CD24 通過調節肝內Ly6Clo 巨噬細胞亞群中PDGF-B 的分泌抑制肝纖維化進程
2022-06-08 02:17:50靳悅汪河李明威張學軍
天津醫科大學學報 2022年3期
關鍵詞:小鼠
靳悅,汪河,李明威,張學軍
(天津醫科大學基礎醫學院免疫學系,天津 300070)
肝纖維化是肝臟損傷后細胞外基質過度沉積與異常分布的修復反應,是各種慢性肝病向肝硬化發展過程中的關鍵階段[1]。研究證明肝內巨噬細胞是肝纖維化進程及轉歸的重要免疫細胞[2-3],單核來源巨噬細胞被招募到受損的肝臟后,可通過分泌轉化生長因子-β(transforming growth factor-β,TGF-β)等促纖維化因子介導的肝星狀細胞活化、分化和增殖,進而促進慢性纖維化進程[4]。
血小板衍生生長因子(platelet derived growth factor,PDGF)是一種促進肝星狀細胞分裂和增殖的生長因子,分為PDGF-A、B、C 和D 4 種不同亞型[5-6],其中PDGF-B 是促進纖維化和HSC 增殖的關鍵細胞因子,它的主要受體PDGFR-β 在活化的HSC 上顯著上調[7-8]。在近幾年的纖維化機制研究中,PDGF信號通路已被認為是與HSC 活化和纖維化進展相關的重要通路[9-10]。
CD24 是一種糖基磷脂酰肌醇錨定分子[11],由多種免疫細胞表達[12]。CD24 也是一種重要的炎癥負反饋分子,在肝損傷和抗腫瘤先天免疫反應中發揮重要作用[13-14]。本研究利用阻塞性膽汁淤積引起的肝纖維化動物模型探討了CD24 分子是否通過調節肝內巨噬細胞分泌PDGF-B 進而延緩纖維化進程。
1 材料和方法
1.1 材料
1.1.1 實驗對象及分組 C57BL/6J 小鼠(購自北京斯貝福生物技術有限公司) 與CD24 基因敲除(CD24-/-)小鼠在天津醫科大學實驗動物中心共同飼養。各基因型均采用6~8 周齡,體重18~20 g 的雌性小鼠。各基因型均分為造模組與假手術組,每組5 只,造模組進行膽總管結扎(BDL)誘導小鼠纖維化,假手術組和造模組處理相同但不結扎膽總管,作為對照組,術后飼養于相同的環境,術后7 d 處死小鼠。
1.1.2 主要試劑和儀器 石蠟切片機購自leica,QPCR 儀LightCycler96 購自Roche 公司,Trizol 購自美國Invitrogen 公司,反轉錄酶試劑盒購自美國Promega公司,SYBR Green 購自Genstar,Percoll 購自英國GE Healthcare 公司,Ⅳ型膠原酶購自Sigma,流式抗體購自美國eBioscience公司,Western印跡抗體購自SantaCruz。
1.2 實驗方法
1.2.1 BDL 小鼠模型的構建 手術組按照4 μL/g腹腔注射10%水合氯醛溶液進行麻醉,之后刮去腹壁周圍絨毛暴露皮膚,消毒后使用手術刀沿腹中線切開腹壁,暴露腹腔,找到膽總管用消毒后的手術線結扎,最后縫合腹壁。術后小鼠飼養于消毒后的籠子里,置于溫暖的環境中。
1.2.2 肝組織的病理染色 冷PBS 灌注肝臟,取下的肝臟在10%甲醛溶液中固定至少24 h,依次進行不同濃度梯度的酒精脫水,浸蠟6~8 h 后包埋。用石蠟切片機進行5 μm 厚度的切片,60℃烤片1 h 后進行HE和天狼星紅染色,其中,蘇木素染色3 min,分化液30 s,伊紅1 min,天狼星紅染色1 h,染色完成后中性樹膠封片,在Nikon 90i 顯微鏡下拍照,每張片子選取5 個視野,用image J 軟件統計陽性區域面積進行比較。
1.2.3 免疫印跡 將液氮速凍的肝組織加入適量裂解液,置于冰上,使用手持組織破碎儀將組織打成勻漿,再經超聲破碎儀對組織勻漿進行超聲破碎,超聲15 s,停10 s,循環3 次。將超聲后的樣品離心后轉移上清至新的EP 管,加入Loading buffer 后5 min,100℃變性。加載等量的蛋白通過10%SDS-PAGE 分離,然后轉移到PVDF 膜上,300 mA 轉膜2 h,用含5%BSA 的TBST 緩沖液封閉1 h,然后用相應的抗體孵育過夜。用對應的二抗室溫孵育1 h,洗膜3~5 遍。α-SMA(1∶500),β-actin(1∶500),辣根過氧化物酶標記的二抗(1∶5 000),通過化學發光液檢測抗體條帶。
1.2.4 定量-PCR 將液氮速凍后的肝組織加入1 mL Trizol 裂解,培養皿中的細胞加入600 μL Trizol裂解,異丙醇沉淀法提取RNA,用分光光度計測定RNA 濃度后,每個樣品取1~2 μg,用反轉錄試劑盒(Promega)進行反轉錄。將得到的cDNA 進行10 倍稀釋,按照cDNA 4.5 μL,引物0.5 μL,SYBR Green 5 μL 的體系使用LightCycler96 運行程序。具體的程序為:預變性:95℃,300 s;設置50 個循環:變性:95℃,15 s;退火:60℃,30 s;延伸:72℃,30 s;溶解:95℃,10 s,65℃,60 s,97℃,1 s;冷卻:37℃,30 s。以Actin 基因為內參,用2-ΔΔCt計算結果,所有用到的引物序列見表1。反轉錄后的樣品加入SYBR Green 嵌合熒光染料并使用LightCycler96 進行QPCR 檢驗。

表1 定量-PCR 所用引物序列Tab 1 Primer sequence used for Q-PCR
1.2.5 制備肝內巨噬細胞與星狀細胞 肝組織依次灌注EGTA 溶液、鏈酶蛋白酶溶液和Ⅳ型膠原酶溶液,將肝臟在消化液中剪碎,37℃、160 r/min 消化15 min,將消化好的肝臟通過70 μm 的篩網過濾,4℃,50×g,3 min,多次離心去除肝細胞。4℃,500×g,8 min 離心2 遍棄上清,用1×PBS 與預先配置的Percoll 原液混合成50%和25%的Percoll,用25%Percoll重懸細胞后緩慢滴加到50%Percoll 上,2 000 r/min,制動0 離心30 min,取Percoll 中間層細胞即為肝內巨噬細胞。將濾過的單細胞懸液4℃,580×g,離心10 min,棄上清到10 mL,加120 μLDNaseⅠ重懸,補GBSS/B 至50 mL,4℃,580×g,離心10 min,棄上清至10 mL加120 μL DNaseⅠ,將細胞懸液平均分到兩個15 mL管中,加GBSS/B 至8 mL,再加入4 mL Nycodenz,混勻后在細胞懸液上加1.5 mL GBSS/B,制動0,1 380×g,4℃,離心17 min,取白膜層即為肝星狀細胞。
1.2.6 制備肝非實質細胞(NPCs)懸液 步驟同提取肝內巨噬細胞,棄上清留底部沉淀后,將10×PBS與預先配置的Percoll 原液混合成30%Percoll 并重懸細胞,450×g,20 min 離心留下最底層細胞,冷ACK進行紅細胞裂解,裂解完成后洗兩遍重懸計數。
1.2.7 流式細胞術檢測 制備肝非實質細胞懸液后,每管取1×106個細胞,用CD16/32 抗體4℃封閉15 min,之后再進行細胞表面染色30 min。所用抗體如下:Apc-cy7 Anti-mouse CD45、FITC Anti-mouse Ly6G、Percp-cy5.5 Anti-mouse CD11b、APC Antimouse F4/80、PE-cy7 Anti-mouse Ly6C、PE Antimouse CD24。使用BD FACS CantoⅡ流式分析儀進行檢測。Flowjo 10.6.2 軟件對實驗結果進行分析。
1.2.8 巨噬細胞亞群分選 制備肝非實質細胞懸液后,將懸液轉移到無菌流式管中,4℃,500×g,離心5 min,冰上封閉15 min,再用標記抗體Apc-cy7 Anti-mouse CD45、FITC Anti-mouse Ly6G、Percp-cy5.5 Anti-mouse CD11b、APC Anti-mouse F4/80、PE-cy7 Anti-mouse Ly6C 染色30 min,用流式PBS 洗兩遍后用FACS AriaⅡ流式分選儀分選巨噬細胞亞群,收集到無菌管中。
1.2.9 共培養 將提取的原代星狀細胞種于6 孔板,于37℃,5%CO2的細胞培養箱中培養。將提取的巨噬細胞亞群加入提前架好的Transwell 培養小室的6 孔板中,培養條件相同。在原代星狀細胞培養兩天貼盤穩定后,將Transwell 小室放置于培養原代星狀細胞的6 孔板上,共培養3 d,提取RNA 用于Q-PCR 檢測。
1.3 統計學處理 使用Graphpad Prism8 進行數據匯總、分析、制圖,數據服從正態分布,以±s 表示,多組間比較采用單因素方差分析,P<0.05 為差異有統計學意義。
2 結果
2.1 BDL 誘導的CD24-/-小鼠肝纖維化程度加重 首先建立了BDL 誘導的小鼠纖維化模型,HE 結果顯示,在造模組中,與WT 小鼠相比,CD24-/-小鼠肝組織炎性細胞浸潤增多,匯管區增生更加明顯,而假手術組中兩種基因型小鼠的肝臟則沒有明顯病理改變(圖1A)。天狼星紅染色結果顯示,造模組CD24-/-小鼠的肝纖維化程度更嚴重(P<0.01),而假手術組無明顯差別(P>0.05,圖1B)。Q-PCR 結果顯示,造模組CD24-/-小鼠的肝纖維化指標α-SMA(P<0.05)、Col1α1(P<0.01)和Timp-1(P<0.000 1)與WT 小鼠相比有顯著升高,假手術組無明顯差別(P>0.05,圖1C)。Western 印跡結果顯示,肝纖維化標志物α-SMA 在CD24-/-小鼠的肝組織中表達明顯高于WT 小鼠(圖1D),上述結果提示,CD24-/-小鼠的肝損傷和肝纖維化程度明顯加重。
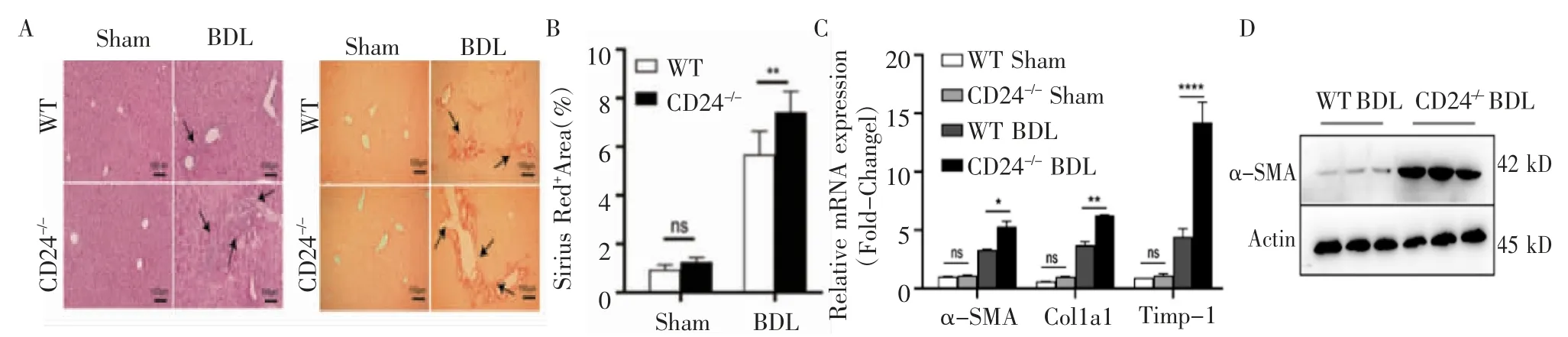
圖1 CD24 基因敲除后BDL 誘導的小鼠肝纖維化加重Fig 1 BDL-induced liver fibrosis in mice aggravated after CD24 geng knockout
2.2 CD24-/-小鼠肝纖維化加重 與WT 小鼠相比,CD24-/-小鼠造模后肝臟中定居的巨噬細胞(CD11bhiF480hi) 比例與數量都沒有明顯差別(P>0.05),募集巨噬細胞群(CD11bhiF480int)比例明顯增加(P<0.001),數量也有增加(P<0.05,圖2A)。進一步分析Ly6Chi和Ly6Clo兩種肝內募集型巨噬細胞亞群,結果發現CD24-/-小鼠造模后的Ly6Clo巨噬細胞亞群比例高于WT 小鼠(P<0.05),數量也高于WT 小鼠(P<0.01),而Ly6Chi巨噬細胞亞群比例和數量均無明顯差異(P>0.05,圖2A)。分析CD24分子在肝內不同的巨噬細胞亞群中的表達發現,CD24 分子在Ly6Clo巨噬細胞亞群中高表達且造模前后差異明顯(P<0.05,圖2B)。分選肝內巨噬細胞亞群與提取的原代星狀細胞共培養后檢測星狀細胞的纖維化指標,結果發現,造模后的CD24-/-小鼠肝內Ly6Clo群巨噬細胞亞群對星狀細胞的活化作用較強,纖維化指標表達明顯升高(圖2C)。
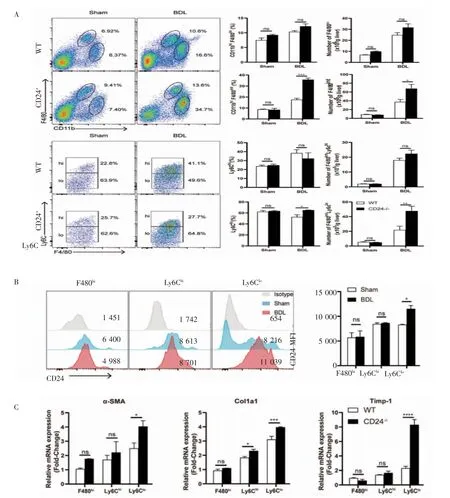
圖2 CD24-/-小鼠肝纖維化加重可能與Ly6Clo 巨噬細胞亞群相關Fig 2 The aggravation of liver fibrosis in CD24-/-mice was related to Ly6Clo macrophages subset
2.3 CD24-/-小鼠纖維化與Ly6Clo巨噬細胞分泌PDGF-B 因子有關 在造模組中,與WT 小鼠相比,CD24-/-小鼠肝組織中PDGF-A 和PDGF-B mRNA表達較高且有顯著差異(P<0.05,P<0.01),而PDGFC 和PDGF-D 表達低且無差異(P>0.05,圖3A)。提取肝內巨噬細胞檢測發現,Q-PCR 顯示PDGF-A 與PDGF-Bm RNA 在CD24-/-小鼠造模后的肝內巨噬細胞上表達均有顯著升高,其中以PDGF-B mRNA表達最為明顯(P<0.01,P<0.000 1,圖3B)。進一步分析肝內不同巨噬細胞亞群PDGF-A 和PDGF-B mRNA 表 達,結 果 發 現,PDGF-B 在CD24-/-小 鼠 的Ly6Clo巨噬細胞亞群上高表達(P<0.000 1,圖3C)。用巨噬細胞上清與肝星狀細胞共培養觀察星狀細胞上PDGF-B 受體的表達,結果發現,與CD24-/-小鼠肝內Ly6Clo巨噬細胞共培養后,肝星狀細胞PDGFR-β 受體mRNA 表達量顯著上調(P<0.000 1,圖3D)。
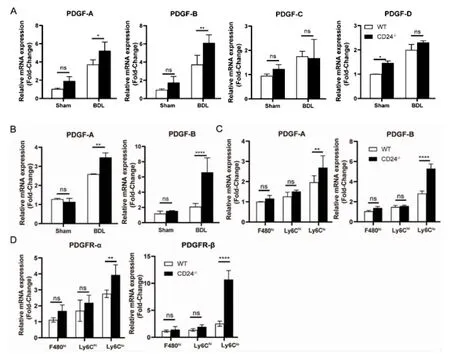
圖3 CD24 小鼠纖維化與Ly6Clo 巨噬細胞分泌PDGF-B 因子密切相關Fig 3 CD24 mouse fibrosis closely related to the secretion of PDGF-B factor by Ly6Clo macrophages
3 討論
在BDL 誘導的纖維化中,巨噬細胞與纖維化發展密切相關[15],在不同的巨噬細胞亞群中,單核衍生的Ly6C 巨噬細胞亞群可在纖維化的不同階段發揮重要作用[16-17]。PDGF-B 被認為是巨噬細胞分泌的促纖維化因子,可促進星狀細胞的增殖分化以控制肝纖維化的發展[18]。
在WT 鼠和CD24-/-小鼠經過膽總管結扎術造成纖維化后,初步發現CD24-/-小鼠的纖維化情況較WT 小鼠嚴重。在研究機制的過程中發現CD24-/-造模小鼠中募集群巨噬細胞顯著增多,CD24 分子在Ly6Clo群巨噬細胞上高表達且在造模前后有明顯差異,提示CD24 分子可能通過Ly6Clo群巨噬細胞來調節肝纖維化。
為了探究巨噬細胞分泌的相關纖維化因子,首先應用Q-PCR 檢測篩選了與纖維化密切相關的PDGFs 家族成員的mRNA 表達變化,結果發現,PDGF-A 和PDGF-B 在CD24-/-小鼠肝纖維化組織中表達顯著升高且與WT 小鼠有明顯差異,PDGFC 和PDGF-D 表達低且無明顯差異,該結果初步排除PDGF-C 和PDGF-D 的作用。進一步檢測肝內巨噬細胞,結果發現,PDGF-A 和PDGF-B 在造模后的CD24-/-小鼠肝內巨噬細胞上表達升高,尤其是PDGF-B 表達顯著升高,因此PDGF-A 和PDGF-B與CD24 敲除后肝纖維化加重有關。深入分析巨噬細胞亞群發現,PDGF-A 和PDGF-B 在CD24-/-小鼠Ly6Clo巨噬細胞亞群上高表達且與WT 小鼠有顯著差異,并且PDGF-B 的表達差異更大,因此,CD24主要通過Ly6Clo巨噬細胞亞群分泌PDGF-B 來調節纖維化的發展。巨噬細胞分泌的PDGFs 等促纖維化因子是通過調節肝星狀細胞表達的相應受體來實現的[19],為了驗證上述結果,首先將不同巨噬細胞亞群與星狀細胞共培養,檢測星狀細胞上的相應受體的變化,結果發現,與CD24-/-小鼠來源的Ly6Clo巨噬細胞亞群共培養后的星狀細胞上PDGFR-β 受體表達顯著升高,而PDGFR-β 是主要與PDGF-B相結合的配體[20],因此,共培養實驗進一步驗證了PDGF-B 是CD24 通過Ly6Clo巨噬細胞亞群調節纖維化的重要因子。綜上所述,CD24 分子可通過影響Ly6Clo巨噬細胞PDGF-B 的分泌來調節纖維化。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
