
馬鈴薯與曼陀羅交互嫁接后藥用生物堿的積累特征
2022-05-27 08:30:40郭開基李有涵郭華春
貴州農(nóng)業(yè)科學(xué) 2022年5期
郭開基, 李有涵, 王 瓊, 郭華春*
(1.云南農(nóng)業(yè)大學(xué) 農(nóng)學(xué)與生物技術(shù)學(xué)院, 云南 昆明 650201; 2.云南農(nóng)業(yè)大學(xué) 機電工程學(xué)院, 云南 昆明 650201)
0 引言
【研究意義】曼陀羅(DaturastramoniumL.)是茄科曼陀羅屬(Datura)的一年生草本植物,資源分布廣泛,其富含多種生物堿,在植株中現(xiàn)已發(fā)現(xiàn)超過70種托品烷類生物堿(Tropane alkaloids,TAs)[1-2]。在曼陀羅的TAs中,阿托品、莨菪堿、(±)山莨菪堿和東莨菪堿是主要成分[3-4],這些生物堿對作物蟲害防治有顯著效果[5],也是現(xiàn)代醫(yī)學(xué)廣泛應(yīng)用的抗菌[6]、抗癌[7-8]、消炎、鎮(zhèn)痛[9-10]等有效藥用成分,對曼陀羅藥用生物堿的積累特征展開研究,為獲取大量TAs進行開發(fā)利用具有重要的意義。【前人研究進展】TAs的生物合成以鳥氨酸、精氨酸和苯丙氨酸為起點[11-12],經(jīng)多步酶促反應(yīng)最后合成莨菪堿(Hyoscyamine)和阿托品(Atropine)[12],莨菪堿和阿托品的藥物活性類似,但莨菪堿的作用強度強于阿托品[5,13]。莨菪堿可經(jīng)H6H(Hyoscyamine 6 beta-hydroxylase)酶羥基化為山莨菪堿(Anisodamine),山莨菪堿為手性分子,根據(jù)其旋光性分為左旋山莨菪堿(-山莨菪堿)和右旋山莨菪堿(+山莨菪堿),兩者除旋光性不同外,藥理性質(zhì)和藥物活性強度基本一致[14]。(±)山莨菪堿可在H6H的催化作用下,繼續(xù)羥基化形成東莨菪堿(Scopolamine)[15]。東莨菪堿被認為是TAs代謝途徑中的最后一個生物堿,也是曼陀羅所有TAs中活性最強和藥用價值最高的生物堿[16]。東莨菪堿在植株中含量很低,通常僅有1~1 000 μg/g[17-18]。已有研究表明,曼陀羅的TAs主要在根中合成,由維管組織運輸至植株地上部,總生物堿在葉和種子中含量較高,莖中最低[4]。嫁接是果木、花卉和蔬菜品種改良常用的手段,通過利用接穗和砧木的各自優(yōu)勢,將優(yōu)良性狀集于一體[19-20]。嫁接技術(shù)在園藝作物的抗旱、抗?jié)澈涂果}堿性狀的改良中有重要作用[21-22],也是煙葉品質(zhì)改良的有效方法[23]。大量研究表明,砧木和接穗的營養(yǎng)物質(zhì)和遺傳信息存在相互交流和轉(zhuǎn)運[24],如以煙堿含量低,且鉀素利用率高的煙草品種為砧木,可提高煙草接穗的鉀素含量,并降低煙堿含量[25];利用不同辣椒品種進行嫁接,能夠提高接穗辣椒的香草醛、水楊酸和木質(zhì)素含量[26];楊懷亮等[27]發(fā)現(xiàn),與自根嫁接的黃瓜相比,雙砧木嫁接(將1個接穗嫁接到2株砧木上)可顯著提高黃瓜游離氨基酸、類黃酮和單寧含量。【研究切入點】目前TAs的主要獲取方式有TAs資源植物直接提取,基因工程生物合成和化學(xué)合成,但成本較高,且不易獲取。為滿足醫(yī)療和農(nóng)業(yè)生產(chǎn)對TAs生物堿的需求,有必要對曼陀羅TAs的合成和積累特征開展研究。【擬解決的關(guān)鍵問題】為研究曼陀羅TAs的合成和積累規(guī)律,將曼陀羅與馬鈴薯進行交互嫁接,分析不同砧木和接穗中TAs含量,探究TAs在不同器官中的積累特性。同時重點關(guān)注嫁接后馬鈴薯塊莖中是否能夠富集TAs,探索利用塊莖的高生物量特性來富集曼陀羅TAs的可能性,以期為進一步拓寬TAs的研究方法和制取方法提供參考。
1 材料與方法
1.1 試驗材料
馬鈴薯品系滇薯1418(D1418)種薯、曼陀羅種子均由云南農(nóng)業(yè)大學(xué)薯類作物研究所繁育保存。
1.2 試驗方法
1.2.1 馬鈴薯和曼陀羅嫁接及取材方法 各植物材料種植于云南農(nóng)業(yè)大學(xué)農(nóng)場溫室大棚。將曼陀羅種子播種,待幼苗出現(xiàn)第4片真葉后用于嫁接。將D1418種薯置于基質(zhì)中催芽直至出苗,當幼苗長出3~4片葉時,從種薯上連根掰開后分株移栽,栽約20 d后選擇生長一致的馬鈴薯幼苗用于嫁接。
兩材料交互分別作砧木和接穗,即嫁接時以馬鈴薯幼苗為砧木,曼陀羅為接穗的嫁接組合(曼陀羅/D1418);以曼陀羅為砧木,馬鈴薯為接穗的嫁接組合(D1418/曼陀羅)。嫁接幼苗成活后,對嫁接組合D1418/曼陀羅進行培土,用土將接穗馬鈴薯嫁接傷口上部的2~5個節(jié)充分覆蓋,以促進馬鈴薯莖基部葉腋處形成塊莖。嫁接50 d后對曼陀羅/D1418嫁接組合的曼陀羅莖、葉、果和D1418的塊莖進行取材;對D1418/曼陀羅嫁接組合的馬鈴薯莖、葉、葉腋處塊莖和曼陀羅的根進行取材,每個樣品3個生物學(xué)重復(fù)。將所取樣品液氮速凍后,于-80℃保存。
1.2.2 用于TAs測定的植物樣品前處理 在液氮中將各植物組織研磨成細粉,分別精確稱取研磨后的樣品各1.000 g置于具塞錐形瓶中。加入濃氨水-乙醇-乙醚(4︰5︰10,V/V)溶液5 mL,振蕩混勻,并靜置過夜。隨后,進行超聲破碎及提取(頻率:40 kHz;功率:250 W;溫度:25~30℃)2次,每次30 min。將提取液進行5 000 r/min離心10 min,吸取上清液至干凈試管中,于60℃水浴揮干溶劑。試管底部殘渣用0.5 mol/L硫酸1 mL復(fù)溶,再加入蒸餾水5 mL充分混勻。將上述獲得的溶液轉(zhuǎn)移到分液漏斗中,用氯仿10 mL萃取3次,合并上清液后水浴揮干。揮干后的殘渣用甲醇定容至10 mL,并振蕩混勻,取上清液1 mL所得溶液經(jīng)0.22 μm微孔濾膜過濾后備用。
1.2.3 TAs標準品制備 精確稱取適量硫酸阿托品、莨菪堿、(-)山莨菪堿、(+)山莨菪堿和氫溴酸東莨菪堿標準品,用色譜級甲醇溶解,并定容至5 mL,分別配制成1.0 mg/mL的標準品溶液。將硫酸阿托品稀釋成0.11 μg/mL、0.22 μg/mL、1.1 μg/mL、2.2 μg/mL、11 μg/mL、22 μg/mL;將莨菪堿標準品稀釋成0.148 μg/mL、0.296 μg/mL、1.48 μg/mL、2.96 μg/mL、14.8 μg/mL、29.6 μg/mL;將(-)山莨菪堿和(+)山莨菪堿分別稀釋成0.15 μg/mL、0.30 μg/mL、1.5 μg/mL、3.0 μg/mL、15 μg/mL、30 μg/mL;將氫溴酸東莨菪堿稀釋成0.1 μg/mL、0.2 μg/mL、1.0 μg/mL、2.0 μg/mL、10 μg/mL、20 μg/mL。將上述標準品溶液經(jīng)0.22 μm微孔濾膜過濾,備用。因(-)山莨菪堿和(+)山莨菪堿僅旋光性不同,作用性質(zhì)和強度相似,在計算山莨菪堿含量時,將二者合并計算表示為(±)山莨菪堿。
1.2.4 TAs的HPLC測定 用高效液相色譜儀(安捷倫1200)進行TAs測定。色譜柱為250 mm×4.6 mm×5 μm VWD C18,柱溫為25℃,檢測波長210 nm。洗脫方式為等度洗脫,流動相為乙腈︰0.085%甲酸=80︰20(V/V),流速1 mL/min,樣品進樣量為10 μL。
1.2.5 托品烷類生物堿測定方法的建立 利用等度洗脫法對5個托品烷類生物堿進行分離,其中,莨菪堿和阿托品出峰時間基本一致,混合測定不能將兩者進行區(qū)分,因此將(-)山莨菪堿、(+)山莨菪堿、東莨菪堿和阿托品標品混合測定,莨菪堿標品單獨測定。HPLC不能直接定量,需使用回歸分析,因此測定不同濃度的4種TAs混合標品和莨菪堿標品峰面積,并繪制各生物堿的標準曲線(即回歸分析得到標準曲線,運用標準曲線倒推生物堿含量)。為驗證色譜方法的正確性,對交互嫁接后馬鈴薯和曼陀羅各器官中的TAs進行提取,樣品的出峰時間與標準品出峰時間基本一致,則色譜方法正確,反之亦然;為保持試驗條件的一致性,根據(jù)確定的標準品測定方法測定5種托品烷類生物堿含量。
1.2.6 托品烷類生物堿含量的測定 以曼陀羅為接穗,D1418為砧木進行嫁接的測定馬鈴薯塊莖、曼陀羅莖、葉和果實中5種托品烷類生物堿的含量。以D1418為接穗,曼陀羅為砧木進行嫁接的測定馬鈴薯莖、葉、葉腋處塊莖及曼陀羅根中的5種托品烷類生物堿的含量。
2 結(jié)果與分析
2.1 5個托品烷類生物堿的測定方法建立
對5個托品烷類生物堿進行分離測定結(jié)果(圖 1)顯示,4種混合生物堿標品可形成4個獨立峰,均有較好的分離度,(-)山莨菪堿、(+)山莨菪堿、東莨菪堿和阿托品的保留時間分別為5.888 min、6.886 min、7.906 min、10.968 min。莨菪堿標品的保留時間為10.868 min。
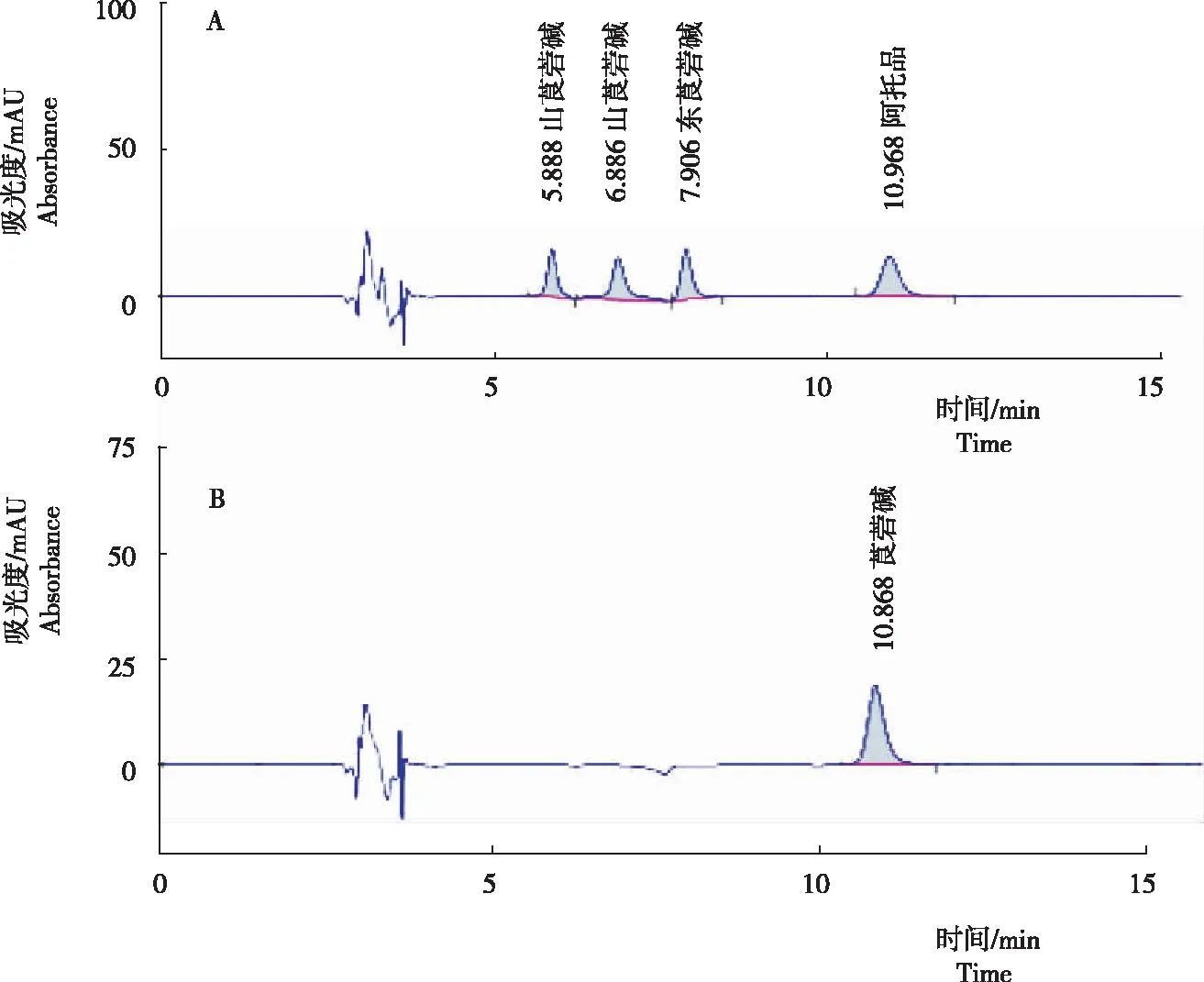
注:A為阿托品、(-)山莨菪堿、(+)山莨菪堿和東莨菪堿混合標品色譜峰圖;B為莨菪堿標品色譜峰圖。
從圖2看出,各托品烷類生物堿標準品的濃度和峰面積均有較好的線性關(guān)系,標準曲線R2均大于0.999,標準曲線線性擬合度較好,可用于后續(xù)樣品測定的計算。
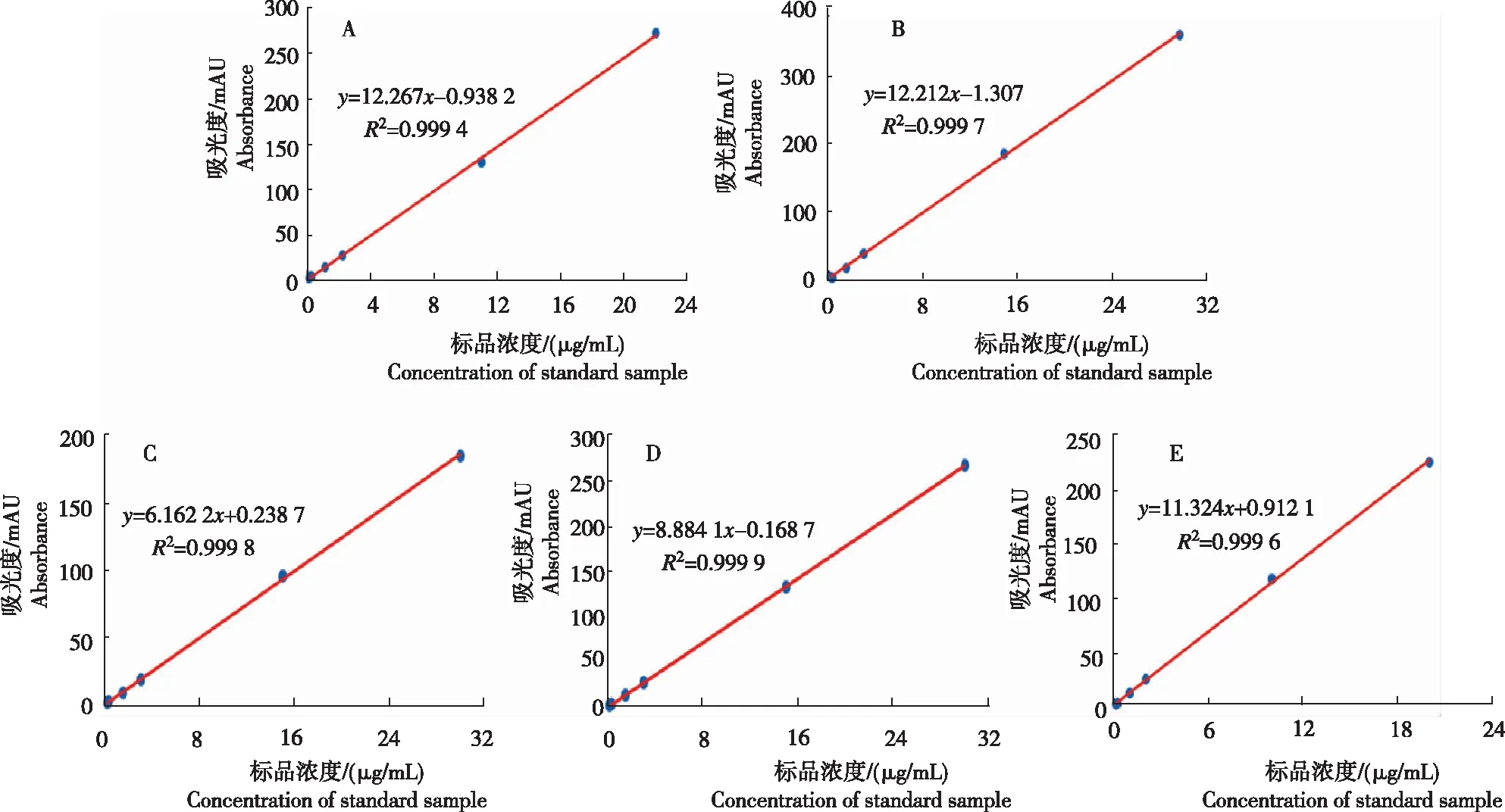
注:A為阿托品,B為莨菪堿,C為(-)山莨菪堿,D為(+)山莨菪堿,E為東莨菪堿。
從圖3看出,以曼陀羅果實為例,樣品提取和HPLC分離后,能夠形成(-)山莨菪堿、(+)山莨菪堿、東莨菪堿和阿托品的特征峰,各成分的保留時間分別為5.817 min、6.771 min、7.751 min和10.791 min,均與混合標準品中各成分的保留時間相符。在曼陀羅果實樣品中,莨菪堿能夠形成較好的特征峰,保留時間為10.876 min。認為,該方法穩(wěn)定可靠;在上述色譜條件下,使用HPLC可進行樣品中5種TAs的測定。
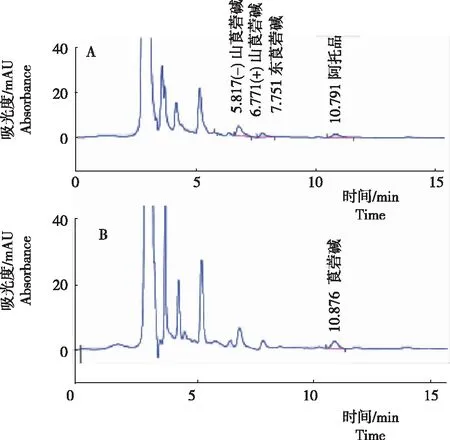
注:A為阿托品、(-)山莨菪堿、(+)山莨菪堿和東莨菪堿色譜峰圖;B為莨菪堿色譜峰圖。
2.2 曼陀羅/D1418嫁接后各器官托品烷類生物堿含量
對以曼陀羅為接穗,D1418為砧木進行嫁接的曼陀羅莖、葉和果實及馬鈴薯塊莖中5種藥用生物堿含量的測定結(jié)果(表1)顯示,在砧木和接穗不同器官中,均可檢測到5種托品烷類生物堿。曼陀羅接穗各器官中,TAs生物堿含量在果實中最高,為121.77 μg/g;葉片中最低,為35.61 μg/g。曼陀羅接穗各器官中,均是(±)山莨菪堿含量最高,為29.15~87.98 μg/g;東莨菪堿含量最低,為1.02~9.07 μg/g。在馬鈴薯砧木所形成的塊莖中,5種生物堿含量總和僅為8.16 μg/g,其中莨菪堿含量最高,為2.87 μg/g;阿托品含量最低,為1.50 μg/g。曼陀羅莖和馬鈴薯塊莖的(±)山莨菪堿含量間差異顯著。表明,曼陀羅果實中托品烷類生物堿含量最高;以曼陀羅為接穗,馬鈴薯為砧木所形成的塊莖中能夠富集一定量的托品烷類生物堿,說明托品烷類生物堿能夠在維管組織中從上至下運輸。
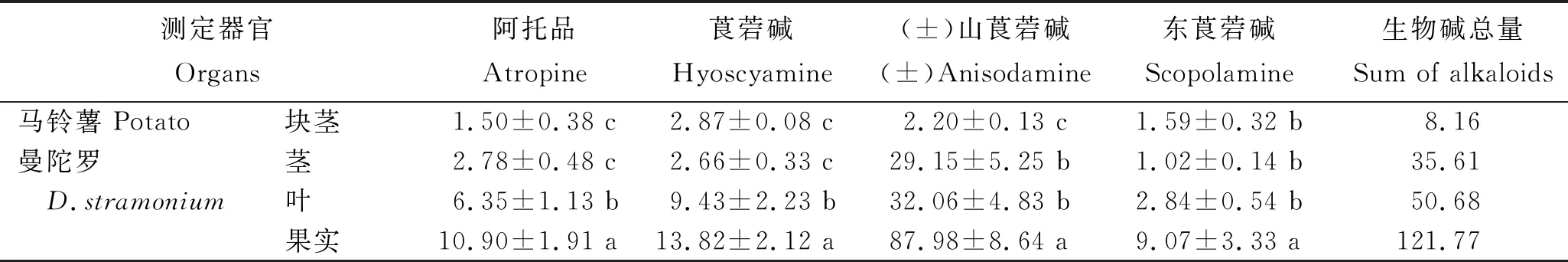
表1 曼陀羅/D1418嫁接后不同器官中5種托品烷類生物堿含量
2.3 D1418/曼陀羅嫁接后各器官托品烷類生物堿含量
對以D1418為接穗,曼陀羅為砧木進行嫁接后馬鈴薯的莖、葉、葉腋處塊莖及曼陀羅根中5種托品烷類生物堿含量的測定結(jié)果(表2)顯示,砧木和接穗器官中均存在5種托品烷類生物堿。生物堿總量以馬鈴薯葉片中的最高,為214.23 μg/g;曼陀羅砧木的根中較低,為34.13 μg/g;馬鈴薯葉腋處塊莖的最低,為13.22 μg/g。4個器官中均是莨菪堿含量最高,阿托品含量其次,(±)山莨菪堿和東莨菪堿含量較低。表明,曼陀羅根系中托品烷類生物堿能夠在維管組織中從下至上運輸,并在馬鈴薯接穗莖葉中富集,也能在葉腋處的氣生薯中富集。
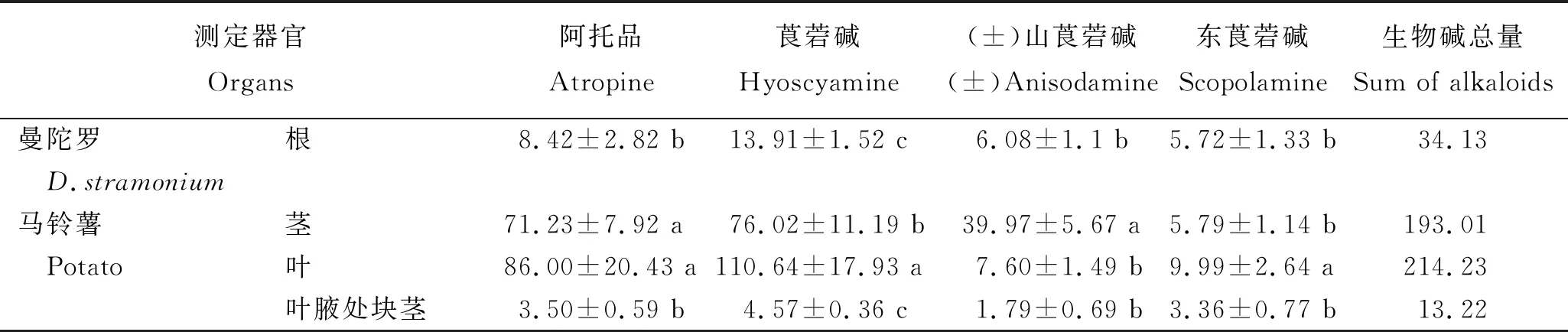
表2 D1418/曼陀羅嫁接后不同器官中5種托品烷類生物堿含量
3 討論
3.1 交互嫁接后曼陀羅不同器官中托品烷類生物堿的積累規(guī)律
曼陀羅中主要的生物堿,如阿托品、莨菪堿、山莨菪堿和東莨菪堿等均在根系中進行生物合成[28-31],并通過維管組織運輸?shù)降厣喜糠帧1418/曼陀羅嫁接組合中,馬鈴薯接穗莖葉的托品烷生物堿含量高于曼陀羅/D1418嫁接中的曼陀羅莖葉。曼陀羅砧木的根系能夠不斷合成TAs,是生物堿合成的“源”;而曼陀羅砧木的根系所產(chǎn)生的TAs,從維管組織大量轉(zhuǎn)運到地上部馬鈴薯接穗莖葉中,使馬鈴薯接穗成為“庫”;當以馬鈴薯為砧木時,失去了TAs合成的“源”,因此曼陀羅接穗中的生物堿含量低于馬鈴薯接穗。在曼陀羅/D1418嫁接組合中,馬鈴薯砧木的塊莖中也積累了一定量的TAs,說明TAs在曼陀羅莖中能夠自上而下運輸。
在TAs生物合成途徑的下游,莨菪堿和阿托品是(±)山莨菪堿的合成前體,而(±)山莨菪堿是東莨菪堿的合成前體,這2步酶促反應(yīng)都受到同一限速酶H6H的催化[15,32]。因此,在曼陀羅中莨菪堿和阿托品的含量高于東莨菪堿。曼陀羅各組織器官中的東莨菪堿含量均較低,該現(xiàn)象與TAs生物合成過程相符。
曼陀羅根系中莨菪堿是5種生物堿的主要組成成分,而在曼陀羅地上部組織中(±)山莨菪堿是主要的組成成分,說明曼陀羅莖葉和果實可能特異性地積累(±)山莨菪堿,同時在莖葉和果實中也可能自主合成(±)山莨菪堿。
3.2 馬鈴薯接穗對曼陀羅中托品烷類生物堿的積累作用
通過嫁接能夠使砧木和接穗之間的營養(yǎng)物質(zhì)產(chǎn)生相互交流[24]。馬鈴薯和曼陀羅都是茄科作物,馬鈴薯的主要生物堿是糖苷生物堿[33],曼陀羅的主要生物堿是托品烷類生物堿[2,34]。D1418/曼陀羅嫁接后,馬鈴薯莖葉中也能夠積累大量的TAs,說明馬鈴薯維管組織也能夠轉(zhuǎn)運和富集曼陀羅的TAs。同時,馬鈴薯接穗和曼陀羅接穗莖葉中的托品烷類生物堿主要組成成分不同,說明馬鈴薯維管組織積累托品烷類生物堿的具體機制可能與曼陀羅不同。馬鈴薯和曼陀羅交互嫁接中,在馬鈴薯的地下塊莖及地上部葉腋氣生薯中均檢測到曼陀羅的TAs,該結(jié)果為利用馬鈴薯塊莖來特異性積累TAs奠定了基礎(chǔ)。另外,在馬鈴薯地上或地下塊莖中莨菪堿均是含量最高的TAs,說明塊莖組織可能有偏好性地積累托品烷類生物堿。
4 結(jié)論
托品烷類生物堿通過維管組織既能從上至下運輸,也能從下至上運輸。以曼陀羅為接穗,馬鈴薯為砧木所形成的塊莖中能夠富集一定量的托品烷類生物堿;以曼陀羅為砧木,馬鈴薯為接穗時,托品烷類生物堿能在馬鈴薯接穗莖葉中富集,同時也能在馬鈴薯葉腋處的氣生薯中富集。
