生物絮團系統中不同養殖比例對松浦鏡鯉營養及生理指標的影響
2022-06-06 13:32:50張美彥張效平李建光商寶娣
西南農業學報 2022年5期
關鍵詞:生物
張美彥,楊 星,張效平,趙 鳳,李建光,商寶娣
(1.貴州省農業科學院水產研究所,貴陽 550025;2.貴州省特種水產工程技術中心,貴陽 550025)
【研究意義】現階段的池塘養殖為達到理想的產量和經濟效益,投入大量肥料、飼料以及藥物,加之池塘內養殖魚類代謝物沉積,導致池塘內源性污染較為嚴重,養殖環境惡化。因此,如何降低池塘養殖水體的自身污染,減少養殖廢水的排出,提高飼料利用效率成為池塘養殖技術中有待突破的一個關鍵技術問題。【前人研究進展】生物絮團技術(Biofloc Technology,BFT)是在養殖水體零換水基礎上,通過人為添加有機碳源調節水體碳氮比(C/N),提高養殖水體中異養細菌的數量,利用微生物同化無機氮,將水體中的氨氮等養殖代謝產物轉化成細菌自身成分,通過細菌絮凝成顆粒物質被養殖對象所攝食,起到調控水質、促進營養物質循環、降低飼料系數、提高養殖對象成活率的作用[1]。傳統養殖技術中,鯉魚與鰱鳙比例一般為80∶20,在生物絮團系統中,營養物質發生變化,導致水體中微生物及藻類發生變化,會影響到主養魚與鰱鳙魚的最適投放比例。以生物絮團技術為基礎的水產養殖模式在對蝦、羅非魚養殖系統中應用較多,主要集中在對水體中C/N、養殖對象的生長、水質變化、微生物群落等[2-6]。【本研究切入點】目前,對生物絮團系統中魚類投放比例研究較少,距離應用到實際生產還需較長時間[7]。【擬解決的關鍵問題】對喀斯特山區池塘生物絮團系統中魚類合理投放比例進行研究,為建立貴州省節能、節水、高效的池塘集約化養殖模式提供理論基礎與數據支撐。
1 材料與方法
1.1 材料
試驗魚苗由綏陽縣風華鎮魚子孔養殖場提供,品種及規格分別為,松浦鏡鯉約110 g/尾,鰱魚約70 g/尾,鳙魚約60 g/尾,草魚約150 g/尾。全價配合魚飼料為貴陽金滿船飼料有限公司生產,主要營養成分含量:粗蛋白質≥32%,粗脂肪≥2%,粗灰分≤16%,粗纖維≤5%,食鹽≤4%,鈣≤5%,總磷≥0.8%,水份≤13.5%,賴氨酸≥1.9%。新型羅茨式增氧曝氣風機功率2.2 kw ,江陰江達機械裝備有限公司生產。噴水式增氧機功率1.5 kw,無錫漁愉魚科技有限公司生產。
1.2 試驗方法
1.2.1 池塘生物絮團系統構建 試驗池塘位于貴州省遵義市綏陽縣風華鎮魚子孔養殖場。試驗池塘養殖條件基本一致,面積均為867 m2,池塘水深約1.6 m,養殖前期根據水體中的氨氮含量,按照C/N為20向池塘中定期潑灑糖蜜作為外源性添加碳源,碳源添加量根據水體氨氮值計算。
△CH=20×H×S×CNH3-N
式中,△CH為碳源量(kg),H為池塘水深(m),S為池塘養殖面積(m2),CNH3-N為池塘水體測定的氨氮濃度(mg/L)。
1.2.2 試驗設計 主養魚為松浦鏡鯉,與鰱魚、鳙魚和草魚混養。松浦鏡鯉與其他魚類的養殖比例分別設為70∶30、75∶25和80∶20,每個養殖比例為1處理,共3個處理,每處理1個池塘,養魚1200 尾,魚苗投放量見表1。魚苗于2019年5月9日放入池塘,2019年11月14日結束養殖,養殖周期共190 d。
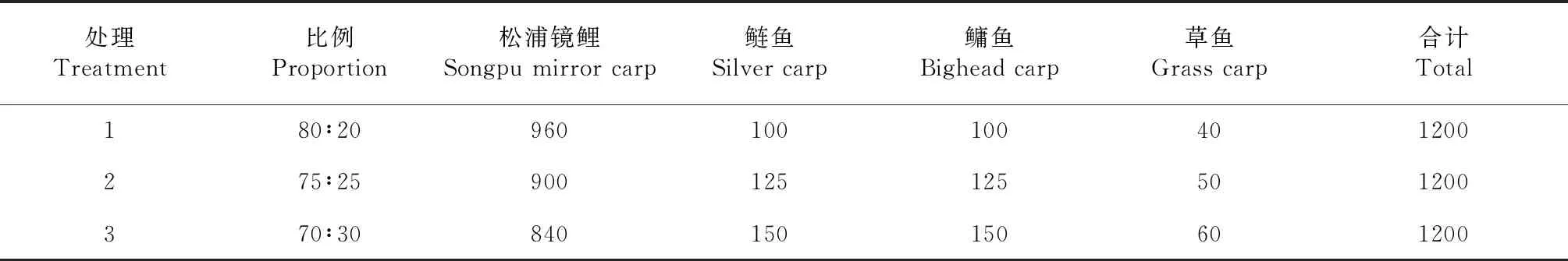
表1 生物絮團系統中不同養殖比例魚苗的投放量
1.2.3 飼養管理 試驗全程投喂全價配合飼料,根據魚體大小選擇對應粒徑飼料,按照“四定原則”進行投喂,每日3次。養殖期間不換水,每20 d向池塘內補充1次水至一定水位彌補池塘內水分蒸發量。池塘用新型羅茨式增氧曝氣風機和噴水式增氧機進行增氧。
1.3 測定指標及方法
1.3.1 樣品采集 養殖試驗結束后,每個池塘取4尾魚,于尾靜脈取血,用于測定血清生化指標;取前腸、中腸、后腸,用于測定消化酶活性;將每尾魚取肌肉,用于肌肉成分分析;取肝臟,用于測定其肝臟抗氧化酶活性。
1.3.2 肌肉成分測定 肌肉初水分測定采用105 ℃干燥恒重法(GB/T 5009.3—2003),粗蛋白含量測定采用凱氏定氮法(GB/T 5009. 5—2003)測定,粗脂肪含量采用索氏抽提法(GB/T 5009.6—2003)測定,灰分含量采用高溫灼燒法(GB/T 5009.4—2003)測定。
1.3.3 消化酶活性測定 蛋白酶用福林—酚法測定,蛋白酶活性測定中,在37 ℃條件下,每1 min水解酪素產生1 μg酪氨酸為1個酶活單位。淀粉酶和脂肪酶均使用南京建成生物工程研究所生產的試劑盒測定。脂肪酶活性測定中,在37 ℃條件下,每1 g組織蛋白在本反應體系中與底物反應1 min,每消耗1 μmol底物為1個酶活單位。淀粉酶測定中,在37 ℃下,組織中每1 mg蛋白與底物作用30 min,水解10 mg淀粉定義為1個酶活單位。
1.3.4 抗氧化酶活性測定 抗氧化酶測定指標包括超氧化物歧化酶(SOD)和丙二醛(MDA)含量,均采用南京建成生物工程研究所生產的試劑盒測定。
1.4 數據統計分析
2 結果與分析
2.1 松浦鏡鯉的肌肉成分
從表2看出,生物絮團系統中不同養殖比例池塘松浦鏡鯉的肌肉營成分差異不顯著(P>0.05)。初水分和灰分含量均以處理3最高,處理1最低;粗蛋白含量以處理1最高,處理3最低;粗脂肪含量處理3最高,處理2最低。
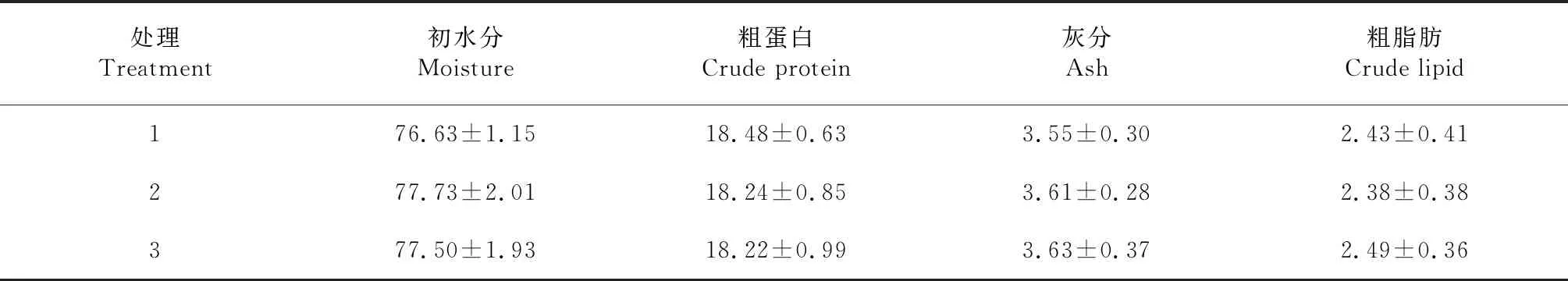
表2 生物絮團系統中不同養殖比例松浦鏡鯉的肌肉成分含量
2.2 松浦鏡鯉的消化酶活性
從表3看出,中腸淀粉酶活性高于前腸和后腸,前腸和后腸淀粉酶活性均以處理2最強,中腸淀粉酶活性以處理3最強,但各處理間差異均不顯著。前腸、中腸和后腸的蛋白酶活性均以處理2的最強,前腸和后腸蛋白酶活性各處理間差異不顯著,中腸處理2顯著高于處理3,與處理1間差異不顯著。前腸和中腸的脂肪酶活性均以處理2最高,后腸則以處理1最高,但各處理間差異不顯著。
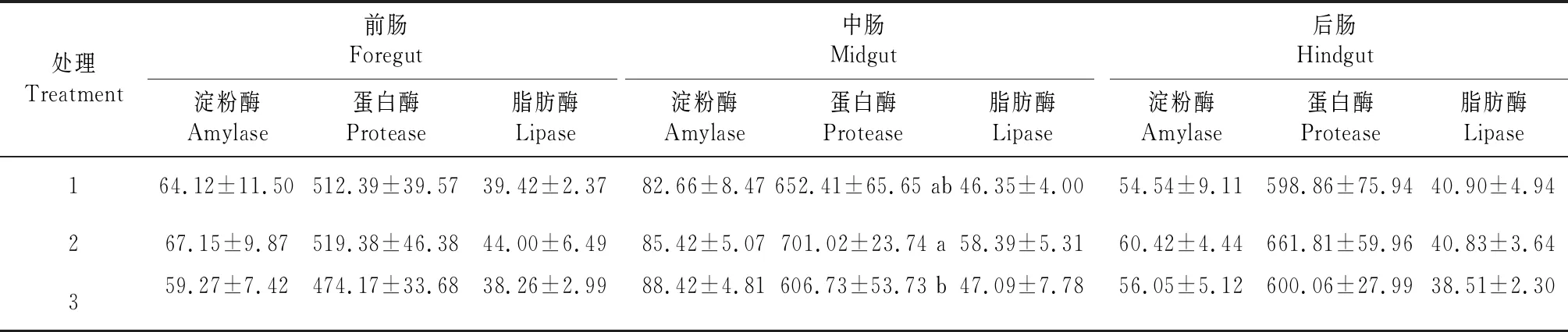
表3 生物絮團系統中不同養殖比例松浦鏡鯉腸道消化酶的活性
2.3 松浦鏡鯉的肝臟抗氧化指標
從表4可知,超氧化物歧化酶(SOD)活性和丙二醛(MDA)含量均以處理2最高,分別為89.70 U/mg和4.07 nmol/mg。SOD活性各處理間差異不顯著(P>0.05),MDA含量處理2顯著高于處理3,與處理1間差異不顯著。
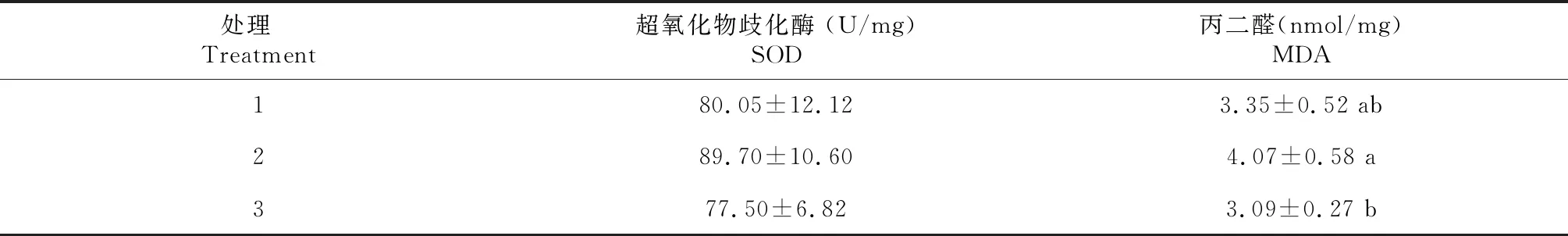
表4 生物絮團系統中不同養殖比例松浦鏡鯉的肝臟抗氧化指標
2.4 松浦鏡鯉的血清生理指標
從表5看出,總蛋白(TP)含量以處理2最高,處理3最低,各處理間差異不顯著(P>0.05)。白蛋白(ALB)含量以處理2顯著高于處理1和處理33(P<0.05)。球蛋白(GLB)含量處理2略高于處理3,處理1最低,各處理間差異不顯著(P>0.05)。堿性磷酸酶(AKP)含量以處理2最高,其次是處理3,處理1最低,但各處理間差異不顯著(P>0.05)。

表5 生物絮團系統中不同養殖比例松浦鏡鯉的血清生化指標
3 討 論
3.1 生物絮團對不同養殖比例池塘松浦鏡鯉肌肉營養成分的影響
試驗結果顯示,3組養殖比例條件下,松浦鏡鯉肌肉營養成分沒有顯著差異,說明生物絮團模式下對不同養殖比例主養鯉池塘中松浦鏡鯉的肌肉成分影響不顯著。葛海倫等[8]研究未發現羅氏沼蝦(Macrobrachiumrosenbergii)試驗組與對照組間存在顯著差異;Luo等[9]對羅非魚(Oreochromisniloticus)的研究中也未發現試驗組與對照組間存在顯著差異,這與本研究結果相同。Lzquierdo等[10]對南美白對蝦(litopenaeusvannamei)的研究及Long等[11]對羅非魚的研究發現,生物絮團模式下肌肉中粗脂肪含量顯著高于對照組,產生這種結果的原因可能是生物絮團中含有一定量(4.6%~8.3%)的脂肪[12],造成養殖動物攝食后體內粗脂肪含量升高;本研究結果表明,不同養殖比例松浦鏡鯉的肌肉成分差異不顯著,原因可能是試驗設置的養殖比例不足以對池塘內生物絮團的產生量造成顯著影響所致。
3.2 生物絮團對不同養殖比例池塘松浦鏡鯉消化酶活性的影響
消化酶與動物的生長發育密切相關,是反映機體營養生理的重要指標之一,消化酶活性直接影響到動物的消化能力,進而直接影響飼料利用效率。本試驗數據表明,添加生物絮團后,75∶25組松浦鏡鯉中腸蛋白酶活性顯著高于其他兩組。Farideh等[13]的研究中表明,生物絮團可顯著提高鯉(Cyprinuscarpio)蛋白酶、脂肪酶、淀粉酶活性。張明明等[14]對異育銀鯽(Carassiusauratusgibelio)的研究中發現,生物絮團可提高腸道脂肪酶、淀粉酶活性。孫盛明等[15]對團頭魴(Megalobramaamblycephala)的研究中也發現,生物絮團可提高團頭魴的蛋白酶和淀粉酶活性;李京昊等[16]在對克氏原螯蝦(Procambarusclarki)的研究中發現,添加生物絮團后可提高肝胰臟中的淀粉酶活性。葛海倫等[8]對羅氏沼蝦的研究發現,添加生物絮團后,腸、胃、胰中脂肪酶活性高于對照組。Najdegerami等[17]在研究鯉魚養殖水體中生物絮團對消化酶活性影響中發現,水體中一定含量的生物絮團能夠顯著提高蛋白酶的活性,本研究雖未對生物絮團中的消化酶活性進行數據分析,但松浦鏡鯉中腸蛋白酶活性的顯著升高很可能與絮團本身含有的消化酶有關。生物絮團中的異養細菌在生長代謝過程中能分泌蛋白酶、淀粉酶等胞外酶,絮團被養殖對象攝食后,這些胞外酶可在腸道中幫助分解蛋白質、碳水化合物等[18]。另外,生物絮團中也有可能存在一些未知的生長因子,刺激動物分泌消化酶或提高消化酶活性[19]。75∶25組生物絮團養殖模式下松浦鏡鯉中腸蛋白酶活性最高,表明在此養殖比例下,生物絮團中的異養細菌及其他生長因子較多,生物絮團更能有效發揮作用,從而提高養殖對象蛋白酶活性。
3.3 生物絮團對不同養殖比例池塘松浦鏡鯉抗氧化指標的影響
氧化與抗氧化處于動態平衡,是機體進行正常代謝的保障[20],機體在代謝過程中會產生少量活性氧物質,抗氧化酶系統能夠將活性氧物質清除,SOD是一種重要的抗氧化酶。MDA是機體脂質過氧化的重要產物,其含量多少會影響機體正常的物質代謝及能量代謝。肝臟SOD、MDA含量可反映機體抗氧化及脂質過氧化水平,MDA同時也可反映細胞氧化損傷程度[21]。在對細角濱對蝦的研究中也發現,生物絮團組繁殖期內的細角濱對蝦親蝦存活率更高,總抗氧化能力更高[22]。本試驗中,75∶25組丙二醛含量最低,可能是此養殖比例模式下,改變了松浦鏡鯉的抗自由基狀態,對氧化應激不敏感,增強其抗氧化能力;另外,養殖水體中含有適量微生物及其代謝產物,松浦鏡鯉可充分利用,作為免疫刺激源,達到較好的抗氧化狀態[18]。
3.4 生物絮團對不同養殖比例主養鯉池塘松浦鏡鯉血清生化指標的影響
血清生化指標可反映動物組織細胞通透性及新陳代謝機能,其中血清中的TP、ALB和球蛋白GLB可反映蛋白吸收和代謝狀況[23],在一定范圍內,血清蛋白含量和機體代謝水平、免疫能力、蛋白合成及氮沉積能力呈正相關[24-25]。血清中的AKP主要由骨細胞產生,經肝膽系統排泄。當體內AKP的生成過多或是排泄過程受阻時,血清中AKP活性發生變化。本試驗中,各組間AKP活性無顯著變化,可能試驗設置的3中不同養殖比例,對水體中的藻類及微生物的影響不足以引起AKP的顯著變化。ALB含量差異較為顯著,75∶25組含量最高,表明此組松浦鏡鯉蛋白吸收、代謝情況及免疫能力高于其他組。在對刺身的研究中也發現,生物絮團系統中,可提高幼參的免疫力[26]。Su等[27]在養殖系統中發現生物絮團對南美白對蝦仔蝦生長和免疫相關的基因表達有積極作用。李朝兵等[28]對鳙魚(Aristichthysnobilis)的研究表明,適量生物絮團可通過換水量的較少來減輕外界病原物質侵擾,從而增強機體免疫力。在對超級鯉的研究中發現,生物絮團系統中有益細菌大量繁殖,進入對蝦腸道,通過機體代謝物或表面抗原可刺激免疫系統,從而提高對蝦免疫力[29]。此試驗中免疫能力的提高也可能和免疫相關基因的表達有關,具體的作用機理還有待深入研究。
4 結 論
在生物絮團系統中,松浦鏡鯉與鰱鳙草放養比例為75∶25組松浦鏡鯉消化酶活性較高,抗氧化能力較強,免疫能力更高。本試驗條件下,生物絮團系統中最適放養比例為75∶25。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
小學科學(學生版)(2018年3期)2018-04-18 12:34:19
知識經濟·中國直銷(2017年10期)2017-11-07 02:39:52
知識經濟·中國直銷(2017年3期)2017-04-16 03:08:01
文學少年(有聲彩繪)(2017年2期)2017-03-06 08:10:54
