海南鸚哥嶺附生蘭物種組成、分布及與生態因子的關系
2022-06-13 01:46:56張培春楊小波曾潤娟戚春林李東海田璐嘉李晨笛梁彩群
熱帶生物學報 2022年2期
張培春,楊小波,夏 丹,王 群,王 豪,曾潤娟,戚春林,李東海,陳 琳,田璐嘉,李晨笛,李 龍,梁彩群
(海南大學 生態與環境學院,海口 570228)
蘭科(Orchidaceae)植物類群龐大,多具有觀賞、藥用、食用等經濟價值。由于其對生境的獨特需求,多以小種群形式分布,較其他植物更容易受到生境退化或喪失的威脅[1]。目前,世界所有蘭科植物已被列入《瀕危野生動植物物種國際貿易公約》加以保護,成為植物保護中的1 個“旗艦”類群[2]。在國家林業和草原局、農業農村部于2021 年9 月最新公布的《國家重點保護野生植物名錄》中,野生蘭科植物共涉及到23 屬349 種,野生蘭科植物的保護現已受到空前重視。
附生型蘭科植物(Epiphytic Orchids)(以下簡稱附生蘭)為沒有地下生長階段,通過氣生根附著于巖石或樹干上的蘭科植物。因為生長的依附性,附生蘭科植物對附生環境和基質要求更為嚴格,對環境變化更為敏感,更容易形成斑塊狀分布[3]。同時,附生蘭生長緩慢,通常1 個生長軸1 年僅發1 個新芽,從萌發到成熟需要超過1 年的時間,當生境和種群遭到破壞后,恢復極為緩慢[4]。附生蘭約占全部附生植物種數的10%~25%[5?6],而附生植物是熱帶、亞熱帶濕性山地森林和少部分溫帶雨林的重要組成部分,對維持森林生態系統生物多樣性、水分平衡、養分循環、為動物提供食物和庇護場所等方面的貢獻不可忽略[7]。因此,研究附生蘭的分布特征可對附生蘭科植物的生境保護和
植株繁育提供理論依據,進而對生態系統的生物多樣性和功能的維持意義重大。近年來,對于附生蘭分布的研究多集中于地區附生蘭物種多樣性與組成[8?13],或側重個別環境因子如海拔[14?16]、溫度[9,16?17]、降水[9,16?17]、植被類型[17?20]等對附生蘭科植物分布的影響,或少數附生蘭如華石斛(Dendrobium sinense)等的適宜生境模擬,而鮮有綜合研究附生蘭分布的多因素作用的報道[17,21],這對于附生蘭生境保護與多樣性維持是不利的。與地生植物不同,附生蘭遠離土壤基質,更多地受到附生基質與氣候因子的影響,因而附生蘭的影響因子分析中應將諸多的氣候因子與附生基質(
樹皮)特征納入其中。相關的少許研究僅局限于年平均氣溫與年降雨量的變化上,時間尺度上過于寬泛,不利于解釋附生蘭生境的氣候差異[13,17],附生基質對附生蘭分布影響的研究報道同樣少見[20]。附生蘭在世界范圍內大多面臨著各種人為因素的生存威脅。要制定地方乃至全國的附生蘭科植物保育及資源合理開發利用策略,通過系統、綜合的附生植物與環境因子的關聯分析研究來建立理論指導是亟待解決的問題。本研究擬結合附生蘭分布狀況與分布點上對附生蘭生存起重要作用的各類生態因子如海拔、地形、植被類型、氣候等,綜合分析鸚哥嶺附生蘭科植物的生境適應規律,旨在為海南熱帶雨林國家公園內附生蘭的保育、適宜生境選擇、氣候變化背景下的種群數量預測及未來的資源開發利用提供理論依據。
1 研究方法
1.1 研究區域概況鸚哥嶺自然保護區,現為海南熱帶雨林國家公園管理局鸚哥嶺分局所轄的自然保護地,位于海南島中南部,地理坐標為109°11′ 27″~109° 34′ 06″ E,18° 49′ 30″~19° 08′ 41″ N,總面積50 464 hm2。地形以山地為主,保護區內最高峰為鸚哥嶺,海拔1 812 m,是海南第2 高峰。保護區屬熱帶海洋性季風氣候,年均氣溫20~24 ℃,在相同高度上,年均氣溫由東北向西南遞增。年平均降雨量1 400~2 200 mm,但季節分配不均,東南坡降雨明顯高于西北坡。依據楊小波等[22]的《海南植被志》,研究區域林區的植被可劃分為熱帶針葉林、高山云霧林、山地雨林、低地雨林、低地雨林次生林、季雨林、旱地農業生產植被等植被型或植被亞型。
盧剛等[23]通過多年的考察,出版了《海南鸚哥嶺蘭科植物圖鑒》,共記錄到野生蘭科植物88 屬204 種。其中,附生蘭科植物104 種,占總數的51.00%。該調查成果為本次研究提供了寶貴資料。
1.2 野外調查方法
1.2.1 樣線設置調查前查閱已有資料記錄,結合野外踩點,根據鸚哥嶺自然保護區的地形、地貌、附生蘭科植物多樣性概況,在各類地形、海拔段、植被區、小生境、附生蘭豐富的區域設置樣線。
1.2.2 調查內容以每條進入林區的路徑為樣線,以樣線兩側5 m 內出現的有蘭科植物附生的喬木或藤本作為調查樣木,記錄附生蘭物種及其宿主信息。參考《海南植物圖志》[24]、《鸚哥嶺蘭科植物圖鑒》[23]、《中國野生蘭科植物原色圖鑒》[25]確定宿主樹、附生蘭科植物的物種,中文名稱、拉丁學名均參照《中國植物志》[26]2019 網頁版中的植物名稱。樣木所在地理坐標及海拔用GPS 記錄,坡向和坡度分別用羅盤、坡度儀進行實測,植被郁閉度采用抬頭望法估測。
于2020 年8 月至2021 年8 月期間分不同季節進行調查,樣木分布情況見圖1。共設置樣木477 個,調查到附生蘭40 屬78 種。
1.3 其他數據來源氣溫和雨量數據來自世界氣候數據網站(http: //www.worldclim.org/)。
1.4 數據處理
1.4.1 野 外 調 查 數 據 處 理應 用 公 式(式中,S 為坡向原始數據,S1為轉化后的坡向數據)將坡向數據轉換為0~1 之間的數據,轉換后的數據越大表示生境條件越干熱,越小則說明生境條件越濕冷;坡向在0°~22.5°和337.5°~360°為北坡,22.5°~67.5°為東北坡,67.5°~112.5°為東坡,112.5°~157.5°為東南坡,157.5°~202.5°為南坡,202.5°~247.5°為西南坡,47.5°~292.5°為西坡,292.5°~337.5°為西北坡;附生蘭物種豐富度由附生蘭物種數表示,附生蘭多度由附生蘭個體數量表示。S1=(1?cos((π/180)(S ?30)))/2
1.4.2 排序數據處理排序及制圖由Canoco for Windows 5.0 完成。排序的物種數據由78 種附生蘭物種在447 個樣木內出現或者不出現(0,1)組成,其矩陣組成(樣方×物種)為447 行×78 列;環境數據矩陣由實測的6 種環境因子和來自世界氣候網站的19 種氣候因子組成,為447 行×25 列。
圖 1 野外調查樣木分布圖
1.4.3 預分析為避免因氣候因子間存在多元共線性而導致的排序結果的不可靠,首先采用前向選擇(forward-selection)通過自變量效應由大到小手動選取氣候因子。被選取的氣候因子有年平均溫度、每月最高氣溫與最低氣溫差值的平均值(以下簡稱平均月溫差)、最干月降雨量。預先進行除趨勢對應分析(detrended correspondence analysis,DCA),結果顯示,最長梯度大于4,說明物種組成數據存在較大差異,故選用典范對應分析(canonical correspondence analysis, CCA)研究環境因子對物種組成的影響。
1.4.4 CCA 重分析以選取的3 種氣候因子和6 種生境因子進行重分析。為明確各個因子對蘭科植物物種組成影響的相對重要性,參照文獻[28]的方法計算每一個環境因子對蘭科植物物種組成的總效應與凈效應,僅以1 個環境因子作為解釋變量進行CCA 排序所得的物種組成變異的解釋量為此環境因子的總效應,而以1 個環境因子作為解釋變量,其他環境因子作為協變量進行CCA 排序所得的物種組成變異的解釋量為此環境因子的凈效應,所有排序的顯著性均由Monte Carlo 隨機置換(999 次)進行檢驗。其余作圖由Origin 2018 完成。
2 結果與分析
2.1 環境因子與CCA 排序軸的相關性用于CCA 排序的9 個環境因子共解釋了鸚哥嶺附生蘭組成總變異的7.8%,基于所有典范特征值之和的Monte Carlo 隨機置換檢驗為極顯著(P< 0.01)(表1),說明物種與所選環境因子之間有著強烈的相關性。CCA 前兩軸有比第3 和第4 軸更高的特征值,說明前兩軸在對物種組成變異的解釋中起主導作用,第1 軸解釋了物種組成變異的1.82%,第2 軸解釋了物種組成變異的1.5%(表1)。CCA 第1 軸和第2 軸代表了不同的環境梯度。CCA 1 軸中,最干月降水量、海拔、山地雨林及熱帶針葉林與之呈顯著正相關,年平均氣溫、平均月溫差、樹皮粗糙、低地雨林次生林、旱地農業生產植被、低地雨林及坡向與之呈顯著負相關;主要為海拔梯度,溫度梯度及降雨量梯度(高的相關系數)。CCA 2 軸中年均氣溫、樹皮有覆蓋物、低地雨林次生林、熱帶針葉林與之呈顯著正相關關系,最干月降水量、海拔、郁閉度山地雨林等與之呈顯著負相關關系;表明該軸為1 個綜合軸,反映了氣溫、植被類型等的變化。
基于Monte Carlo 隨機置換的檢驗結果表明(表2),除坡度因子外,其余生境因子均顯著地解釋了附生蘭組成的變異。凈效應由大到小依次為植被類型、樹皮類型、年平均氣溫、每月最高氣溫與最低氣溫的差值平均值、最干月降雨量、海拔、郁閉度、坡向、坡度,僅坡度的凈效應未達到顯著。
2.2 環境因子對附生蘭科植物物種組成的影響
在進行排序分析的78 種蘭科植物中,分布較廣的種有金塔隔距蘭(Cleisostoma filiforme)、釵子股(Luisia morsei)、海南石斛(Dendrobium hainanense)、純色萬代蘭(Vanda subconcolor)、大序隔距蘭(Cleisostoma paniculatum)等,發生樣木數分別為55、50、41、40、32,在總樣木數(447)中的占比分別為12.30%、11.86%、9.17%、8.95%、7.16%。而分布樣木數僅為1 的稀有種類也達到了14 種,包括胼胝蘭(Biermannia calcarata)、蛇舌蘭(Diploprora championii)、短耳鳶尾蘭(Oberonia falconeri)、厚唇 蘭(Epigeneium clemensiae) 、 鐮 葉 盆 距 蘭(Gastrochilus acinacifolius)等。
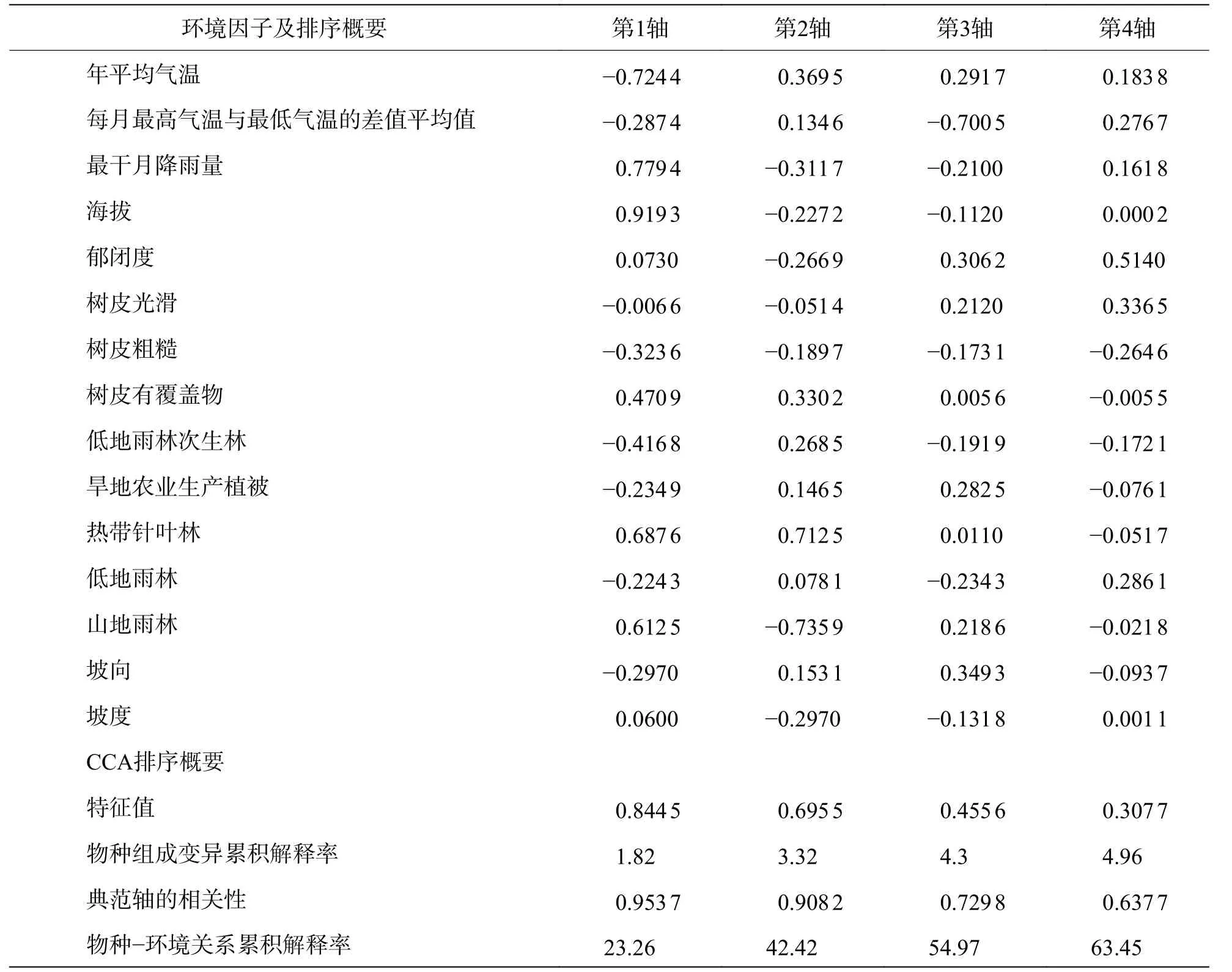
表1 環境因子與典范對應分析(CCA)前4 排序軸的相關系數及排序概要
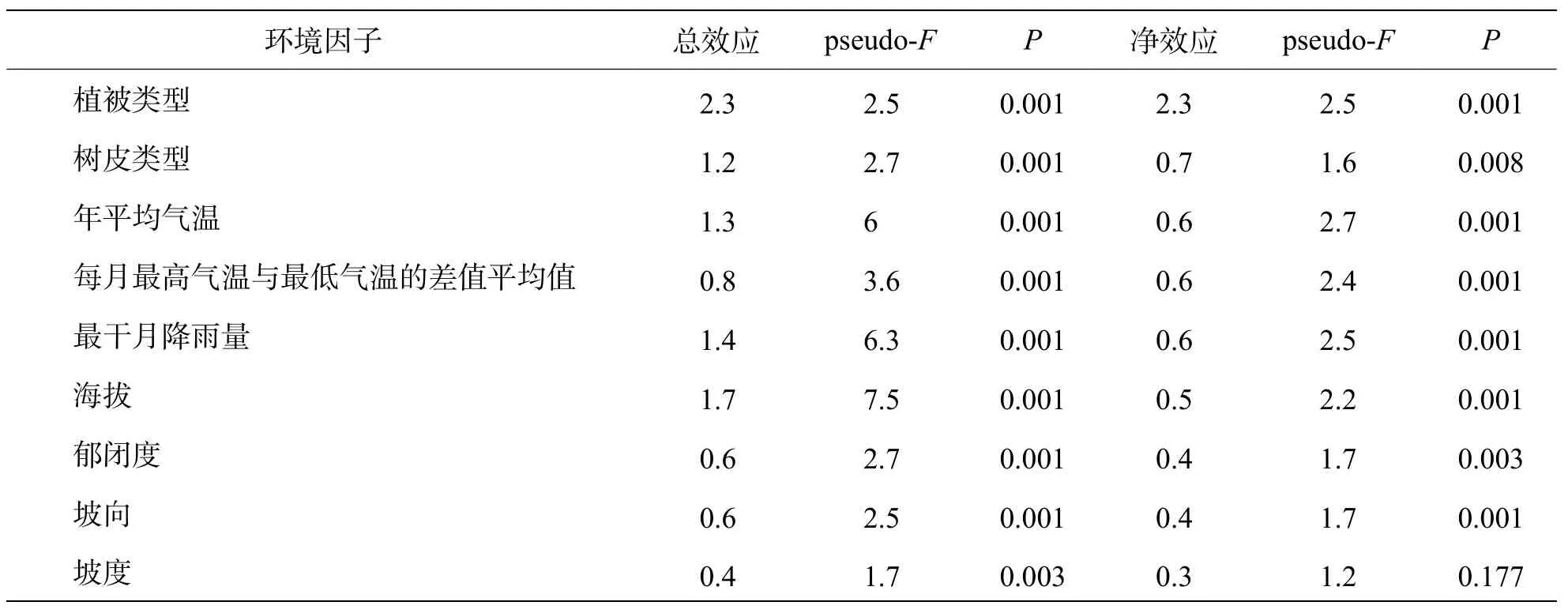
表2 環境因子對物種組成的總效應與凈效應
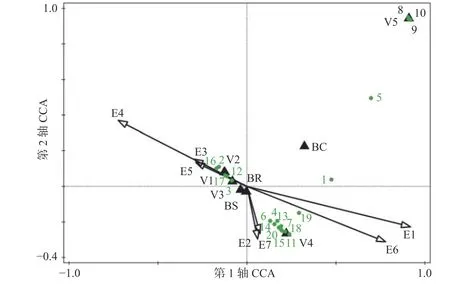
CCA 雙序圖(圖2)中展示的受生境因子影響最顯著的20 種蘭科植物。位于排序圖右上角的物種,如白唇槽舌蘭(Holcoglossum subulifolium)、樂東石豆蘭(Bulbophyllum ledungense)、伏生石豆蘭(Bulbophyllum reptans)、黑毛石斛(Dendrobium williamsonii)等附生蘭,對應的生境條件為中高海拔、較多的降雨量、較低的年均氣溫及較小的平均月溫差,其多生于濕冷、樹皮有厚覆蓋物的的熱帶針葉林中的宿主上;位于排序圖中下部的物種,如石斛(Dendrobium nobile)、狹葉金石斛(Flickingeria angustifolia)、石仙桃(Pholidota chinensis)、粗莖蘋蘭(Pinalia amica)、指葉擬毛蘭(Mycaranthes pannea)、匙唇蘭(Schoenorchis gemmata)、牛角蘭(Ceratostylis hainanensis)等附生蘭,對應的生境為中低海拔地段、坡度大、郁閉度較高、降雨較為充沛的山地雨林;位于雙序圖左部的附生蘭,如多花脆蘭(Acampe rigida)、海南石斛(Dendrobium hainanense)、釵子股(Luisia morsei)、金塔隔距蘭(Cleisostoma filiforme)、大尖囊蝴蝶蘭(Phalaenopsis deliciosa)等,對應的生境條件為低海拔、較高的年平均溫度、較大的平均月溫差,其分布區多為旱地農業生產植被或低地雨林次生林,往往生長于陽坡。其他未展示的大部分種類出現在排序圖中心點附近,它們的分布受到生境因子的綜合影響相對較小。
2.3 環境因子對附生蘭物種豐富度和多度的影響在垂直水平上,把鸚哥嶺林區劃分為8 個海拔段,各海拔段附生蘭物種豐富度及物種多度情況見圖3-A。隨著海拔的升高,附生蘭物種數呈現出先上升后下降的趨勢。700~900 m 海拔段出現峰值,生長有47 種附生蘭,占調查到的總物種數的60.26%;1 300~1 500 m 海拔段附生蘭物種數最少,僅芳香石豆蘭、流蘇貝母蘭、厚唇蘭、黑毛石斛4 種,占總物種數的5.13%。附生蘭個體數隨著海拔的升高而遞增。300 m 以下海拔段平均每木有附生蘭33.51 株,而在海拔1 500 m 以上平均每木附生蘭達686 株。
研究區域的坡度可劃分為6 個等級(圖3-B),5°以下為平坡,5°~15°為緩坡,15°~25°為斜坡,25°~35°為陡坡,35°~45°為急坡,45°以上為險坡(含最小值不含最大值,下同)。緩坡的附生蘭物種數最多,達63 種,占總物種數的80.77%;陡坡的附生蘭物種數最少,僅4 種,分別為蘚葉卷瓣蘭、石斛、白點蘭、金塔隔距蘭,占總物種數的5.13%。附生蘭個體數目的峰值同樣出現在緩坡地段,平均每木著生有附生蘭93.59 株,平坡地段平均每木附生蘭個體數最少,為31.52 株。
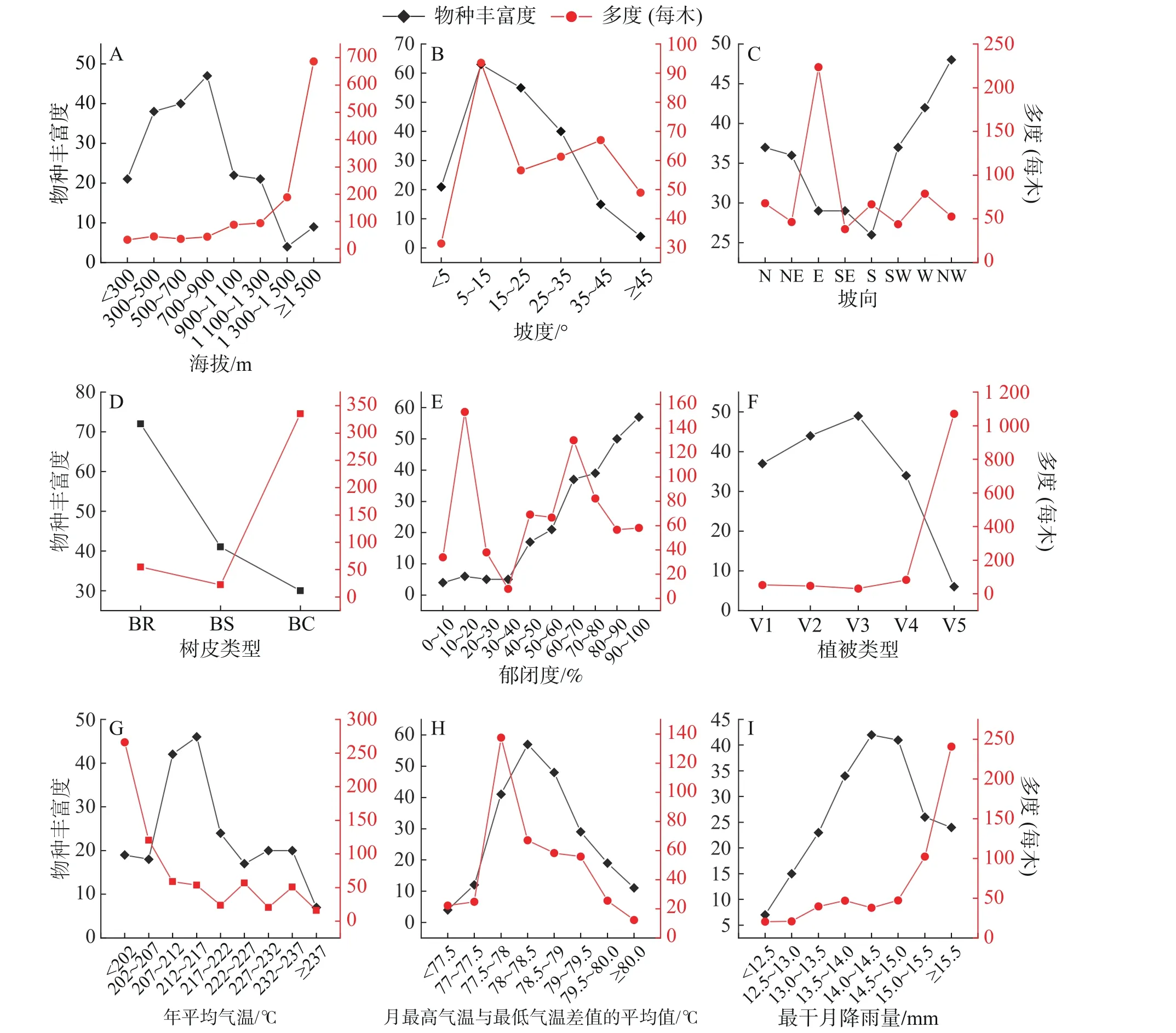
圖 3 附生蘭物種豐富度、多度與9 個生態因子的關系A.海拔;B.坡度;C.坡向;D.樹皮類型;E.郁閉度;F.植被類型;G.年平均氣溫;H.每月最高氣溫與最低氣溫差值的平均值;I.最干月降雨量;V1.旱地農業生產植被;V2.低地雨林次生林;V3.低地雨林;V4.山地雨林;V5.熱帶針葉林;BR.樹皮粗糙;BS.樹皮光滑;BC.樹皮有覆蓋物。
圖3-C 顯示,在研究區域的8 個坡向地段中,附生蘭物種分布數目較多的為西北坡、西坡、北坡,而南坡、東南坡、東坡物種分布數目相對較少,南坡分布的附生蘭物種數最少,為26 種,占總物種數的33.33%,多為相對耐旱物種,如釵子股、硬葉蘭,多花脆蘭,金塔隔距蘭、海南石斛等。東坡分布的附生蘭個體數量最大,平均每木達223.40株,其余坡向上附生蘭個體數量差別不明顯。
根據樹皮表面特征,將附生蘭樣木樹皮劃分為粗糙,光滑,有覆蓋物(如苔蘚植物)3 類(圖3-D)。樹皮粗糙的樣木上分布有附生蘭達72 種,占總物種數目的92.31%。而樹皮上有覆蓋物的樣木著生有個體數量較多的附生蘭,平均每木可達335 株,大量著生于有覆蓋物樹皮上的代表物種有樂東石豆蘭、伏生石豆蘭、黑毛石斛、指葉擬毛蘭、芳香石豆蘭等。
附生蘭在不同郁閉度區間的分布情況見圖3-E。附生蘭豐富度隨著郁閉度的增大而遞增,郁閉度在10%以下的林區僅分布有絨蘭、海南石斛、紅花隔距蘭、短耳鳶尾蘭等4 種附生蘭,占總物種數的5.13%,而在郁閉度90%以上的林區則分布有57 種附生蘭,占總物種數的73.77%。附生蘭個體數量在10%~20%、60%~70%兩個郁閉度區間出現峰值,暗示著不同的附生蘭對郁閉度有不同的偏好。
在本研究區域內,低地雨林中附生蘭物種豐富度最高,為49 種,占總物種數的62.83%,代表物種有純色萬代蘭、海南鉆喙蘭、白點蘭、紅花隔距蘭、海南石斛等,熱帶針葉林中分布的附生蘭物種最少,僅伏生石豆蘭、白唇槽舌蘭、流蘇貝母蘭、黑毛石斛、樂東石豆蘭、粗莖蘋蘭等6 種,占總物種數的7.69%。在熱帶針葉林種,附生蘭多度明顯較高,平均每木可達1 070 株,表明熱帶針葉林中的附生蘭具有較高的優勢度。
圖3-G 和3-H 分別展示了附生蘭物種豐富度、物種多度隨生境年平均氣溫、平均月溫差變化的情況。研究區域附生蘭物種豐富度隨年平均氣溫的升高呈現先升高后降低的趨勢,在21.2~21.7 ℃段出現峰值,物種豐富度達47 種,占總物種數的60.26%,年平均氣溫達到23.7 ℃以上時,附生蘭物種豐富度最低,僅大尖囊蝴蝶蘭、寄樹蘭、齒瓣鳶尾蘭、短序脆蘭、多花脆蘭、大葉寄樹蘭、硬葉蘭等7 種,占總物種數的8.97%;附生蘭多度隨著年平均氣溫的升高呈現出遞減的趨勢。隨著平均月溫差的增大,附生蘭物種豐富度和多度均呈現出先升高后降低的趨勢,溫差在7.75~7.90 之間的地段附生蘭物種豐富度及多度較高。
附生蘭物種豐富度及多度隨研究區域最干月降雨量的變化情況見圖3-I。附生蘭物種豐富度隨最干月降雨量增多而先升高后降低,在最干月降雨量為14~15 mm 地段附生蘭物種豐富度最高,達40 種以上。在12.5 mm 以下附生蘭分布很少,僅大尖囊蝴蝶蘭、寄樹蘭、齒瓣鳶尾蘭、短序脆蘭、多花脆蘭、大葉寄樹蘭、硬葉蘭等7 種,占總物種數的8.79%,而這些附生蘭均生長在溝谷旁林內。隨著最干月降雨量的增多,附生蘭多度呈現出遞增的趨勢。
3 討 論
本研究中的環境因子可劃分為地形因子如海拔、坡度和坡向等,氣候因子如年平均氣溫、平均月溫差和最干月降雨量等,生物因子如植被郁閉度、植被類型和樹皮類型等。
3.1 附生蘭物種組成與環境因子的關系
3.1.1 CCA 排序結果的解釋率CCA 分析確定了研究區域種類組成的主要梯度,并用解釋變量對其進行解釋。雖然環境梯度具有明顯的生態解釋,但CCA 排序軸解釋的變異很低(7.8%)。采用間接梯度分析DCA 排序進一步探索物種組成數據,結果顯示4 個軸對物種組成變異的解釋量為8.34,高于CCA 排序軸(4.96),表明該研究對部分生態因子如生境濕度,群落的發育歷史、樹皮的化學特性等的影響沒有考慮到,但是DCA 分析也表現出較低的變異量,表明潛在的缺失變量并不比包含的4 個變量更重要。同時,也說明研究區域的附生蘭分化較深入,物種組成十分復雜。物種組成解釋率較低的可能原因是:物種組成數據矩陣是由(0,1)組成,且物種數據矩陣中有較多的0 值;排序軸的多項式的失真[27]。但是,低的解釋率并不影響所選環境因子對附生蘭物種組成解釋的高顯著性。
3.1.2 附生蘭物種組成與地形因子、氣候因子及生物因子的關系海拔、坡度、坡向等地形因子一般通過影響濕度、光照、溫度、降雨等能量環境間接對陸生植物的分布產生影響,且地形因子其較其他環境因子易于測量,因此常用來解釋植物物種組成的變異[28?29]。本研究結果表明,海拔為影響鸚哥嶺附生蘭分布的重要環境因子,沿海拔梯度附生蘭組成出現明顯變異,基于總效應與凈效應的結果也清晰表明了其對物種組成影響的顯著性,相似的結論在其他地區的研究中也有報道[17,20?21]。有研究認為,坡向與坡度的變化,可引起生境光照,濕度的變化進而影響物種組成[30?31],在本研究中,坡向的總效應和凈效應均達顯著水平,而坡度盡管總效應達到了顯著水平,其凈效應卻不顯著。可能的原因是,附生蘭遠離土壤基質,常著生在高大喬木的冠層區,坡度對其的影響顯然要比地生植物小很多,余東莉等[12]和周康等[21]對其他地區蘭科植物的研究也有相似的結果。
光照、溫度、水分等生態因子直接影響著植物的生理過程,對植物的分布起著決定性的作用。對于附生蘭科植物而言,林區郁閉度決定了生境的光照條件,溫度則限制著附生蘭的分布,降水則更多的是對林區空氣濕度及附生基質(宿主樹皮)含水量的調節。有人通過研究得出年平均溫度、年降雨量等氣候因子對蘭科植物的物種組成具有顯著影響[17]。本研究中,郁閉度、年平均溫度、平均月溫差及最干月降雨量的總效應及凈效應均達到了高的顯著性,可見不同的附生蘭對氣候因子的需求相異,表明了研究區域的附生蘭較高的分化程度。
植被類型本身是特定區域的各類植物與諸多環境因子、植物之間長期相互作用的結果,附生蘭的存在即表明其在當前的植被類型中占據一定的生態位,因而植被類型的變化能夠很好的反映附生蘭在物種組成上的變異。據筆者觀察,植被類型往往比其他環境因子具更高的解釋率,如熱帶針葉林中的黑毛石斛+樂東石豆蘭+伏生石豆蘭組合、低地雨林中的大序隔距蘭+純色萬代蘭+窄唇蜘蛛蘭組合等,亦有相關研究支持了此觀點[21]。區別于眾多地生植物所依賴的土壤,宿主樹皮是附生蘭著生的載體,其對附生蘭的生長發育的作用至今尚未完全明晰[11,20],但不難推斷的是,樹皮的表面特征決定了附生蘭附著及種子萌發的難易程度[5]。本研究結果表明,植被類型為影響附生蘭物種組成最重要的因子,在所選的9 個環境因子中,植被類型對蘭科植物物種組成變異的解釋率最高并且也達到了高的顯著水平,其次則為樹皮類型。
3.2 附生蘭物種豐富度及多度與地形因子、氣候因子及生物因子的關系對于熱帶蘭科植物的多數研究表明,其垂直分布格局為“中間膨脹型”[15?16,19]。本研究結果顯示,鸚哥嶺附生蘭在中低海拔段(700~900 m)有著最高物種豐富度,這與許多學者的研究一致,其原因可能是低海拔地區人為干擾強度較大,附生蘭和其生境易遭破壞,而高海拔地區極端的環境條件不利于多數附生蘭的生長。少數適應于高海拔濕冷條件的附生蘭如黑毛石斛、伏生石豆蘭、流蘇貝母蘭等,由于競爭小,環境條件穩定且極少受到人類活動干擾,因而個體數量龐大,在高海拔地區占有絕對的優勢。坡度對不同類型的植物分布具有不同的影響[32],對于遠離土壤的附生蘭而言,其產生的作用則更加微妙復雜。本研究結果表明,緩坡地段,附生蘭物種豐富度較高,而物種多度的變化在坡度梯度上呈現出先升高后降低的趨勢。可能的原因是,緩坡地段土壤肥沃且排水良好,有利于高大粗壯宿主樹的長成,可為附生蘭提供更多的生長環境。在山地區域,坡向的變化影響了環境的光照條件和水分的分配,從而間接影響了植物的分布。蘭科植物多為喜濕耐陰的草本植物。本研究結果顯示,附生蘭在陰坡與半陰坡的蘭科物種數和個體數量要多于陽坡與半陽坡,證明附生蘭對光照的低的需求度和對空氣濕度的高的需求。
氣候是影響植物分布的主導因素,表現在熱量是植物生命活動能量來源、水分是影響植物生理活動的因素和植物基本成分[33]。一般而言,附生型蘭科植物多產于熱帶或亞熱帶,可在較高溫度的環境下生長。在本研究中,附生蘭物種豐富度隨年平均氣溫的升高呈現先升高后降低的趨勢,在21.2~21.7 ℃段出現峰值,物種豐富度最高,年平均氣溫達到23.7 ℃以上時,附生蘭物種豐富度很低,類似的結論在同為中國熱帶地區的西雙版納的蘭科植物分布研究中亦見報道[17]。溫差可影響植物生理活動中有機物質的合成量、開花率[34]等,從而影響植物的分布與繁殖力。本研究表明,溫差在7.8~7.85 ℃之間的地段附生蘭物種豐富度較高,該研究結果對于當地附生蘭集中保護區域的選取具重要的參考意義。對于附生蘭而言,降雨影響著附生基質(樹皮)的含水量和林分的濕度。本研究得出,附生蘭物種豐富度隨最干月降雨量增多而先升高后降低,在最干月降雨量為14~15 mm 地段附生蘭物種豐富度最高,隨著最干月降雨量的增多,附生蘭多度呈現出遞增的趨勢,可見,多數附生蘭分布的適宜生境為降雨量較大的林區。
區域植被類型是一定區域內植物群落與地形、氣候、人為因素等長時間相互作用的結果,往往能綜合反映該區域的環境條件與植物特征。附生蘭是熱性林的代表成分之一,因而在位于熱帶邊緣的海南島,附生蘭物種豐富度一定程度上反映了該地段熱帶屬性的強弱[35?36]。在研究區域內,低地雨林中附生蘭物種豐富度最高,熱帶針葉林中附生蘭物種豐富度最低。該結果與劉廣福等[19]在海南霸王嶺對附生蘭在不同類型森林植被中的分布結論略有不同,原因可能是,盡管附生蘭的分布依賴于特定的植被類型,但在確定的植被類型內部,附生蘭生境小氣候很容易受到各種因素復雜的影響。實際上,不論是在哪種植被類型的林區,均存在有大片面積的地段中觀察不到附生蘭的情況,而附生蘭如灰塵般的種子的傳播能力是毋庸置疑的。因此,筆者認為,附生蘭對的生長對小氣候有著極高的要求,大多數環境難以滿足附生蘭種子萌發及幼苗生長的要求[37]。植被郁閉度一方面塑造了附生蘭的光照環境,另一方面影響了林分的空氣濕度。本研究得出,郁閉度較大的林區具有略高的附生蘭物種豐富度和個體數量,表明多數附生蘭對光照需求很低,這與其他地區對附生蘭的研究結論一致[17]。樹皮類型決定了附生蘭種子附著與萌發的難易程度,從而影響了附生蘭的分布。研究區域內附生蘭廣泛分布于粗糙樹皮上,而高海拔區域的覆蓋有厚苔蘚的樹皮上,往往分布有數目龐大的附生蘭,可見樹皮基質對于附生蘭的生長具有重要作用。樹皮可能包含諸多對附生蘭生長起重要作用的因素,如含水量、化學物質、微生物群落、pH 等等,相關的研究尚不深入[20, 38 ? 40]。
總的來說,自然環境中附生蘭的分布受多種生態因子的共同作用,其作用機制也十分復雜,在不同的環境中,附生蘭對各單一的生態因子的需求也有所變化。植被類型較大程度包含了附生蘭分布的生境需求,實際上也代表了各個環境因子的適宜范圍。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
中老年保健(2021年12期)2021-08-24 03:30:40
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
中國生殖健康(2020年6期)2020-02-01 06:28:50
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
中國生殖健康(2019年11期)2019-01-07 01:28:02
汽車觀察(2018年10期)2018-11-06 07:05:26
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
