湖北省部分規模豬場豬繁殖與呼吸綜合征流行病學調查
2022-06-07 05:51:44王妍趙潤澤王鈺李桐陳少賢張子微楊小林郭利偉劉國平
中國動物檢疫 2022年6期
陳 騰,王妍,趙潤澤,王鈺,李桐,陳少賢,張子微,楊小林,2,郭利偉,2,劉國平,2
(1.長江大學動物科學學院,湖北荊州 434025;2.長江大學交叉生物技術產業研究中心,湖北荊州 434025)
豬繁殖與呼吸綜合征(porcine reproductive and respiratory syndrome,PRRS)是一種以繁殖障礙和呼吸系統癥狀為特征的接觸性豬病毒傳染病,是制約生豬養殖業發展的重要疾病之一。其病原體豬繁殖與呼吸綜合征病毒(porcine reproductive and respiratory syndrome,PRRSV)是動脈炎病毒科動脈炎病毒屬的成員,是一種有囊膜的單股正鏈RNA 病毒[1]。研究[2]表明:各生長階段豬只均易感染PRRSV,傳播途徑主要有水平傳播和垂直傳播,其中水平傳播包括與患病豬接觸、空氣傳播以及精液傳播等,垂直傳播主要經胎盤傳播。目前在國內流行的PRRSV 按致病力不同,大致可以分為經典毒株、高致病性變異毒株和類NADC30 毒株。1995 年國內首次發現疑似PRRS 豬,1996 年從豬陽性血清中成功分離出PRRSV,從而證實了國內有PRRS 流行[3]。1995—2006 年,PRRS 表現經典臨床癥狀,以經典毒株(CH1a 毒株)流行為主。經典毒株主要導致母豬繁殖障礙、仔豬呼吸系統疾病,以及育肥豬呼吸道疾病、生長發育緩慢,還會出現繼發性疾病,導致哺乳豬、保育豬死亡率上升。母豬群暴發的經典PRRS,一般會在一個半月內趨于穩定,但潛伏期較長。而高致病性PRRS從2006 年開始在我國流行,其病原是毒力更強的PRRSV 變異株(JXA1 毒株),相比較于經典毒株,高致病性變異毒株致病力更強,傳播速度更快,除了引起經典毒株的癥狀外,對神經系統、免疫系統、消化系統也有較強的致病性,仔豬發病率甚至可高達100%,死亡率達50%,母豬流產率超過30%[4-5],給國內大部分地區的規模化豬養殖場造成了重大經濟損失。從2012 年開始,陸續有PRRSV 新毒株在國內出現的報道,其與NADC30 毒株同源關系較近,因此將這類病毒統稱為類NADC30 毒株,但不同NADC30 毒株之間的致病力呈現出一定的差異[6-8]。近年來有文獻[8-9]報道,類NADC30 毒株與其他毒株發生了重組變異,使PRRSV 基因型更加復雜,這給我國PRRS 防控帶來了更大挑戰。
為探究PRRSV 在規模豬場的流行特征,2021年從湖北省7 個規模豬場疑似感染PRRSV 豬群以及所有種公豬群中收集樣品進行病原學檢測,并從養殖類型、生長階段、季節及感染毒株等方面進行統計分析,以期為今后的PRRS 防控提供依據。
1 材料和方法
1.1 病料采集
2021 年,從湖北省7 個規模化養殖場(1 個存欄1 000~2 000 頭的核心育種場、1 個存欄>5 000 頭的商品豬擴繁場和5 個存欄>5 000 頭的育肥場),對臨床上出現呼吸和繁殖障礙癥狀以及耳朵發紫和有繁殖障礙史的豬群進行采樣檢測,對核心育種場的種公豬進行普檢,共采集3 324 份臨床樣品。將樣品置于冰上運輸至實驗室,并于24 h內完成后續檢測。
1.2 樣品預處理及病毒RNA 提取
樣品預處理方法參照文獻[10],預處理樣品用于后續病毒RNA 提取。樣品病毒RNA 提取,采用Axygen 體液病毒核酸提取試劑盒,具體步驟參照試劑盒說明書進行。用Nanodrop 1 000 檢測所提取RNA 的濃度和純度,將RNA 于-20 ℃保存備用。
1.3 病原學檢測
采用三重巢式RT-PCR 法[11],對樣品進行PRRSV 病原學檢測(此方法可同時檢測經典毒株、高致病性變異毒株和類NADC30 毒株)。引物序列如表1 所示。

表1 引物序列
1.4 數據統計與分析
對不同類型養殖場、不同年齡段、不同季節及混合感染的檢測結果,采用SPSS 24.0 卡方檢驗進行差異性分析,P≥0.05 為差異不顯著,P<0.05 為差異顯著。通過Origin 2019 軟件,對檢測樣品的PRRSV 陽性檢出率進行處理分析。從檢測出的不同毒株陽性產物中各隨機選取3 份(共9 份陽性產物),送至生工生物工程(上海)股份有限公司測序;將測序序列通過NCBI 進行BLAST 比對,并利用MEGA7.0 軟件中Neighbor Joining 法,繪制系統發育進化樹。
2 結果與分析
2.1 不同養殖類型
不同類型養殖場檢測結果(表2)顯示:共檢出PRRSV 陽性樣品數323 份,平均陽性檢出率為9.72%(323/3 324),其中核心育種場陽性檢出率為2.66%(20/752),商品豬擴繁場為11.82%(169/1 430),育肥場為11.73%(134/1 142);核心育種場陽性檢出率顯著低于擴繁場和育肥場(P<0.05),而商品豬擴繁場與育肥場陽性檢出率差異不顯著(P≥0.05)。結果表明,與商品豬擴繁場和育肥場相比,核心育種場PRRSV 感染程度較輕。

表2 不同類型養殖場PRRSV 檢測結果
2.2 不同生長階段
不同生長階段豬群檢測結果(表3)顯示:種公豬未檢出陽性,陽性檢出率為0(0/183),母豬陽性檢出率為3.24%(20/618),哺乳仔豬為5.26%(30/570),保育豬為17.23%(152/882),生長育肥豬為11.30%(121/1 071);不同生長階段豬群的PRRSV 陽性檢出率差異顯著(P<0.05)。結果表明,不同生長階段豬群的PRRSV感染程度差異較大,保育豬感染最嚴重,其余依次是生長育肥豬、哺乳仔豬和母豬,而種公豬經普檢未發現感染。
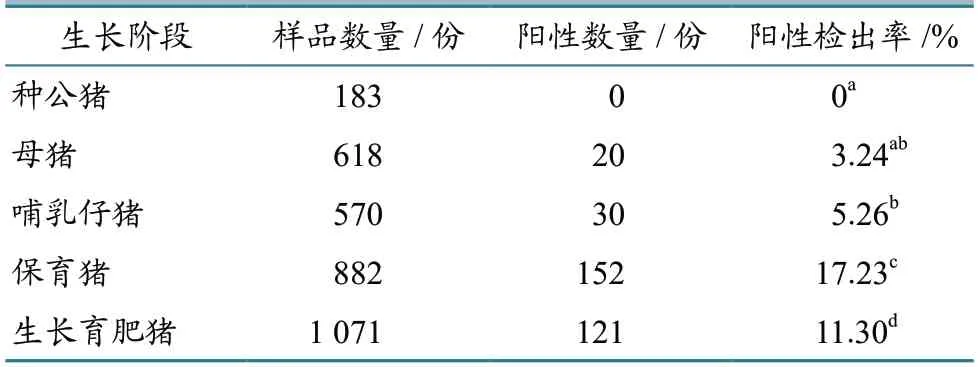
表3 不同生長階段豬群PRRSV 檢測結果
2.3 不同季節
不同季節豬群檢測結果(表4)顯示:春季(3—5 月)的PRRSV 陽性檢出率為18.16%(148/815),夏季(6—8 月)為6.94%(60/864),秋季(9—11 月)為6.10%(34/557),冬季(12月—次年2 月)為7.44%(81/1 088),春季陽性檢出率顯著高于其他季節(P<0.05),而夏秋冬三季間的陽性檢出率差異不顯著(P≥0.05)。結果表明,春季是PRRSV 感染的高峰期,夏秋兩季豬群感染程度明顯下降,冬季開始反彈。

表4 不同季節豬群PRRSV 檢測結果
2.4 不同毒株
陽性樣品中不同PRRSV 毒株檢出結果(表5)顯示:323 份PRRSV 陽性樣品中,類NADC30 毒株檢出占比最高,為49.54%(160/323),其余依次是高致病性變異毒株(29.10%,94/323)、經典毒株(21.36%,69/323),未發現多重毒株混合感染情況。類NADC30 毒株檢出占比顯著高于其他毒株(P<0.05),而高致病性變異毒株與經典毒株檢出占比差異不顯著(P≥0.05)。

表5 陽性樣品中不同PRRSV 毒株檢出情況
選取9 份陽性產物測序并建立系統發育進化樹。結果(圖1)顯示:9 條序列分別屬于PRRSV經典毒株、高致病性變異毒株與類NADC30 毒株,其中3 條序列(HB-01、HB-02 及HB-05)與以VR2332 為代表的經典毒株屬于同一進化分支,3 條序列(HB-03、HB-08 及HB-09)與以JXA1為代表的高致病性變異毒株屬于同一進化分支,剩余3條序列(HB-04、HB-06 及HB-07)與 以NADC30 為代表的類NADC30 毒株屬于同一進化分支。
3 討論
本研究對2021 年湖北省7 個規模豬場的PRRSV 感染情況進行調查,發現平均陽性檢出率為9.72%(323/3 324)。趙永祥[12]2013—2014 年對四川省部分地區進行PRRSV 流行病學調查,發現平均陽性率為38.76%(69/178);劉曉東[13]2015—2017 年對國內部分地區進行PRRSV 流行病學調查,發現平均陽性率為31.84%(1138/3 574);李焱[14]2018—2020 年對山東省進行PRRSV 流行病學調查,發現平均陽性率為16.57%(355/2 143)。從上述調查結果可以看出,2018 年后國內PRRSV的感染程度呈下降趨勢,與本研究以及牛婷婷等[15]的研究結果一致。這可能是自2018 年非洲豬瘟在國內暴發以來,國內許多規模豬場配套了更完善的生物安全體系,加上全國限制跨區域調豬,這些措施對PRRS 防控也有積極成效。
不同養殖類型調查結果顯示,核心育種場感染程度(2.66%)低于擴繁場(11.82%)和育肥場(11.73%),差異顯著(P<0.05)。核心育種場配備了更嚴格的生物安全制度,飼養管理水平更高,且是自繁自養體系,杜絕了外來攜帶病毒豬只的進入;PRRSV 也是核心育種場選育體系的凈化病原之一,抗原陽性豬只會被優先淘汰。
不同生長階段豬群調查結果顯示,各生長階段豬群的感染程度差異顯著(P<0.05),其中保育豬陽性檢出率最高,其余依次是生長育肥豬、哺乳仔豬和母豬,而種公豬普檢未發現PRRSV 感染。保育豬陽性檢出率最高的原因可能是,保育階段處于母源抗體衰減和疫苗免疫的臨界點。何海建等[16]研究表明,隨著日齡增加,仔豬體內的母源抗體水平逐漸降低,至42 日齡時,仔豬體內基本檢測不出PRRSV 抗體。所調查的規模豬場大多使用PRRSV 滅活苗,而滅活苗的免疫抗體產生期較長,同時由于個體差異的存在,部分保育豬只未能達到免疫全覆蓋,因此免疫程序需根據抗體消長規律制定[17]。再加上保育豬的內臟器官尚未發育成熟,自身免疫力低,在高飼養密度的環境下,易通過接觸和空氣傳播造成大規模感染[18]。保育豬是PRRSV 感染的重點豬群,須作為重點防控對象實時監測PRRSV 抗體消長規律,建立合理的免疫程序,同時提高飼養管理水平,降低保育豬的感染程度,這對各階段豬只PRRS 防控意義重大。
不同季節調查結果顯示,PRRSV 一年四季均可流行,但與夏季(6.94%)、秋季(6.10%)、冬季(7.44%)相比,春季(18.16%)是PRRSV感染的高峰期。出現該季節性差異特征可能是由于湖北省屬于亞熱帶季風氣候,夏秋季節干燥炎熱的環境易使PRRSV 失去感染性,而春冬季寒冷潮濕的環境則有利于其存活[15]。在當前非洲豬瘟防控的壓力下,國內許多豬場完善了生物安全制度,通過帶豬消毒這一措施進行防控,在炎熱的夏秋季節,帶豬消毒會減少熱應激對豬只的影響,而在寒冷的春冬季節,為了保持豬舍內的溫度,極大減少了室內外空氣的流通,此時帶豬消毒加劇了豬舍環境的濕度,導致PRRSV更易通過環境傳播。相較于冬季,春季的氣溫更加多變,冷熱應激對豬只的影響更大,因此春季PRRSV 的感染最為嚴重。因此,春季需要加強PRRS 防控,在確保豬舍內溫度的同時要保持豬舍內干燥。
陽性樣品中不同毒株感染結果顯示,類NADC30 毒株檢出占比(49.54%)高于高致病性變異毒株(29.10%)和經典毒株(21.36%),差異顯著(P<0.05),說明類NADC30 毒株是所調查規模豬場的優勢流行毒株。目前國內市場上的PRRSV 商品化疫苗主要是基于經典毒株和高致病性變異毒株研制而成的,對類NADC30 毒株的交叉保護力度不夠[19-20]。并且類NADC30 毒株之間差異較大,變異速度快,易與其他毒株發生重組變異[4],使PRRSV 基因型更加復雜,這給養殖場PRRS 防控帶來更大挑戰。在PRRSV 基因型復雜多變的情況下,需要實時監測毒株變異情況,分析出毒株的遺傳進化規律,提前制定出相應的防控方案[13]。
綜上所述,要做好PRRS 防控工作,需要采取綜合防控策略:一要制定嚴格的生物安全條例,建立完善的生物安全體系;二要堅持自繁自養,確需引種,則需制定完善的引種制度,避免外來攜帶病毒豬只的進入;三要實時監測豬只PRRSV 抗體水平及毒株的遺傳進化規律,重點做好保育階段豬只和春季時節的PRRS 防控。
猜你喜歡
環球時報(2022-04-25)2022-04-25 17:20:21
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
今日農業(2021年15期)2021-10-14 08:20:18
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
人大建設(2020年3期)2020-07-27 02:48:40
今日農業(2019年14期)2019-09-18 01:21:44
中國衛生(2016年8期)2016-11-12 13:27:08
