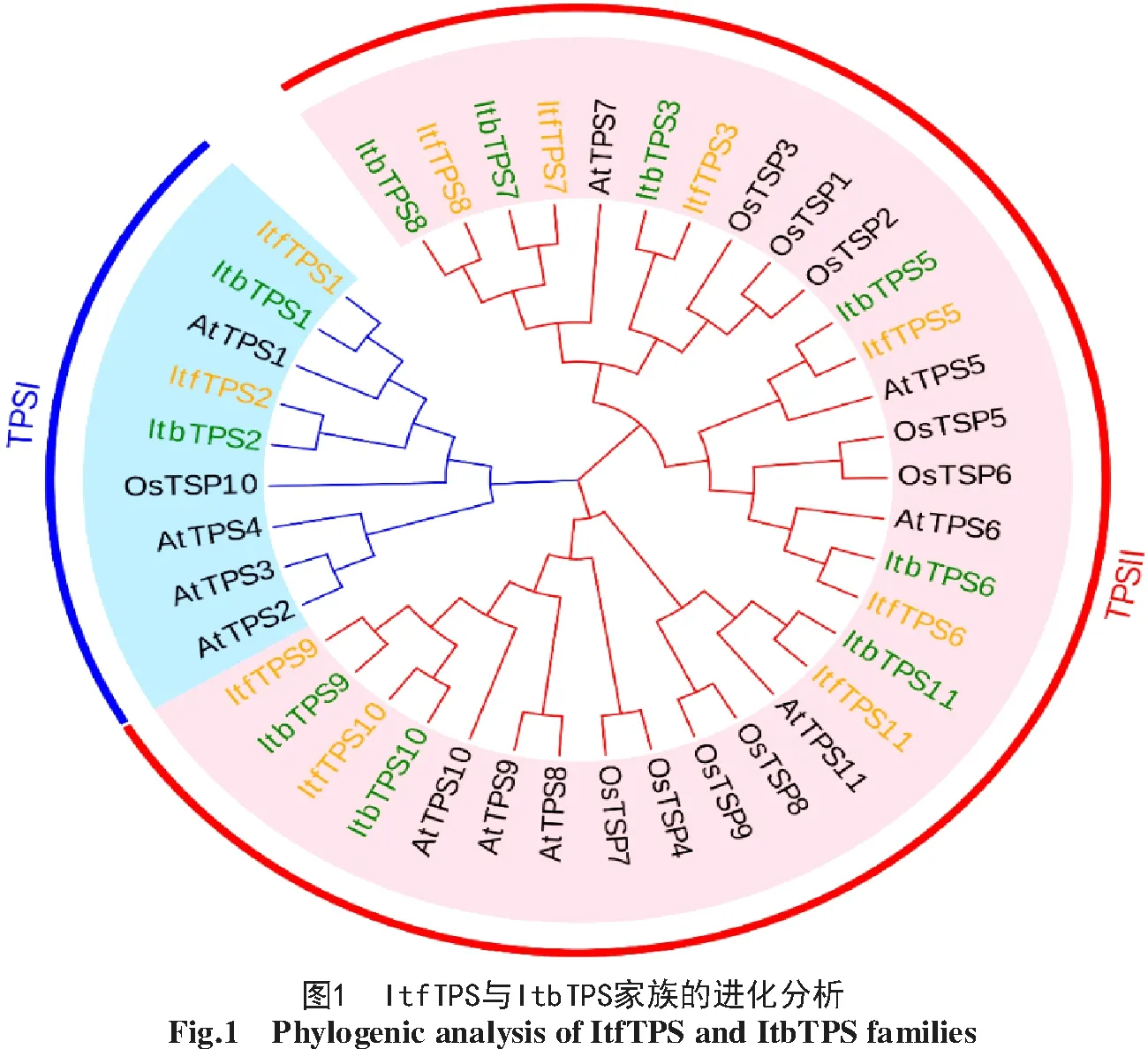
2.1 ItfTPS和ItbTPS家族成員的鑒定及系統進化分析
通過隱馬爾可夫模型,在2個甘薯近緣野生種中分別鑒定到10個Itf/ItbTPS,結合11個AtTPS和9個OsTPS構建系統進化樹,并根據其與AtTPS的同源關系進行命名,結果如圖1所示,ItfTPS與ItbTPS高度同源,Itf/ItbTPS均分為2個亞族TPSⅠ與TPSⅡ,其中TPSI包含Itf/ItbTPS1和Itf/ItbTPS2,與AtTPS1同源性較為接近;TPSⅡ包含8個Itf/ItbTPS,除Itf/ItbTPS3與OsTPS1~OsTPS3高度同源外,Itf/ItbTPS與擬南芥的親緣關系更近。
2.2 Itf TPS和Itb TPS家族蛋白理化性質分析
對ItfTPS和ItbTPS蛋白進行理化性質與亞細胞定位預測結果顯示(表1),Itf/ItbTPS基因編碼氨基酸長度在842~940個;分子質量跨度較大,在95 774.49~105 877.07 u;等 電 點 范 圍 在5.46~6.69。亞細胞定位顯示,ItfTPS有4個定位于細胞核,4個定位于細胞質,2個定位于線粒體基質;ItbTPS有5個定位于細胞核,4個定位于細胞質,1個定位于線粒體基質。
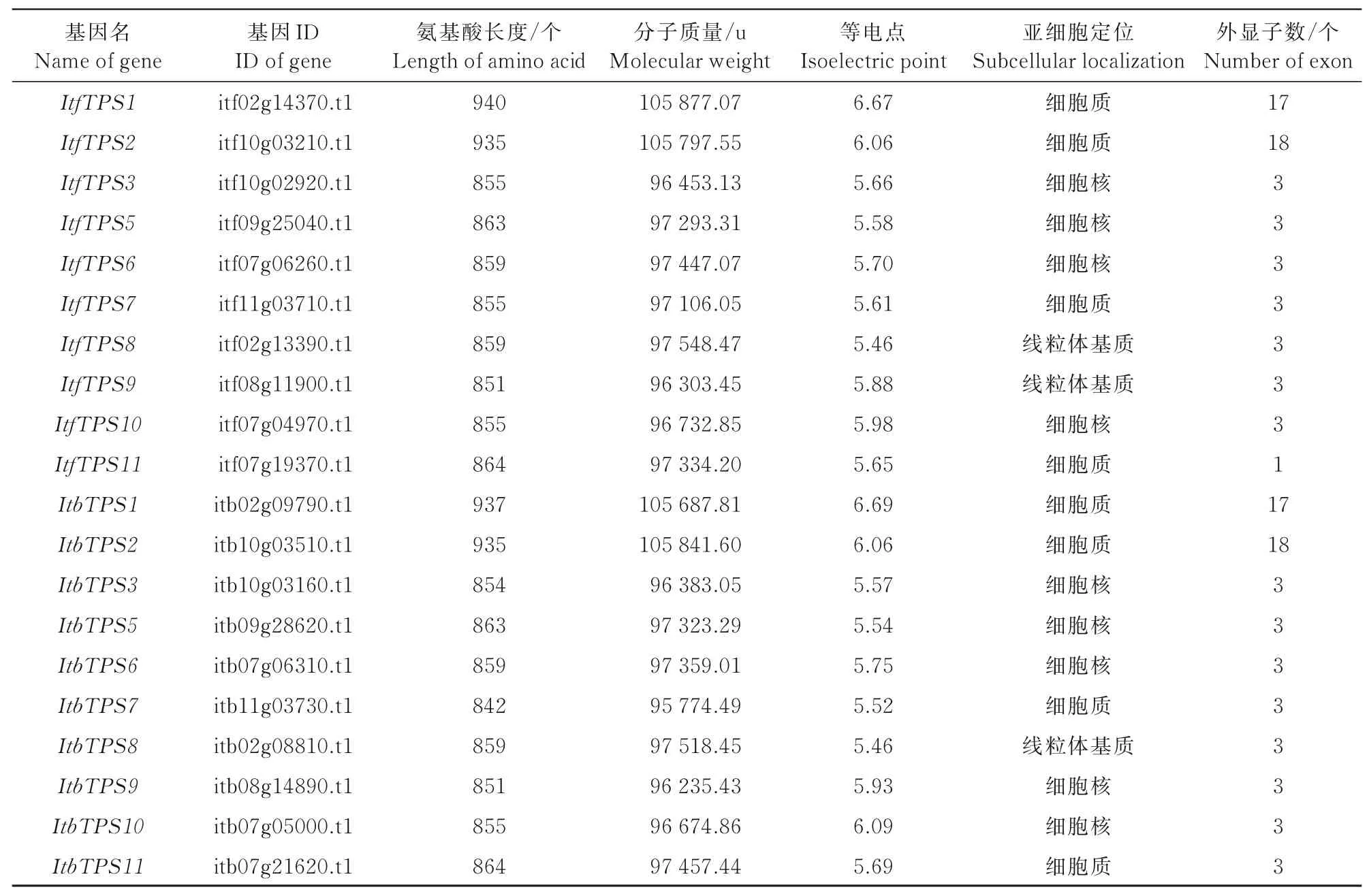
表1 ItfTPS和Itb TPS蛋白理化性質分析Tab.1 Analysis of physical and chemical properties of Itf TPSand Itb TPSproteins
2.3 Itf TPS和Itb TPS基因家族成員motif和基因結構分析
由TBtools軟件繪制的ItfTPS和ItbTPS家族motif與基因結構分析結果顯示(圖2),TPSⅠ中Itf/ItbTPS1包含17個外顯子,7個motif(motif1~motif7);Itf/ItbTPS2包含18個外顯子,8個motif(motif1~motif7,motif3);TPSⅡ中,除ItfTPS11只含有1個外顯子外,其余均含有3個外顯子,且Itf/ItbTPS均含有motif1~motif10。這些結果表明,TPS基因在進化過程中motif基序和基因結構的保守性較強,且TPSⅠ和TPSⅡ之間存在高度分化。

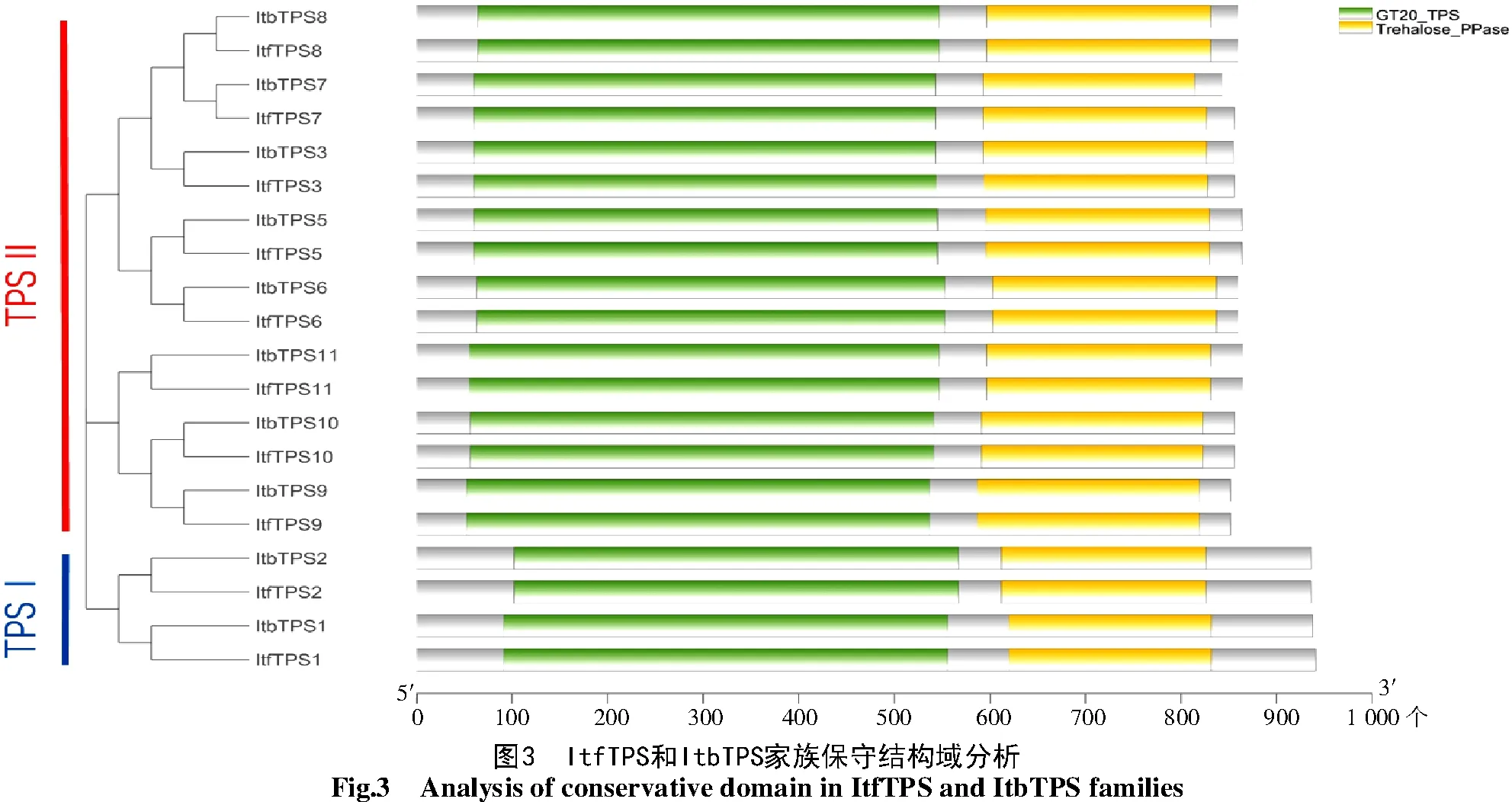
2.4 ItfTPS和ItbTPS家族成員保守結構域分析
對ItfTPS和ItbTPS家族成員的結構域進行分析可知(圖3),ItfTPS和ItbTPS家族均為雙結構域,分別為糖基家族轉移酶(Glyco_transf_20,TPS;PF00982)和海藻糖磷酸酶(Trehalose_PPase,TPP;PF02358)。TPS結構域位于N端,TPP結構域位于C端,二者在TPS家族成員的相對位置較為相似,說明TPS基因在進化過程中結構域的保守性較強,但TPSⅠ和TPSⅡ之間存在高度分化。
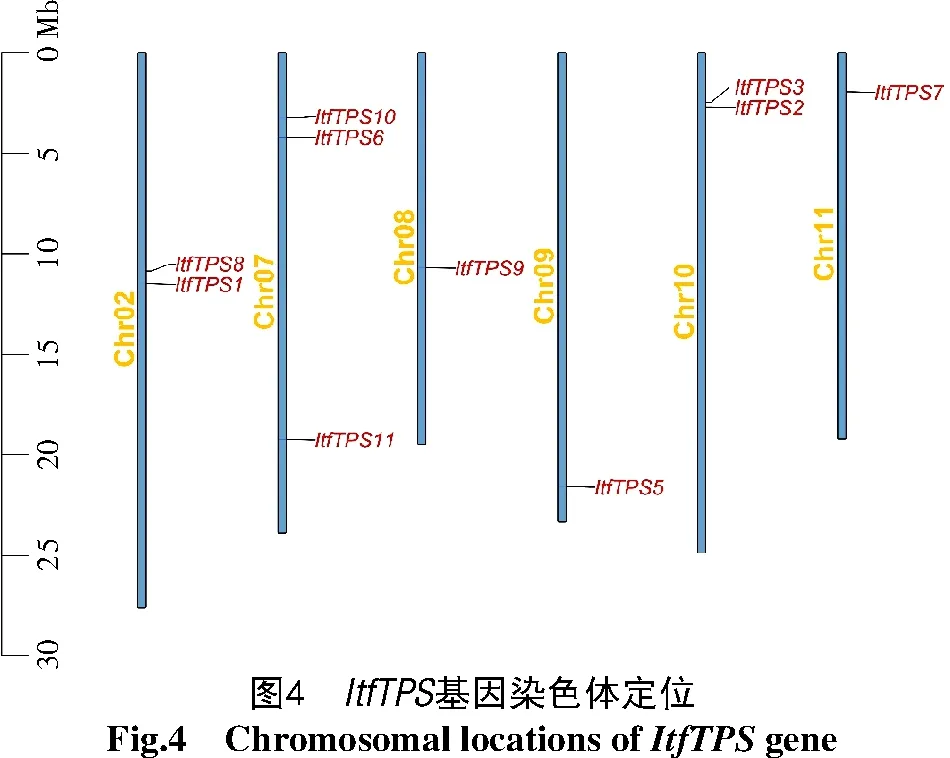
2.5 ItfTPS和Itb TPS基因家族染色體定位
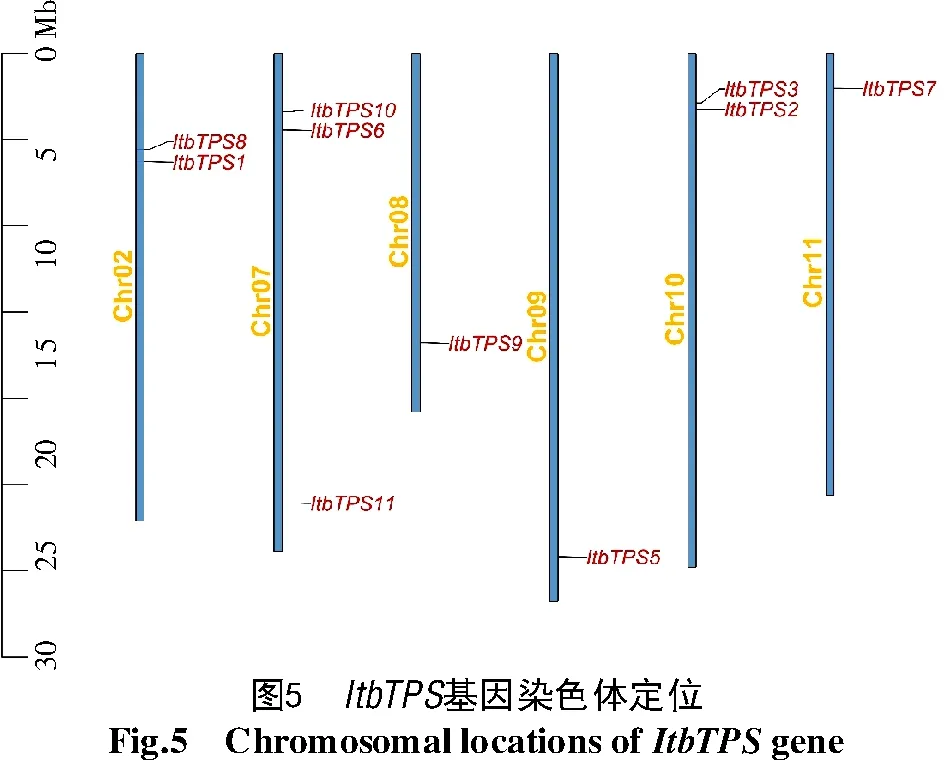
利用Tbtools繪制ItfTPS(圖4)和ItbTPS(圖5)基因家族在染色體中的分布情況,結果發現,10個Itf/ItbTPS位于2號及7~11號染色體上,且分布位置較為相似,除了ItfTPS1和ItfTPS8位于2號染色體中部而ItbTPS1和ItbTPS8位于2號染色體上部以及ItfTPS9位于8號染色體中部而ItbTPS9位于8號染色體上部外,其他Itf/ItbTPS基因家族成員在染色體上的位置基本一致。
2.6 Itf TPS和Itb TPS基因家族成員的組織表達分析
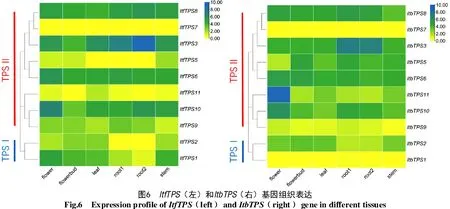
根據轉錄組數據繪制ItfTPS和ItbTPS在花、花蕾、葉、根、莖中的表達量熱圖,結果發現(圖6),TPSⅠ的ItbTPS1和TPSⅡ的Itf/ItbTPS7在各組織中基本無表達;Itf/ItbTPS8、Itf/ItbTPS6在各組織中均相對高表達。值得注意的是,Itf/ItbTPS2在根中的表達量最低,而Itf/ItbTPS3在根和葉中的表達量最高;在花中最高表達的基因分別為ItfTPS10和ItbTPS11。此外,ItfTPS9在各組織中的表達量均高于ItbTPS9,而ItbTPS5的表達量均高于各組織中的ItfTPS5,說明2個近緣野生種中對應基因的表達量具有差異性,且從熱圖可直觀看出,同種中同一基因在不同組織中的表達也具有差異性。
2.7 Itf TPS和Itb TPS基因家族成員在不同脅迫下的表達模式分析
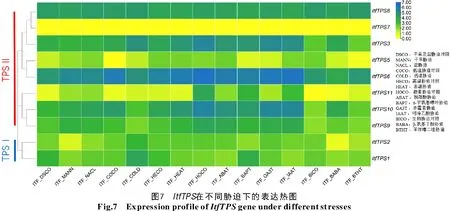
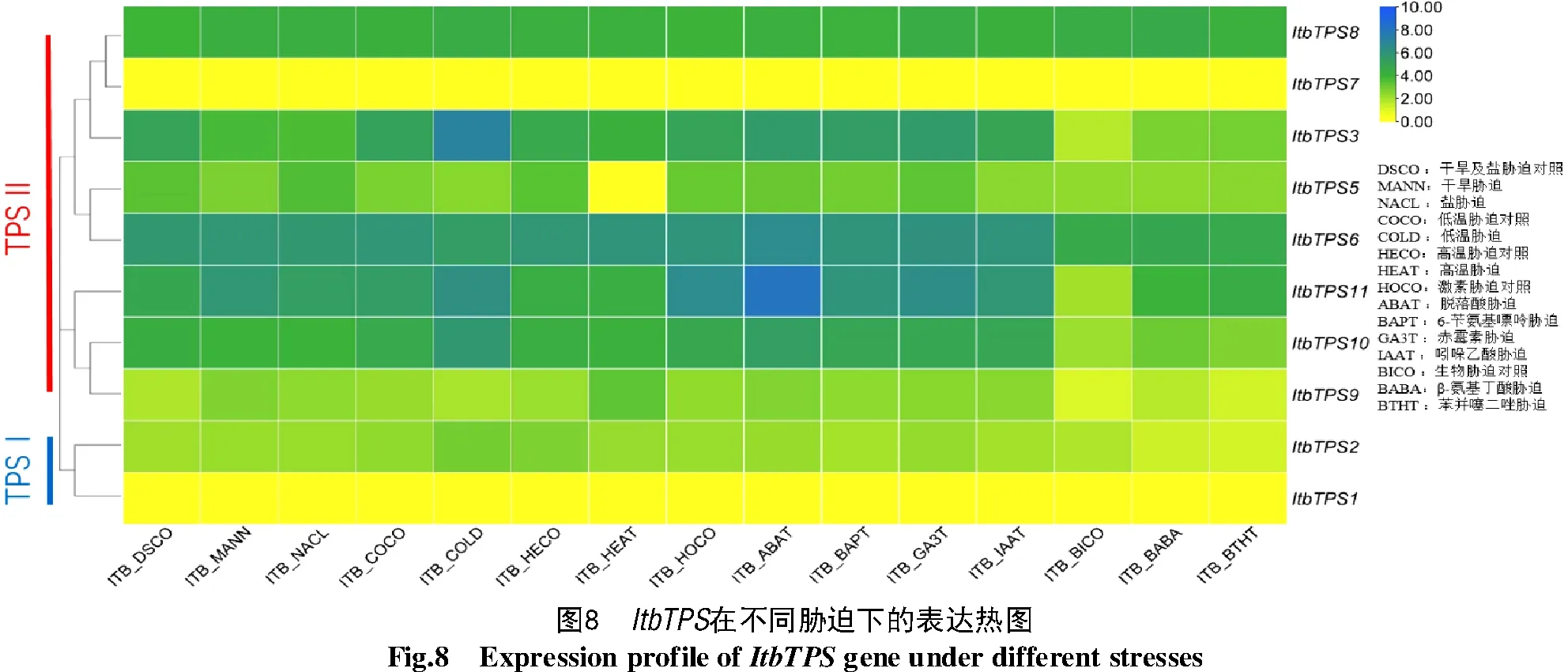
根據組培幼苗全株在不同脅迫下的轉錄組數據繪制的的表達熱圖可知(圖7、8),除Itf/ItbTPS7、ItbTPS1均無表達外,TPSⅡ的基因表達量相對較高,尤以Itf/ItbTPS6的表達量最高,而高表達的Itf/ItbTPS8在各脅迫間的表達無差異。Itf/ItbTPS6、Itf/ItbTPS11和ItfTPS9在干旱和鹽脅迫下表達量略有提升,推測這些基因可能參與干旱和鹽脅迫響應;ItfTPS1、ItfTPS3和ItbTPS10在生物脅迫下表達量有所提高,推測這些基因主要參與生物脅迫響應過程。值得關注的是,Itf/ItbTPS3在低溫脅迫下表達量升高;ItbTPS11噴施ABA后表達量升高,而ItfTPS11在ABA脅迫后表達量反而降低;ItfTPS5受低溫脅迫后表達量升高而ItbTPS5受高溫脅迫后表達量降低。此外,ItfTPS和ItbTPS基因在不同脅迫下的表達量并不完全相似,說明這2種近緣野生種的TPS基因在進化的過程中發生了不同的變異。
3 結論與討論
本研究在三淺裂葉牽牛和三裂葉薯中各鑒定出10個TPS基因,均含有GT 20_TPS和HAD_TPP結構域,其中,TPS結構域位于蛋白的N端,TPP結構域位于蛋白的C端,其結果與擬南芥[19]和水稻[20]的TPS蛋白一致;值得關注的是,由motif和保守結構域位置和長度的相關性可以推測,motif1、motif3、motif4是TPS結構域的主要組成部分,motif2、motif5、motif6是TPP結構域的主要組成部分。定位于第2、7、10號染色體上的Itf/ItbTPS1和Itf/ItbTPS8、Itf/ItbTPS6和Itf/ItbTPS10、Itf/ItbTPS2和Itf/ItbTPS3緊密相連,可以推測基因簇中的基因協同參與海藻糖-6-磷酸合成酶的合成或是共同調控植株的生長發育[21]。此外,亞細胞定位結果說明,ItfTPS和ItbTPS蛋白在胞內的分布區域不同,其功能可能也不同,位于細胞核的可能與基因的自我復制、轉錄過程有關;位于細胞中的可能參與部分基因的翻譯或翻譯后蛋白修飾;而位于線粒體基質的可能催化如三羧酸循環、脂肪酸氧化、氨基酸降解等重要的生化反應[22]。
系統進化樹結果表明,Itf/ItbTPS與AtTPS的親緣關系更近,根據擬南芥的聚類特征[23]進行命名,Itf/ItbTPS分為TPSⅠ和TPSⅡ兩大類,其中,Itf/ItbTPS1、Itf/ItbTPS2均與AtTPS1高度同源,而Itf/ItbTPS3與OsTPS1~OsTPS3高度同源。結合組織表達特異性,許多TPS基因家族成員在根和花中表達量相對較高,推測TPS基因可能對植株根和花的發育起調節作用,這與大豆[24]和擬南芥[25]中的表達結果相一致,已知AtTPS基因在擬南芥花中的過量表達能夠使擬南芥植株花序的分枝明顯增加[26],由此推測,在花中表達量較高的ItfTPS10和ItbTPS11基因也起著相似的作用。
研究發現,AtTPS1參與脅迫下的信號轉導,AtTPS5參與海藻糖的合成調控[26],而不同脅迫下的差異表達結果顯示,ItbTPS1在各組織不同脅迫下均無表達,故推測Itf/ItbTPS2是TPSⅠ族中參與脅迫的關鍵基因;ItfTPS5受低溫脅迫表達量升高,ItbTPS5受高溫脅迫表達量升高,推測Itf/ItbTPS5可能通過調控海藻糖合成而抵御溫度脅迫;Itf/ItbTPS6、Itf/ItbTPS11和ItfTPS9在干旱和鹽脅迫下表達量略有提升,推測這些基因可能參與干旱和鹽脅迫響應,但具體的脅迫響應調節機制還有待進一步驗證。
本研究從甘薯的2個近緣野生種三淺裂野牽牛和三裂葉薯的全基因組中各鑒定到10個Itf/ItbTPS,與AtTPS的親緣關系更近,聚為TPSⅠ和TPSⅡ兩大類,分別含有2個Itf/ItbTPS和8個Itf/ItbTPS。結構分析顯示,TPSⅠ中Itf/ItbTPS1、Itf/ItbTPS2分別含有17~18個外顯子,7~8個motif;而TPSⅡ成員含有1~3個外顯子,10個motif;所有Itf/ItbTPS家 族 成 員 均 含 有TPS(PF00982)和TPP(PF02358)保守結構域,且定位于2、7、10號的染色體上的Itf/ItbTPS1和Itf/ItbTPS8、Itf/ItbTPS6和Itf/ItbTPS10、Itf/ItbTPS2和Itf/ItbTPS3緊密相連。表達模式分析結果顯示,Itf/ItbTPS3可能對根的生長發育起著關鍵作用,ItfTPS10和ItbTPS11對花的生長發育起到關鍵作用;而Itf/ItbTPS5可能通過調控海藻糖合成而抵御溫度脅迫,Itf/ItbTPS6、Itf/ItbTPS11和ItfTPS9可能參與干旱和鹽脅迫響應。該結果為六倍體甘薯TPS家族的研究提供了參考,同時為甘薯抗逆種質培育提供了候選基因。
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
