三亞蜈支洲島海洋牧場區域夏季食物網研究
2022-05-20 09:46:56尹洪洋朱文濤馬文剛夏景全李建龍何法慶李秀保
生態學報 2022年8期
尹洪洋,朱文濤,馬文剛,章 翔,夏景全,許 強,李建龍,何法慶,李秀保,*
1 海南大學海洋學院,海口 570228 2 海南大學生態與環境學院,海口 570228 3 海南大學南海海洋資源利用國家重點實驗室,海口 570228
隨著熱力學理論的研究及Nier型質譜儀的不斷改進,穩定同位素技術(Stable Isotope Analysis,SIA)不斷發展,其主要用于食物網中營養結構和營養級的分析、確定食物來源及食物貢獻、生態位空間及水生生物洄游分布的研究等[1—5]。不同的碳源、碳氮比在珊瑚礁生態系統的結構和功能中所起的作用不同,可系統分析不同生物的營養關系、確定食物網以及食物網的相互作用關系等[6—8],如Graham等[9]利用250個珊瑚礁的經驗數據,分析了人為介導下的珊瑚礁魚類生物量的數量級梯度。海洋牧場生態系統功能復雜、漁業資源豐富,具有復雜的食物網結構[10]。目前,基于穩定同位素技術構建生態系統食物網分析不同生物的營養關系及生態系統的結構和功能已成為重要的研究手段之一[3]。有關食物網結構與功能的研究,國內外多采用穩定同位素方法來評估生態系統的營養關系,McMahon等[6]采用貝葉斯混合模型對穩定碳同位素比值(13C)進行分析,定量研究了紅海初級生產者對珊瑚礁魚類在不同攝食階段和營養位置的碳流量。郭衛東等[11]利用穩定碳同位素分析技術研究了南沙渚碧礁生態系食物網主要生物類群之間的營養關系。朱文濤等[12]以碳、氮同位素分析了大亞灣底棲群落營養結構的季節性變化。相較傳統胃含物技術的瞬時分析及樣本量大的局限性,SIA能反映一定時間的生物攝食和消化吸收情況,能更準確的量化評估生物的營養級[1—2,13]。Dromard等[14]運用穩定同位素技術及胃含物方法分析了加勒比海珊瑚礁草食性魚類的營養位置,發現SIA的結果較為準確。楊國歡等[15]根據δ13C、δ15N值分析珊瑚礁魚類營養層次并與傳統胃含物技術比較得出,SIA與傳統的胃含物分析法所得的結果有很好的一致性。
三亞蜈支洲島海域資源豐富,共記錄造礁石珊瑚13科40屬90種,珊瑚礁魚類33科52屬75種,大型無脊椎動物種類多樣性也非常豐富[16]。目前在該海域的研究主要集中于珊瑚礁修復技術[17]、環境脅迫對珊瑚生理狀態的影響[18—19]、人為因素對珊瑚群落的擾動[20]等,但關于三亞蜈支洲島牧場區域生態系統食物網結構的研究尚未見報道。本研究基于碳氮穩定同位素技術初步建立了三亞蜈支洲島牧場區域生態系統食物網,研究主要消費者穩定同位素特征,分析營養級層次并構建牧場區域食物網的連續營養譜。通過比較鄰近海域的營養結構特征,揭示食物網內不同層次消費者的主要食源,旨在為了解該生態系統營養結構提供基礎資料和參考數據。
1 材料及方法
1.1 樣品采集和處理
2020年7月對海南省蜈支洲島牧場區域共布設10個站點進行了生物樣品的采樣工作(圖1)。其中,核心區(珊瑚礁區域D1、D2、D3、D4)四個站位進行浮游生物、底棲生物及游泳生物調查;外圍區進行游泳生物樣品采集(以拖網作業為主),平均拖速為2.1節,每站拖網40 min左右。在核心區站位分別選用淺水Ш型和淺水Ⅱ型浮游生物網采集浮游植物和浮游動物;并以垂釣等方式采集核心區魚類;大型底棲生物由潛水及采樣帶法相結合的方法收集,采集生物樣品進行物種鑒定與同位素樣品處理。大型海藻經蒸餾水清洗干凈,烘干后使用錫箔紙包裹備用。取5 L表層海水經浮游植物網過濾后抽濾至灼燒后的GF/F濾膜上,經濃鹽酸酸熏5 h后烘干,所得樣品即懸浮顆粒有機物(POM);沉積有機物(SOM)及底棲微藻樣品采集制備方法參考徐軍[21],取適量沉積物研磨后,加入1 mol/L稀鹽酸進行酸化,充分反應至無氣泡產生,再用去離子水洗滌3次后烘干。在各個站點取適量礁石,刮取分離底棲藻類后,將樣品抽濾至灼燒后的GF/F濾膜直接烘干以測定底棲微藻同位素。浮游植物樣品用160 μm的篩絹進行過濾,過濾后的樣品抽濾到預先經馬弗爐450℃灼燒5 h的Whatman GF/F膜上,浮游動物挑選出雜質后過濾到同樣處理后的濾膜上。魚類取其背部肌肉,頭足類取胴體部位,蝦類取適量肌肉,蟹類取螯肢或腹部肌肉,海參取體壁肌肉,海膽取亞氏提燈附屬肌肉及性腺,海星取其性腺。

圖1 三亞蜈支洲島牧場區域采樣站位圖 Fig.1 Sampling stations in the marine ranch ecosystem of Wuzhizhou Island,SanyaC1—C4:核心區調查站位 Core area survey station;O1—O6:外圍區調查站位 Outer area survey station
經處理的上述樣品置于-50℃真空冷凍干燥機中冷凍干燥48 h至恒重,裝入研磨器中研磨60 s至粉末狀,存于5 mL棕色瓶中裝入自封袋后,冷凍保存以待后續的同位素樣品分析。
1.2 穩定同位素分析
所有樣品碳、氮穩定同位素比值通過元素分析儀(PYRO Cube,德國Elementar 公司)和穩定同位素質譜儀(Isoprime 100,英國 Isoprime公司)測定,碳氮同位素比值以 δ 值的形式表達。為保持實驗結果的準確性,每測試 10 個樣品后加測 1—2 個標準樣,個別樣品進行3次重復測定,δ13C 值和 δ15N 值的分析精度均為±0.2‰。碳、氮穩定同位素值計算公式如下:
δ=[(Rsample/Rstandard)-1]×103
式中,δ代表碳、氮同位素 (δ13C和δ15N);Rsample表示所測樣品的同位素比值 (13C/12C或15N/14N);Rstandard是國際上通用的標準物的同位素豐度之比,碳、氮穩定同位素標準物分別為咖啡因 (IAEA-600)和大氣氮。
1.3 不同食源對主要消費者貢獻率的計算
利用IsoSource模型計算三亞蜈支洲島牧場區域珊瑚礁食物網中主要食源對消費者的貢獻率,計算原理如下:
δjE=fAδjA+fBδjB+fCδjC+fDδjD
δkE=1×fAδkA+fBδkB+fCδkC+fDδkD
1=fA+fB+fC+fD
式中,jE、kE表示生物消費者E的2種同位素j、k的比值;A、B、C、D為食物源;f為食物對消費者的貢獻率。本研究利用Phillips[22]等利用質量守恒原理反復疊代編寫的IsoSource程序來計算各食源對珊瑚礁海域主要消費者的貢獻率。
1.4 數據處理及分析
以珊瑚礁生態系統中消費者的 δ15N 平均值來計算平均營養級,營養級的計算公式如下:
TL=(δ15Nsample-δ15Nbaseline)/△15N+λ

基于標準化歐氏距離(Standardised Euclidean distance)對三亞蜈支洲島牧場區域主要消費者的營養類群層次進行聚類分析,聚類分析(Hierarchical Cluster Analysis)方法采用瓦爾德法;基于IsoSource模型分析主要食源對消費者的貢獻率。單因素方差分析(ANOVA)檢驗三亞蜈支洲島珊瑚礁海域食物網各成分之間的穩定同位素差異。用 R 語言(R CoreTeam,2018)的 SIBER 模型計算食物網中的6個群落營養結構指標[26],利用Excel 2019、ArcGIS 10.2、SPSS 25.0、Origin 75、Adobe Illustrator 2020等軟件進行數據處理及繪圖。
2 結果分析
2.1 種類鑒定
2020年7月海洋生物調查共采集魚類24科24屬31種,頭足類3科3屬3種,蝦蟹類7科9屬10種,腹足類2科2屬2種,海參2科2屬3種、海膽2科2屬2種、海星1科1屬1種,大型海藻4科4屬4種(表1)。
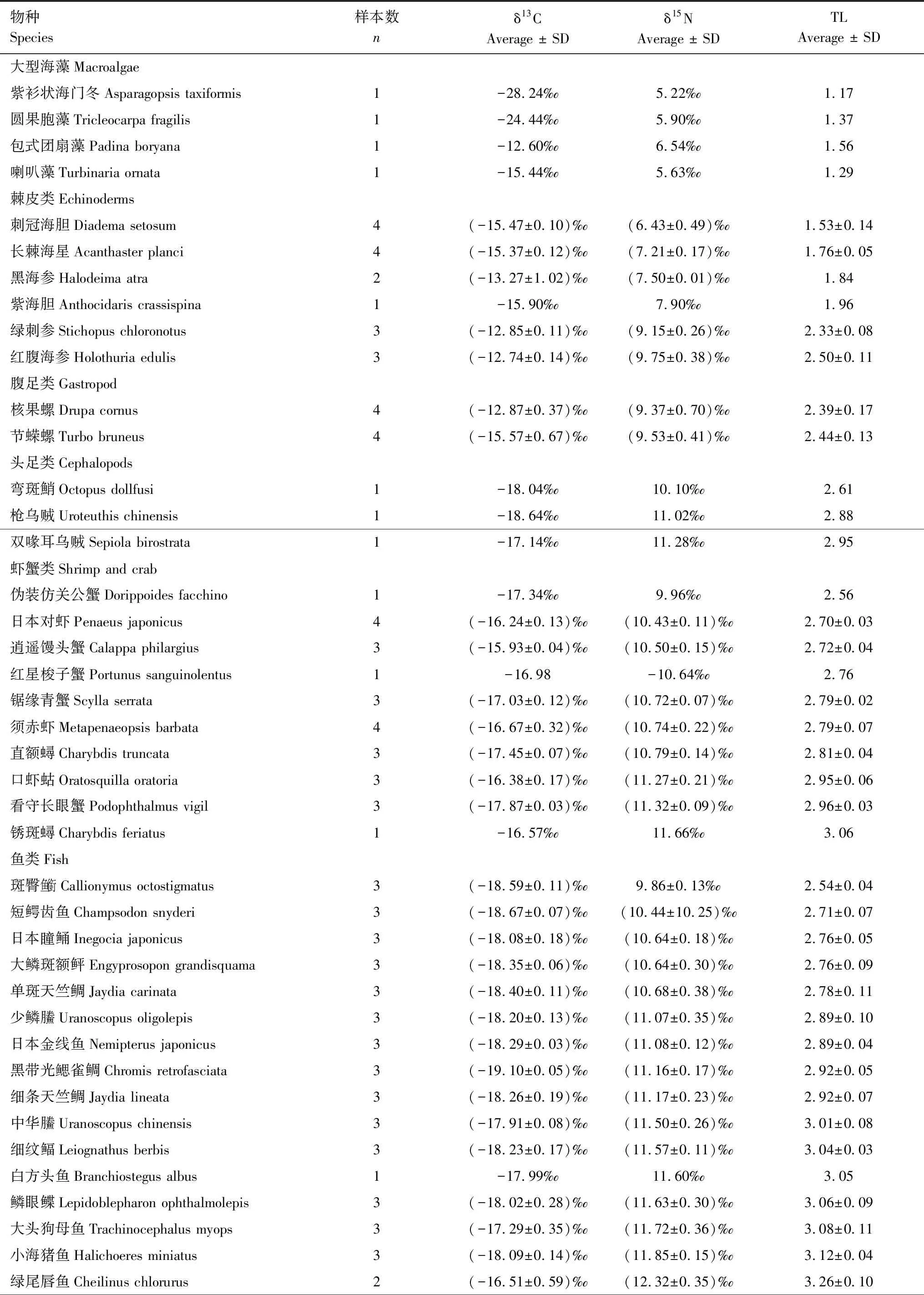
表1 三亞蜈支洲島牧場區域生態系統主要消費者δ13C、δ15N值及營養級(TL)Table 1 The δ13C,δ15N values and trophic level (TL) of major consumers in the regional ecosystem of Wuzhizhou Island Ranch in Sanya

續表物種Species樣本數nδ13CAverage ± SDδ15NAverage ± SDTLAverage ± SD中線天竺鯛Apogon kiensis3(-17.47±0.32)‰(-12.499±0.28)‰3.31±0.08蠕紋裸胸鱔Gymnothorax kidako1-18.14‰13.15‰3.35 東方豹魴鮄Dactyloptena orientalis1-17.23‰12.68‰3.36 須擬鲉Scorpaenopsis cirrosa1-15.56‰12.75‰3.39 克里裸胸鱔Gymnothorax cribroris1-18.37‰12.81‰3.40 線紋叉鼻鲀Arothron immaculatus1-18.384‰-12.86‰3.42 花斑蛇鯔Saurida undosquamis3-18.14‰13.15‰3.50 日本緋鯉Upeneus japonicus1-18.48‰13.22‰3.52 伯恩斯裸胸鯙Gymnothorax buroensis1-16.24‰13.23‰3.53 黃紋擬鱸Parapercis xanthozona3(-16.31±0.09)‰(13.55±0.12)‰3.62±0.03斑柄鸚天竺鯛Ostorhinchus fleurieu1-17.49‰13.60‰3.64 齒頜眶棘鱸Scolopsis ciliatus1-15.77‰13.64‰3.65 密花裸胸鯙Gymnothorax thyrsoideus1-16.91‰13.70‰3.66 帶紋躄魚Antennarius striatus117.28‰13.73‰3.67 橫帶九棘鱸 Cephalopholis boenak 1-16.90‰14.03‰3.76
2.2 穩定同位素特征

圖2 牧場區域生態系統主要消費者的 δ13C和 δ15N值的雙位圖Fig.2 The stable isotope biplots of the δ13C and δ15N values of the main consumers in the marine ranch ecosystem
2020年7月海洋生物調查除浮游生物、大型海藻外共采集52種主要消費者,以主要消費者的δ13C、δ15N值為橫縱坐標繪制珊瑚礁生態系統食物網的穩定同位素雙位圖(圖2)。
三亞蜈支洲島牧場區域珊瑚礁生態系統食物網δ13C值范圍為-19.10‰—-12.74‰,平均值(-16.99±1.52)‰;δ15N值范圍為6.43‰—14.03‰,平均值(11.24±1.70)‰(表1)。穩定同位素結果顯示,棘皮類δ13C、δ15N值范圍分別為-15.90‰(紫海膽Anthocidariscrassispina)—-12.74‰(紅腹海參Holothuriaedulis)和6.43‰(刺冠海膽Diademasetosum)—9.75‰(紅腹海參Holothuriaedulis),均值分別為(-14.27±1.33)‰、(7.99±1.14)‰;魚類δ13C、δ15N值范圍分別為-19.10‰(黑帶光鰓雀鯛Chromisretrofasciata)—-15.56‰(須擬鲉Scorpaenopsiscirrosa)和9.86‰(斑臀魚銜Callionymusoctostigmatus)—14.03‰(橫帶九棘鱸Cephalopholisboenak),均值分別為(-17.65±0.90)‰、(12.16±1.15)‰,δ13C、δ15N值跨度較大。蝦蟹類δ13C、δ15N值范圍分別是-17.87‰(看守長眼蟹Podophthalmusvigil)—-15.93‰(逍遙饅頭蟹Calappaphilargius)和9.96‰(偽裝仿關公蟹Dorippefacchino)—11.66‰(銹斑蟳Charybdisferiatus),均值分別為(-16.85±0.57)‰、(10.80±0.39)‰;腹足類δ13C、δ15N值范圍分別在-15.06‰—-13.33‰、9.37‰—9.53‰之間;頭足類δ13C、δ15N值范圍分別為-18.64‰—-17.14‰和10.10‰—11.28‰,均值分別為(-17.94±0.62)‰、(10.80±0.51)‰。單因素方差分析顯示,棘皮類、蝦蟹類、腹足類、魚類等不同類群δ13C、δ15N值差異極顯著(P<0.01)。
2.3 營養級及連續營養譜
本研究以浮游動物為基線生物,計算牧場區域珊瑚礁生態系統主要消費者的營養級并繪制連續營養譜。主要消費者的營養級范圍為1.53—3.76,均值為2.94±0.50,跨度為2.23,屬于3個營養級(圖3)。其中,橫帶九棘鱸Cephalopholisboenak營養級最高為3.76,刺冠海膽Diademasetosum營養級最低為1.53±0.14。各類群營養級之間差異較大,棘皮類、蝦蟹類、腹足類、頭足類和魚類的營養級范圍分別為1.53—2.50、2.56—3.06、2.39—2.44、2.61—2.95和2.54—3.76。珊瑚礁魚類的碳氮值跨度最大,而其他種群生物碳氮值相對跨度較小。

圖3 三亞蜈支洲島牧場區域生態系統主要消費者連續營養譜Fig.3 Continuous nutritional profile of major consumers in the regional ecosystem of Wuzhizhou Island Ranch in Sanya
2.4 牧場區域生物群落營養結構指標特征
進一步應用SIBER模型分析三亞蜈支洲島牧場區域穩定同位素數據的6個營養結構指標,通過與鄰近海域的研究結果比較,發現主要消費者的攝食來源多樣性水平(CR)、營養級長度(NR)分別為6.36和7.60,表明牧場區域珊瑚礁生態系統的食源多樣性水平較高,但食物鏈長度較短;生態位空間(TA)和平均營養級多樣性(CD)分別為29.52和1.96,說明該食物網營養級多樣性的總程度和平均程度相對較高;平均最近相鄰距離(MNND)及最近相鄰距離的標準偏差(SDNND)分別為0.37和0.30(表2)。
2.5 牧場區域的聚類分析及潛在碳源對不同消費者的貢獻率
通過系統聚類分析(Hierarchical Cluster Analysis)及IsoSource模型分析發現,大型海藻和底棲微藻對主要消費者的貢獻率最大為41.50%,沉積有機物(SOM)和浮游植物也是消費者的重要碳源(貢獻率分別為20.05%、19.97%),懸浮顆粒有機物(POM)對消費者的碳源貢獻率最低,為18.48%(表3)。
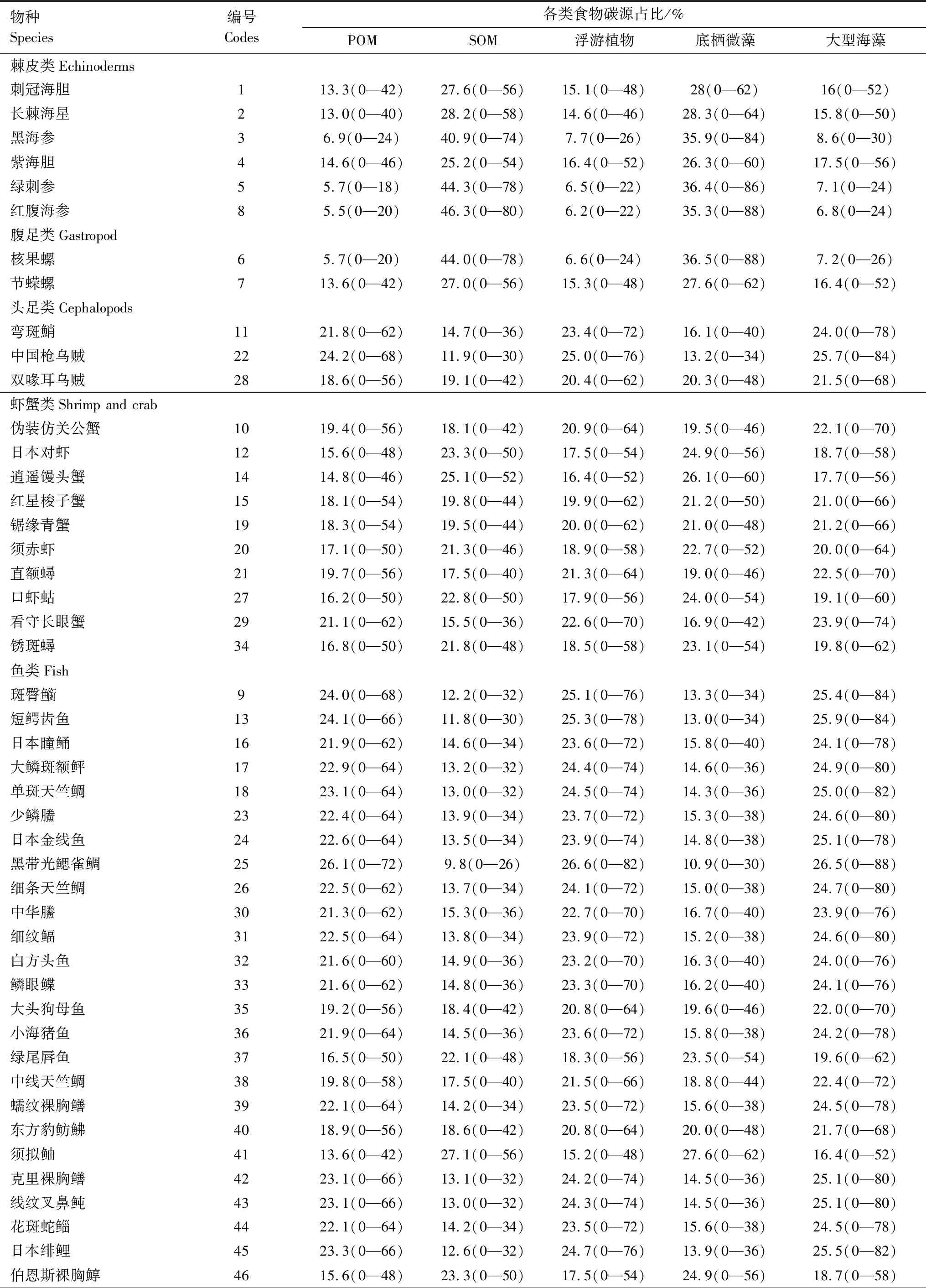
表3 三亞蜈支洲島牧場區域生態系統不同食物碳源的貢獻率Table 3 The contribution rate of different food carbon sources in the regional ecosystem of Wuzhizhou Island Ranch in Sanya

續表物種Species編號Codes各類食物碳源占比/%POMSOM浮游植物底棲微藻大型海藻黃紋擬鱸4716.0(0—46)23.2(0—50)17.7(0—56)24.3(0—56)18.9(0—60)斑柄鸚天竺鯛4819.9(0—58)17.3(0—40)21.4(0—66)18.8(0—44)22.7(0—72)齒頜眶棘鱸4914.2(0—42)25.8(0—54)15.9(0—50)26.9(0—60)17.1(0—54)密花裸胸鯙5017.9(0—54)20.3(0—44)19.6(0—60)21.4(0—50)20.8(0—64)帶紋躄魚5119.1(0—56)18.5(0—42)21.0(0—64)19.6(0—46)21.8(0—68)橫帶九棘鱸 5217.8(0—54)20.2(0—44)19.6(0—60)21.6(0—50)20.8(0—66)
3 討論3.1 三亞蜈支洲島牧場區域生態系統營養結構特征
CR 值表示食物網中的食源多樣性特征,CR值越大初始食源越多。本研究碳值跨度為6.36‰(即CR值),高于南海中西部漁場[27](3.49‰)、華南典型海灣[28](海陵灣2.44‰、陵水灣4.45‰)和浙江南部近海[30](5.70‰)。三亞蜈支洲島海洋牧場生態系統屬于典型南方熱帶海洋牧場生態系統,與鄰近海域相比,三亞蜈支洲島牧場區域生態系統中的食物來源除浮游植物、POM及有機碎屑外,還有自養細菌、珊瑚共生藻、大型海藻等多類初級生產者[10]提供碳源,這會導致生態系統內主要消費者在攝食餌料范圍上更廣,其碳源更為復雜。此外,三亞蜈支洲島海域臨近瓊東上升流區域[31],底層營養物質上涌使其更具高生產力,攝食來源多樣性水平(CR)的值可能會受其影響。但研究海域的CR值要低于膠州灣(8.47‰)[32]、珠江口(12.58‰)[24]的研究,這可能與南北方水質環境差異有關,南方地區有較高的日照水平,初級生產者穩定同位素組成可能出現明顯的碳富集[33—34],而且珠江口位于咸淡水交匯區域,水體、底質中有機質來源多樣化,水域環境的生物種類食源更為復雜[24]。三亞蜈支洲島牧場區域生態系統內主要消費者的NR值為 7.60,也明顯高于鄰近海域(表2),這可能與研究區域資源量有關,蜈支洲島牧場區物種多樣性高。牧場區中棘皮類、蝦蟹類等底棲動物,頭足類、腹足類等軟體動物以及游泳動物的存在可能使食物鏈長度更長。TA與CD表示食物網營養級多樣性的總程度和平均程度,MNND 和 SDNND 用來描述群落營養冗余的大小[35]。本研究中,生態系統的TA與CD值分別為29.52、1.92,MNND 和 SDNND值為0.37、0.30,與黃佳興[27]、張文博[28]、寧加佳[29]等人分別在南海中西部、南海西南部、陵水灣、海陵灣的研究結果比較,三亞蜈支洲島牧場區域生態系統的營養多樣性水平及平均程度較高。三亞蜈支洲島牧場區域食物網中具有相似營養特征的物種占多數,且營養生態位分布廣[36],因此冗雜程度也比鄰近海域高。

表2 不同區域主要消費者的群落結構指標Table 2 Community structure indicators of major consumers in different regions
三亞蜈支洲島牧場區域珊瑚礁生態系統的主要消費者可以分為4種營養類群:第一類主要以大型海藻、浮游植物、POM為主的混合有機碳源包括日本瞳鲬(Inegociajaponicus)、細紋鲾(Leiognathusberbis)、鱗眼鰈(Lepidoblepharonophthalmolepis)等近海中下層、底棲性魚類以及頭足類構成,占采樣總數的34.62%;第二類群食源以底棲微藻為主,SOM為輔,由紅星梭子蟹(Portunussanguinolentus)、日本對蝦(Penaeusjaponicus)等雜食性的蝦蟹類構成,占采樣總數的21.15%;第三類群主要包括橫帶九棘鱸(Cephalopholisboenak)、黃紋擬鱸(Parapercisxanthozona)、花斑蛇鯔(Sauridaundosquamis)、密花裸胸鱔(Gymnothoraxthyrsoideus)等肉食性魚類,主要食源以大型海藻為主,底棲微藻為輔,占采樣總數的30.77%;第四類群由紅腹海參(Holothuriaedulis)、紫海膽(Anthocidariscrassispina)等營底棲生活的棘皮類和腹足類組成,主要食源為SOM和底棲微藻;占采樣總數的13.46%(圖4),經單因素方差分析,各組群間具有顯著差異(P<0.01)

圖4 三亞蜈支洲島牧場區域主要消費者碳氮穩定同位素比值的聚類分析Fig.4 Cluster analysis of stable carbon and nitrogen isotope ratios of major consumers in Wuzhizhou Island Ranch area,Sanya
3.2 牧場區域食物網主要消費者的營養級
三亞蜈支洲島牧場區域食物網中主要消費者的營養級范圍為1.53—3.76,總體上三亞蜈支洲島魚類的平均營養級最高,其次為頭足類和蝦蟹類(平均營養級均為2.81),腹足類平均營養級為2.41,棘皮類平均營養級最低為1.99。根據國內學者營養層次的劃分標準[37]計算得出,刺冠海膽Diademasetosum、長棘海星Acanthasterplanci等4種雜食性動物和橫帶九棘鱸Cephalopholisboenak、花斑蛇鯔Sauridaundosquamis、齒頜眶棘鱸Scolopsisciliatus、密花裸胸鱔Gymnothoraxthyrsoideus等10種高級肉食性動物分別占總數的7.69%、19.23%;由紅星梭子蟹Portunussanguinolentus、大鱗短額鲆Engyprosopongrandisquama、彎斑鮹Octopusdollgusi以及日本瞳鲬Inegociajaponicus、大頭狗母魚Trachinocephalusmyops、銹斑蟳Charybdisferiatus等為代表的低級肉食性與中級肉食性動物分別占總數的32.69%、40.38%。牧場區域食物網的主要消費者分布在兩端(雜食性動物與高級肉食性動物)的只占少數,絕大多數居于中間的層次,為低級肉食性動物與中級肉食性動物(表1),與大亞灣珊瑚礁生態系統營養層次分布特征相似[12]。
本次調查采集31種魚類以中小型為主,營養級范圍為2.54—3.76,跨度1.22。三亞蜈支洲島魚類營養級主要集中于2.92—3.40之間,營養級別較高,約占魚類總數的49.21%。基準生物和采樣方式會對魚類營養級產生影響[38],本研究中基準生物取浮游動物,其δ15N值較低(8.04‰),采樣方式可能是導致魚類營養級較高的原因。三亞蜈支洲島珊瑚礁區域無法進行拖網采樣,本研究中魚類主要使用動物餌料垂釣獲得,具有一定的選擇偏向性,后續研究應考慮流刺網或地籠網等更全面的采樣方法。基于穩定同位素分析大部分魚類的營養級結果和Fishbase資料中數據相差不大,表明穩定同位素技術是研究海洋生物營養級別、分析其營養層次的可靠手段。但其中花斑蛇鯔Sauridaundosquamis和大頭狗母魚Trachinocephalusmyop營養級偏低,花斑蛇鯔營養級為3.50,高于楊國歡[15]在徐聞珊瑚礁關于長蛇鯔的胃含物分析(3.20),花斑蛇鯔與長蛇鯔屬于同科同屬,且食性相似,應具有相近的營養級別[39],而低于Fishbase資料中的4.50±0.4;而大頭狗母魚的營養級水平(3.08)也略低于楊國歡[15]的研究(3.20)和Fishbase資料中的數據4.40±0.4。多項研究表明體長的差異會影響魚類營養級[40—42]。Nakazawa[41]等研究發現掠食性魚類體長與δ15N值有呈現正相關的趨勢,張波等[42]研究幼魚階段赤鼻棱鰷營養級隨叉長增加而增加。本研究中采樣魚類平均體長為(8.0±5.52)cm,體型較小,魚類可能無法消耗高營養水平的較大獵物導致其食物種類范圍在低營養級群落中。而且本次調查采集的魚類主要為中小型魚類,一些鯊類、鰩類等大型肉食性魚類[11,27,29]可能未被采集到,這也可能是導致營養級偏低的原因。另外,本研究中基準生物選擇為浮游動物,運用整體的浮游動物的δ15N值為基準估算魚類營養級是有局限性的,浮游動物不同大小、不同種類、營養物質來源以及季節性變化等都會對營養級估算產生影響[38]。
本次采樣調查中頭足類的平均體長為(4.7±2.17)cm。頭足類會攝食蝦蟹類及魚類,應位于營養層次較高的位置,但頭足類胴長會顯著影響其δ13C和δ15N值,在稚魚階段頭足類常以端足類、糠蝦等小型甲殼類為食[43—44]。聚類分析中,第二類群除蝦蟹類外,雙喙耳烏賊也被聚類,說明具有相似食性。此次采樣雙喙耳烏賊體長范圍在3.0—4.2 cm之間,體重在8.78—24.21 g之間,體型較小。在南海中西部研究中也發現,頭足類平均營養級隨著體長(胴長)增大其營養級有相應增大的趨勢[27],因此,兩類平均營養級相同可能與采集樣品中頭足類的生長階段及個體大小有關。另外,蝦蟹類中銹斑蟳營養級最高,銹斑蟳食性比較復雜,會攝食甲殼類、藻類、魚類等[45],張文博[28]、高春霞[30]、莫寶霖[46]等人研究中銹斑蟳也具有較高的δ15N值或營養級。棘皮類中營養級最高的生物是紅腹海參,與其他棘皮動物相比,紅腹海參除攝食沉積物、浮游生物外,還會攝食海草、藻類等[47]。食性差異可能是導致銹斑蟳與紅腹海參營養級相對較高的原因。
3.3 牧場區域珊瑚礁食物網主要消費者食源分析
浮游植物、底棲微藻、大型海藻在海洋牧場生態系統中是重要的潛在食源,對確定食物網中的主要碳源具有重要意義[6,48—49]。穩定同位素分析在海洋牧場區域主要消費者食性來源方面的研究已有諸多報道。本研究中,底棲微藻貢獻率為20.59%,大型海藻占比20.91%。其中,底棲微藻在蝦蟹類、腹足類和棘皮類為主的第二、四類群生物中攝食最高,平均貢獻率分別為21.84%、32.05%和32.44%,Briand[7]等也發現基于底棲藻類的底棲食物網為大多數中間消費者、雜食性魚類提供了重要的食物來源。δ13C值富集程度可用于示蹤及分析生態系統食物鏈[50—51],碳同位素高的生物基本與底棲食物網有關[30]。經過單因素方差分析,魚類與蝦蟹類、腹足類、棘皮類的穩定碳同位素之間呈現極顯著性差異(P<0.01),魚類與棘皮類差值最大(6.36%),δ13C值富集程度較低,研究區域可能存在以底棲微藻為食源基礎的底棲食物鏈。本研究中,第三類群魚類的主要食源是大型海藻,在海洋牧場生態系統中,大型海藻與珊瑚會有一定的競爭機制,但在營養物質的吸收與釋放、生物餌料、空間庇護等方面也有重要作用[52—53]。如大型海藻在黑帶光鰓雀鯛與細條天竺鯛中的碳源占26.5%、24.7%,蔣日進等[54]發現在枸杞島食物網中魚類的主要碳源也是大型海藻,McMahon[6]等利用貝葉斯模型研究表明大型海藻是紅海珊瑚礁區域食物網的重要基礎。SOM也是該海域生物食源的組成部分,是重要碳源。基于IsoSource模型分析,本研究中海參(黑海參、綠刺身、紅腹海參)、海膽(刺冠海膽、紫海膽)等棘皮類的SOM貢獻率最高,平均貢獻率分別占43.83%、26.4%。SOM是棘皮類的基礎食源,這與魏虎進[55]在象山港海洋牧場區研究發現SOM是食物網的營養基礎和莫寶霖[46]基于穩定同位素技術分析大亞灣紫海膽食性的結果相似。
4 結論
本研究構建了三亞蜈支洲島海洋牧場生態系統食物網,分析牧場區域內主要消費者的營養級特征,應用SIBER模型對比牧場區域與鄰近海域的營養結構差異,進一步利用聚類分析與IsoSource模型揭示主要消費者的食源,為海洋牧場生態系統食物網的構建及實現基于生態系統的漁業管理提供科學的理論依據。在后續研究中,應針對牧場內珊瑚礁生態系統的規模、穩定性和成熟度研究其內部的物質循環和能量流動,分析海洋牧場關鍵捕食者或關鍵生境改造者在珊瑚礁區域的重要性。
猜你喜歡
今日農業(2021年9期)2021-11-26 07:41:24
今日農業(2021年11期)2021-08-13 08:53:34
發明與創新·小學生(2021年3期)2021-03-25 11:48:49
中國生殖健康(2019年8期)2019-01-07 01:18:24
幸福(2018年33期)2018-12-05 05:22:46
作文周刊·小學一年級版(2017年17期)2017-06-27 19:32:55
廣東第二課堂·小學(2017年2期)2017-02-20 15:10:08
中國科技博覽(2016年2期)2016-04-25 20:32:39
小學生導刊(2016年34期)2016-04-11 00:49:44
海峽姐妹(2016年5期)2016-02-27 15:20:20
