呼倫貝爾沙地樟子松生產力及其對氣候因子的響應
2022-05-24 06:57:30牛進松劉小粉SeMyungKwon潘磊磊時忠杰楊曉暉張日升
林業科學研究 2022年3期
牛進松,劉小粉,SeMyung Kwon,潘磊磊,時忠杰,楊曉暉,張 曉*,張日升
(1. 河北工程大學園林與生態工程學院,河北 邯鄲 056038;2. 中國林業科學研究院荒漠化研究所,北京 100091;3. 中國林業科學研究院生態保護與修復研究所,北京 100093;4. 韓國公州國立大學生態修復學院,忠清南道 32439;5. 遼寧省沙地治理與利用研究所,遼寧 阜新 123000)
最新發布的第六次氣候變化評估報告顯示,2019 年大氣CO2濃度高達410 ppm,是工業化前的148%,人類活動與氣候變化導致全球地表溫度進一步升高,2011—2020 年地表溫度比1850—1900 年升高1.09 ℃[1]。森林覆蓋全球陸地面積的30.6%,是陸地生態系統中最大的碳庫,在緩解CO2濃度升高和全球變暖方面發揮著重要作用[2-3]。森林凈初級生產力是植物在扣除自身呼吸后,剩余的單位面積單位時間的有機物增量,能夠表示森林的碳匯功能,因此,研究森林中主要植被的生物量和凈初級生產力及其與氣候間的關系,有助于評估森林生態系統的固碳功能,并預測未來氣候變化下該系統的功能穩定性。
目前,森林凈初級生產力及碳儲量的估算方法主要包括:基于森林清查資料的估算方法、遙感反演法及模型模擬法等[4],但這些方法均難以獲取樹木生長歷史中長時間序列的生物量及生產力,樹木年輪方法利用年輪寬度得到逐年胸徑,并與生物量方程結合,能夠得到樹木整個生長歷史中的生物量及凈初級生產力,因此,樹木年輪法在研究植被的凈初級生產力及其氣候響應中具有較大的優勢且已有較多的應用,如于健等[5]分析了長白山紅松(Pinus koraiensisSieb. et Zucc.)林生產力的年際動態及其對氣候因子的響應,王云霓等[6]利用華北落葉松年輪寬度反演了其碳密度的歷史動態,劉立斌等[7]對比了貴州喀斯特地區與非喀斯特地區主要闊葉樹與針葉樹的地上生物量及生產力。
樟子松(Pinus sylvestrisvar.mongolicaLitv.)又稱歐洲赤松,天然分布在我國大興安嶺和呼倫貝爾沙地、俄羅斯和蒙古的部分地區[8]。由于沙地樟子松具有耐寒、耐旱、耐貧瘠及生態適應性強等特點,已被廣泛應用到我國北方沙地造林中,在防風固沙、水土保持及森林固碳等方面發揮重要的生態功能[9]。第八次森林資源清查(2009—2013 年)結果顯示,目前我國沙地樟子松人工林面積達到41.7 萬hm2,僅分布在科爾沁、渾善達克和毛烏素沙地的樟子松人工林面積就達33 萬hm2,赤峰市計劃在“十二五”至“十四五”期間完成100 萬hm2樟子松林基地的建設。目前已有學者針對樟子松林凈初級生產力及其氣候響應展開研究,叢俊霞等[10]結合地面調查及遙感數據分析了呼倫貝爾樟子松天然林的地上碳密度及空間分布特征,并提出了分區經營以提高碳匯的建議;曹恭祥等[11]研究了41 年生呼倫貝爾沙地樟子松人工林的固碳速率及對氣候的響應,目前針對不同密度樟子松林的凈初級生產力及其對氣候響應的對比研究尚未開展。密度通過調整樹木間競爭來影響樹木生長及生物量累積,是影響林分凈初級生產力的重要因素,間伐是通過調整林分密度來改善林分結構、生產力及其功能的一種重要的森林經營管理措施,更是森林生態研究的熱點。
通常,位于天然分布邊緣區的植被具有更高的氣候敏感性,更能預測未來氣候變化對該植被的潛在影響。本研究以位于地理分布南緣的呼倫貝爾沙地不同密度的天然樟子松林為研究對象,結合樹木年輪學方法與生物量方程計算樟子松林的生物量及凈初級生產力,對比不同密度天然樟子松林的凈初級生產力及其對氣候的影響,不僅有助于預測未來氣候變化下天然樟子松林的生態功能,也為預估我國北方營造的大規模人工樟子松林的固碳潛力及其制定適應未來氣候變化的營林措施有重要意義。
1 材料與方法
1.1 研究區概況
研究區位于呼倫貝爾沙地東南端、新巴爾虎左旗罕達蓋蘇木中蒙邊界附近(47°38′~47°39′ N,119°09′~119°12′ E),屬中溫帶半干旱大陸性氣候,夏季溫暖,冬季嚴寒、日照充足,晝夜溫差大,年均氣溫?2.6 ℃,年降水量 270~650 mm(圖1),多集中于夏秋季,年蒸發量1 400~1 900 mm,無霜期 90~100 d,積雪期長達 140 d左右[12]。研究區以固定沙地和半固定沙地為主,土壤為風沙土[13],處于森林-草原過渡帶上,是沙地樟子松種群在我國地理分布的最南緣。本區的樟子松林多以純林為主,并伴生有白樺(Betula platyphyllaSuk.)、榛 子(Corylus heterophyllaFisch.)等喬灌樹種,以及線葉菊(Filifolium sibiricum)、大針茅(Stipa grandis)、羊草(Leymus chinensis)、日陰菅(Carex pediformis)、地榆(Sanguisorba officinalis)等草本物種[14]。

圖1 研究區1960—2016 年的年平均氣溫和年降水量Fig. 1 Changes of annual mean air temperature and precipitation from 1960 to 2016
1.2 樣地設置與調查
2017 年7—9 月,在人為干擾較小的沙地樟子松天然林選擇5 個不同密度的標準樣地(30 m ×30 m),記錄樣地的海拔、坡度等立地因子,并進行每木檢尺,測定樣地內所有樹木的胸徑、樹高、枝下高及東西南北4 個方位的冠幅等樹木生長信息,胸徑徑級分布見圖2。樟子松的林分密度利用胸高斷面積來表示,利用胸徑計算每個樣地的胸高斷面積分別為8.83、21.94、24.28、32.36 和34.9 m2hm?2(表1)。利用內徑為5.15 mm 的瑞典產Haglof 生長錐沿東西、南北兩個交叉方向對所有胸徑大于10 cm 的樹木進行胸徑位置(1.3 m)處的樹芯取樣(2 芯/樹),盡量取到髓芯。

圖2 不同密度天然樟子松林的胸徑徑級分布Fig. 2 Diameter at breast height (DBH) distribution of naturalPinus sylvestrisvar.mongolicaLitv. forest with different density
1.3 樹木年輪寬度測定
將采集的樹芯帶回實驗室風干,利用白乳膠將樹芯粘貼、固定于木制樹芯凹槽內,并依次利用由粗到細不同粒度(240 目、600 目和800 目)的砂紙進行打磨直至肉眼清晰可辨樹木的年輪界限[15]。在顯微鏡下用骨架圖法進行目視交叉定年,利用精度為0.01 mm 的Lintab 6 樹木年輪分析儀(Frank Rinn,德國)與TSAP 軟件進行年輪寬度測量,利用 COFECHA 程序[16]對所有的樹輪寬度測量結果進行交叉定年檢驗,手動調整或消除定年不準確和相關性較低的樹芯樣本,5 個樣地的樹輪年表統計特征見表2。5 個天然樟子松林的平均樹齡均在23 a 左右(表1),樣地1 和樣地2 僅個別樹木的年齡為40 a 以上,因此研究樣地樟子松林處于相似的生長發育階段,其生產力的差異主要來自于林分密度影響。

表1 樣地基本情況Table 1 Basic information of sample plot
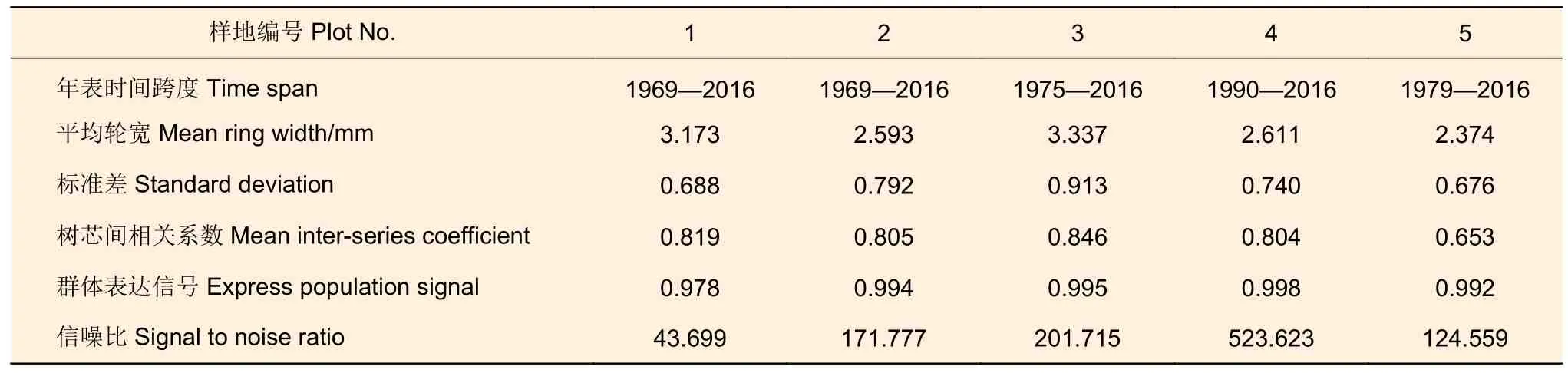
表2 5 個不同密度樣地樹輪寬度年表的統計特征參數Table 2 Chronology statical characteristic of tree-ring width of 5 sampling plots with different densities
1.4 生物量和凈初級生產力的估算
根據樹芯取樣年份(2017 年)測定的樹木胸徑及逐個樹木的年輪寬度原始數據,減去后面所有年份的年輪寬度即得到這一年份的樹木胸徑值。然后,根據以樹木胸徑為參數的樟子松生物量方程計算得到這一年份的樹木各組分生物量及總生物量,樹干、枝、葉的生物量方程來自叢俊霞等[17],根的生物量方程來自董利虎[18],樟子松林的總生物量為樣地內所有樹木各組分的累積生物量:

其中,W干、W枝、W葉、W根依次為樟子松樹木的干、枝、葉及根的生物量(kg),W為樣地內所有樹木的總生物量(kg·hm?2),DBH為樹木胸徑(1.3 m 處)(cm),m為每個調查樣地的樟子松株數,S為樣地面積(hm2)。
樣地的年凈初級生產力(NPP)為樣地歷年單位面積上的生物量之差:

其中,NPPi是第i年的凈初級生產力(t·hm?2·a?1),Wi、Wi分別表示第i年與前一年的總生物量(t)。
1.5 數據獲取及分析
氣象數據來源于國家氣象科學數據中心(http://data.cma.cn),下載臨近的阿爾山氣象站(47°10′ N,119°56′ E)1960—2016 年的月平均氣溫、月平均最高氣溫、月平均最低氣溫和月降水量數據。帕爾默干旱脅迫指數(Palmer drought severity index,PDSI)利用荷蘭皇家氣象研究所數據共享網(http://climexp.knmi.nl/)中的格點數據,該數據集為利用CRU TS 3.26 計算而來的自校正PDSI數據(scPDSI,時長1901—2017),經緯度的精度都為0.5°[19]。下載的NC 格式scPDSI數據利用美國國家航空航天局(NASA)開發的Panoply 4.11.1 軟件(https://www.giss.nasa.gov/tools/panoply/)讀取。本研究選取天然樟子松林樹木生長的公共時間段進行凈初級生產力與氣候因子的相關分析,即1990—2016 年。考慮到上一年氣候對當年樹木生長存在明顯影響[20-21],相關分析的時間跨度為上一年6 月至當年12 月。統計分析利用SPSS Statistics 20 完成,利用Pearson 相關分析凈初級生產力與氣候因子間關系,繪圖利用Excel 2010 完成。
2 結果與分析
2.1 凈初級生產力及生物量的變化
不同密度樟子松天然林凈初級生產力年際波動趨勢基本一致,所有密度樟子松林的1998 年凈初級生產力均較大,最大值為樣地4 的11.9 t·hm?2,最小值為樣地1 的1.2 t·hm?2,2007 年的凈初級生產力較小,所有林分的年凈初級生產力均在2 t·hm?2以內(圖3)。5 個樣地天然樟子松林的多年平均凈初級生產力分別為1.17、2.99、3.55、4.69 和4.86 t·hm?2,平均凈初級生產力隨胸高斷面積增加而增加。

圖3 不同密度樣地樟子松天然林年凈初級生產力Fig. 3 Annual net primary productivity ofPinus sylvestrisvar.mongolicaLitv.natural forest with different density
不同密度天然樟子松林的生物量逐年增高,1990—2016 年的27 年間樣地1 增長幅度最小,為31 t·hm?2,樣地2 增長了79 t·hm?2,樣地3 增長了100 t·hm?2,樣地4 增長了125 t·hm?2,樣地5 增長幅度最大為130 t·hm?2(圖4)。隨林分胸高斷面積增加,天然樟子松林的生物量逐漸增加,2016 年5 個樣地的樟子松林生物量分別為38.4、91.5、102.4、126.6 和132.9 t· hm?2。

圖4 不同密度樟子松天然林生物量Fig. 4 Biomass ofPinus sylvestrisvar.mongolicaLitv. natural forests with different densities
2.2 樟子松凈初級生產力對氣溫的響應
不同密度天然樟子松林的凈初級生產力與平均氣溫的相關分析表明溫度對樟子松林凈初級生產力無明顯影響,僅樣地5 樟子松林凈初級生產力與前一年7 月平均氣溫顯著正相關(P<0.05),樣地4 樟子松林凈初級生產力與當年9 月平均氣溫顯著正相關(P<0.05),其余密度天然樟子松林的凈初級生產力與平均氣溫的相關關系均不顯著(圖5a)。
與平均氣溫影響一致,樣地4 和樣地5 天然樟子松林凈初級生產力與前一年7 月的平均最高氣溫顯著正相關(P<0.05),相關系數分別為0.392和0.407;樣地5 的天然樟子松林與當年9 月的平均最高氣溫顯著正相關(P<0.05),其余樣地林分凈初級生產力與平均最高氣溫的相關性均不顯著(圖5b)。
天然樟子松林凈初級生產力與前一年及當年的月平均最低氣溫大多無顯著相關關系,僅低密度天然樟子松林(樣地1 和樣地2)凈初級生產力與前一年6 月平均最低氣溫顯著正相關(圖5c)。天然樟子松林凈初級生產力與氣溫相關性不大,僅秋季9 月的月平均氣溫和月平均最高氣溫對大密度樟子松林有顯著的正相關關系,表明大密度天然樟子松林生物量累積僅受秋季9 月最高氣溫的限制,較低的月平均最高氣溫影響了樟子松林的生長。總體來說,氣溫不是限制研究區天然樟子松林凈初級生產力的主要氣候因子。

注:p6…p12 表示前一年6 月…12 月;c1…c12 表示當年1 月…12 月;虛線表示在0.05 水平上顯著。Note: P6...P12 means June...December of the previous year; C1...C12 means January...December of the current year; the dotted line means significant at the 0.05 level.
2.3 樟子松凈初級生產力對降水的響應
不同密度樟子松天然林凈初級生產力與月降水量的相關分析表明:樣地1 的天然樟子松林凈初級生產力與前一年8 月降水顯著正相關(P<0.05),所有密度的樟子松林凈初級生產力均與當年5 月降水顯著正相關(P<0.05),樣地1 的樟子松林凈初級生產力與當年7 月降水顯著正相關(P<0.05),樣地4 的樟子松林凈初級生產力與當年10 月降水顯著正相關(P<0.05)(圖6)。當年5 月降水是限制樟子松林凈初級生產力的主要氣候因子,這與樟子松樹木進入生長期及相對缺乏的降水有關。
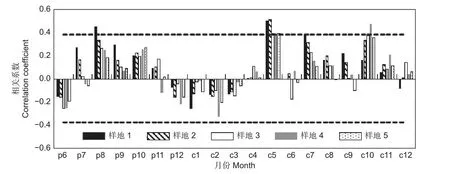
注:p6…p12 表示前一年6 月…12 月;c1…c12 表示當年1 月…12 月;虛線表示在0.05 水平上顯著。Note: P6...P12 means June...December of the previous year; C1...C12 means January...December of the current year; the dotted line means significant at the 0.05 level.
2.4 樟子松凈初級生產力對干旱指數的響應
不同密度天然樟子松林凈初級生產力與帕爾默干旱脅迫指數的相關性見圖7,結果表明樣地1 的天然樟子松林凈初級生產力與當年1 至7 月的PDSI均呈顯著正相關關系(P<0.05),樣地2 的天然樟子松林凈初級生產力與當年6 至7 月的PDSI顯著正相關(P<0.05),樣地3 的天然樟子松林凈初級生產力與當年5 月的PDSI顯著正相關(P<0.05),相關系數為0.409。此外,在前一年11 月至當年7 月,天然樟子松林凈初級生產力與PDSI的相關系數隨密度增加呈減小趨勢,表明相比于高密度天然樟子松林,低密度天然樟子松林的凈初級生產力對PDSI的響應更為敏感。

注:p6…p12 表示前一年6 月…12 月;c1…c12 表示當年1 月…12 月;虛線表示在0.05 水平上顯著。Note: P6...P12 means June...December of the previous year; C1...C12 means January...December of the current year; the dotted line means significant at the 0.05 level.
3 討論
3.1 樟子松林的生物量與凈初級生產力
2016 年5 個不同密度的天然樟子松林生物量分別為38.4、91.5、102.4、126.6 和132.9 t·hm?2,5 個不同密度天然樟子松林在1990—2016 年的年平均凈初級生產力分別為1.17、2.99、3.55、4.69和4.86 t·hm?2。隨林分基面積增加,天然樟子松林的生物量及多年平均凈初級生產力逐漸增加。小興安嶺幼齡、中齡、近熟及成熟的樟子松林生物量分別為47.25、106.25、143.09 和185.01 t·hm?2[22],苑增武等[23]研究了兩個林場內人工樟子松林的多年平均凈初級生產力,數值分別介于0.53~8.17 t·hm?2和0.99~6.68 t· hm?2,本研究天然樟子松林的生物量及多年平均凈初級生產力值多位于過去研究值范圍之內。
本研究中各樣地樟子松林地上生物量分別為31.60、74.51、83.70、101.30 和105.43 t·hm?2,5 個不同密度的天然樟子松林地上生物量遠遠低于黑龍江省西部地區的154.07 t·hm?2[23],而屬于高密度林的樣地4 和樣地5 高于黑龍江大興安嶺45 a的(94.87 t·hm?2)、小 興 安 嶺45 a 的(80.50 t·hm?2)[22]、塞罕壩地區45 a 左右樟子松林地上生物量(94.20 t·hm?2)[24]和內蒙古科爾沁沙地35 a 的(86.45 t·hm?2)[25]。
生物量或生產力的差異可能是由立地條件、林齡、林分密度、氣候等因素引起的。袁立敏等[25]研究認為:科爾沁沙地樟子松林生物量隨著林齡的增加而增大,王云霓等[6]發現華北落葉松同齡人工林生物量具有顯著的坡位差異,賈忠奎等[26]發現塞罕壩華北落葉松同齡人工林生產力具有顯著的坡向差異,叢俊霞[10]研究得出結論樟子松天然林植被碳儲量隨海拔升高先增加后減少,隨坡度增大急速減少,在海拔和坡向上無明顯規律。以上因素可能是本研究中樟子松林生物量與生產力與其它研究結果存在差異的原因。
3.2 天然樟子松林凈初級生產力對氣候因子的響應
不同密度的天然樟子松林凈初級生產力均受到溫度、降水和PDSI的影響。高密度的樣地4 和樣地5 天然樟子松林凈初級生產力分別與當年9 月平均氣溫和當年9 月平均最高氣溫顯著正相關;所有密度的天然樟子松林凈初級生產力均與當年5 月降水顯著正相關,樣地1 與樣地4 的凈初級生產力還分別與當年7 月和10 月降水顯著正相關;樣地1、2 和3 的凈初級生產力分別與當年1—7月、當年6—7 月和當年5 月的PDSI顯著正相關,還呈現出隨胸高斷面積增加,凈初級生產力與PDSI的相關系數減小的趨勢。由于呼倫貝爾沙地9 月氣溫逐漸降低,影響了樟子松的光合作用及生物量積累,且在密度較大林分競爭大,對溫度抑制生長的過程表現更加敏感,這導致了9 月平均氣溫、平均最高氣溫與高密度天然樟子松林凈初級生產力的顯著正相關關系。
呼倫貝爾沙地天然樟子松樹木在5 月進入樹木生長期,樹木生長需水與5 月相對較少的降水導致了所有密度天然樟子松林凈初級生產力與5 月降水間的顯著正相關關系[27]。10 月份進入沙地天然樟子松生長的末期,降水不足也同樣導致了樣地4 林分凈初級生產力對當年10 月降水具有較高的敏感性。此外,樣地1 天然樟子松林凈初級生產力與上一年8 月降水顯著正相關,可能是由于前一年降水充足為樹木體內儲存了較多的養分,進而為下一年生長提供了更好的營養條件[28]。PDSI綜合考慮降水、氣溫等因素的影響,是衡量植物生長可利用水分的重要指標[29],在前一年11 月至當年7 月,天然樟子松林凈初級生產力與PDSI的相關系數隨密度增加呈減小趨勢,可能是由于相比于氣候因素,高密度天然樟子松林的凈初級生產力更多地受到了競爭等生物因素干擾的影響。
很多研究表明:樹木的氣候敏感性隨地理位置(緯度、經度或海拔高度)、土壤等立地條件和樹木間競爭等影響而存在分異[30-31]。本研究也表明競爭強度的差異導致了不同密度天然樟子松林凈初級生產力對氣候因子響應的差異,與氣溫、水分及水熱綜合因子PDSI等氣候因素相比,高密度樟子松林的凈初級生產力還受林內樹木間競爭的較大影響,使其與PDSI的相關系數低于低密度的天然樟子松林。氣候變化導致全球多地干旱事件的頻率與程度增加,并引起大范圍的森林衰退與樹木死亡[32],在此背景下,間伐作為重要的森林管理措施,被認為在調節林分結構、競爭及提升應對干旱抵抗力等方面有積極作用[33],本研究明確了密度對天然樟子松凈初級生產力及其氣候響應的影響,不僅為未來氣候變化對樟子松林生產力影響提供依據,還為樟子松天然林與人工林的經營管理措施制定提供理論依據。
4 結論
天然樟子松林的生物量及多年平均凈初級生產力隨林分密度增加而增加,高密度天然樟子松林具有更高的生物量及生產力,本研究中最高密度林分生產力及生物量分別是最低密度林分生產力及生物量的4.2 和3.5 倍。通過分析不同密度天然樟子松林凈初級生產力對氣溫、降水與帕爾默干旱脅迫指數的響應,發現高密度天然樟子松林的凈初級生產力主要受9 月平均氣溫及平均最高氣溫的影響,低密度天然樟子松林的凈初級生產力主要受6—7 月干旱指數影響,所有密度天然樟子松林均與5 月降水顯著正相關,表明5 月降水是限制天然樟子松林的關鍵因子。本研究還發現隨林分密度增加,凈初級生產力與干旱指數的相關系數減小,表明了林分競爭對天然樟子松凈初級生產力存在明顯影響,林分競爭導致了高密度天然樟子松林凈初級生產力對干旱指數的敏感性降低。本研究明確了呼倫貝爾沙地不同密度天然樟子松林的生物量及凈初級生產力動態變化,并分析了不同密度天然樟子松林凈初級生產力對氣候因子的響應,有助于預測未來氣候變化下天然樟子松林的生產力及固碳功能,也能為制定相應的森林經營管理措施提供支撐。
