谷子SHR 基因的全基因組鑒定與表達分析
2022-06-16 15:01:14李露露李艷芬尹美強原向陽溫銀元
山西農業科學 2022年6期
關鍵詞:分析
李露露,李艷芬,尹美強,趙 娟,原向陽,溫銀元
(山西農業大學 農學院,山西 太谷 030801)
谷子(Setaria italic)因其根系發達、次生根生長數量多、密集、龐大,且具有耐瘠薄、耐干旱,適應性強等特點[1-2],深入開展對谷子根系的研究對提高谷子的抗倒伏能力以及抗旱保苗能力至關重要,同時對于谷子的高產栽培也具有重要意義。
研究發現,GRAS 家族蛋白是植物特有的一類轉錄因子,分為8 個亞家族:SCL、LISCL、SHR、PAT1、DELLA、SCR、LS、HAM,參與植物的信號轉導、根和腋芽的發育、根瘤和菌根的形成、生長發育、信號轉導、解毒作用、脅迫相關的應答過程[3]。其中,SHORT-ROOT基因(SHR)對根的輻射模式和根頂端分生組織的形成至關重要[4-5]。BENFEY等[6]于1993 年首次發現SHR基因,它與擬南芥根的伸長生長和輻射形態形成有關。2000 年,擬南芥的SHORT-ROOT基因(AtSHR)被成功克隆,其編碼區全長1 593 bp,在根和莖中都有轉錄產物[7]。shr突變體的凱氏帶結構消失[8],內皮層缺失,只保留有皮層,表現出短根、不定根增多的表型[9-10]。THOMAS 等[11]研究表明,SHR基因可作為細胞增殖的調節因子,影響玉米葉片維管組織和花環結構的形成而調控葉片的發育。此外,SHR基因還介導植物生長發育過程中許多重要的信號途徑[12-13],參與生長素的極性運輸(PIN3、PIN7、CYP79B2和SPS等12 個基因)[14-15]、側根和不定根的發生[16]、根尖分生組織干細胞的維持[17]、GA 信號轉導途徑(GA3、SNE、RGL1 和RGL2基因)[18]和油菜素內酯的生物合成、信號轉導和感應(BIN3、BRL3、Br6ox2和CYP90D1基因)[19-20]等多種代謝過程。
目前,已在多種植物如水稻[21]、甘藍型油菜[22]、白楊[23]和蒺藜苜蓿[24]等植物中發現了SHR基因,但在谷子中關于SHR基因的特征與表達模式的研究尚未見報道。
本研究參照AtSHR基因,在NCBI和Phytozome數據庫中篩選谷子的SHR基因,對其加以鑒定和生物信息學分析,并對SHR基因在谷子不同生育時期各組織中的表達模式進行分析,旨在為進一步研究谷子SHR 蛋白的功能、作用途徑和谷子根的生長發育及其輻射形態的形成提供參考依據。
1 材料和方法
1.1 谷子SHR 基因的鑒定和基因信息挖掘
以AtSHR基因(AT4G37650)的cDNA 序列為基礎,在NCBI、Phytozome 上對谷子全基因組進行BLASTn 比對,獲得SiSHR基因,下載擬南芥、水稻和谷子SHR基因的全基因序列、CDS 序列和蛋白序列用于后續生物信息學分析。利用已獲得的SHR基因的全基因序列和CDS 序列在GSDS 在線工具中繪制谷子SHR基因、擬南芥和水稻SHR基因的基因結構圖[25];用MEME 4.10.2(http://memesuite.org/tools/meme)在線軟件進行motif 分析[26]。并用MEGA 7.0 軟件構建SHR基因的系統發育樹,分析其進化關系。用SHR基因轉錄起始位置(ATG)上游2 000 bp 的序列,利用PlantCARE 在線數據庫進行啟動子順式作用元件分析。
1.2 SiSHR 蛋白的理化性質分析和空間結構預測
用ProtParam(http://web.expasy.org/protparam/)[27]分析谷子SHR 蛋白的理化性質和氨基酸組成,并用ProtScale 分析其疏水性。采用在線工具PRABI(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl? page=/NPSA/npsa_sopma.html)對其二級結構進行預測;谷子和擬南芥SHR 蛋白的三級結構利用在線軟件SWISS-MODEL(https://swissmodel.expasy.org/interactive)預測。
1.3 SiSHR 基因在谷子中的組織特異性表達分析
以晉谷42 號為材料,分別在苗期和灌漿期取谷子根、莖、葉、葉鞘、穗頸、穗共6 個組織,液氮速凍,保存于-80 ℃超低溫冰箱中。
使用Primer Premier 5.0 設計引物(表1),以谷子Actin基因作為內參基因,引物由上海生工生物有限公司合成。采用改良RNSiso Plus 法[28]提取總RNA 并反轉錄為cDNA,稀釋3 倍于-20 ℃保存。qRT-PCR使用SYBR?PremixExTaqⅡ(TliRNaseH Plus)試劑盒(TaKaRa)完成。采用2-ΔΔCt計算相對表達量,每個樣品3 個重復。

表1 SHR 蛋白引物序列Tab.1 Primer sequence of SHR proteins
2 結果與分析
2.1 SiSHR 基因的鑒定和生物信息學分析
以AtSHR的cDNA 序列為模板,在NCBI、Phytozome 上對谷子全基因組進行BLASTn 比對,獲得了2 個SiSHR基因,分別命名為SiSHR1(Seita.9G355300)、SiSHR2(Seita.2G372300),其中SiSHR1基因的CDS 長度為1 908 bp,編碼635 個氨基酸,位于谷子的9 號染色體上;SiSHR2基因的CDS 長度為1 785 bp,編碼594 個氨基酸,位于谷子的2 號染色體上。
用GSDS 繪制SHR基因的基因結構圖如圖1所示,與水稻和擬南芥相比,SiSHR基因在結構上基本一致,SiSHR基因與水稻和擬南芥的SHR基因都只有1 個外顯子,沒有內含子。SHR基因的基因結構物種間差異不大。
系統進化分析結果表明,SiSHR基因與OsSHR基因在進化上同源性較高(圖2-A)。SiSHR1基因與OsSHR1基因聚為一類,SiSHR2基因與OsSHR2基因聚為一類,擬南芥、煙草(A0A1S4A5 E5 TOBAC)、辣 椒(A0A2G3AB65 CAPAN)的SHR基因聚為一類。motif 分析發現了10 個保守基序,依次命名為motif 1~motif 10(圖2-B)。其中,motif 6 位于氨基酸序列中部,為VHIID 保守基序,motif 9 為PFYRE 保守基 序,motif 10 為SAW保守基序(圖2-C)。SHR 蛋白在結構上總體保守。

圖2 SHR 蛋白的系統進化分析與motif 分析Fig.2 Phylogenetic analysis and motif analysis of SHR proteins
2.2 SiSHR 基因的啟動子區順式作用元件分析
為認識SiSHR基因應答各種反應的潛在機制,分析了其啟動子區順式作用元件,結果如圖3所示,2 個SiSHR基因的啟動子區均含有光信號、激素(ABA、生長素、MeJA)、脅迫(低溫)、分生組織表達、醇溶蛋白代謝調控相關元件和MYBHv1 轉錄因子結合位點。除此之外,SiSHR1特有晝夜節律控制響應元件和GA 代謝響應元件,SiSHR2特有細胞周期調控元件和水楊酸響應元件。存在的這些調控元件說明SiSHR基因家族成員不僅能夠響應光、脅迫等外界環境信號,而且還能應答多種激素信號,調控谷子的生長發育。
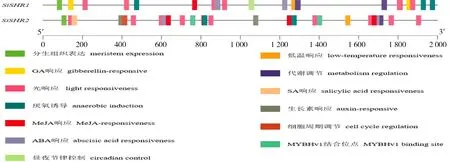
圖3 SiSHR 基因啟動子區域順式作用元件分析Fig.3 Cis-acting elements analysis in SiSHR gene promoter region
2.3 SiSHR 蛋白的理化性質和二、三級結構分析
谷子SHR 蛋白的理化性質分析結果表明(表2),SiSHR1 蛋白的分子式為C2933H4485N877O943S15,分子質量為67 601.09 u,其中絲氨酸(Ser,14.0%)含量最高;SiSHR2 蛋白的分子式為C2784H4299N817O899S16,分子質量為64 111.65 u,其中絲氨酸(Ser,13.3%)與丙氨酸(Ala,13.3%)含量最高;SiSHR1 蛋白和SiSHR2 蛋白的等電點均小于7,不穩定系數均大于40,平均親水性指數均小于0,說明SiSHR1 和SiSHR2 均為酸性不穩定的親水蛋白。ProtScale 疏水性分析結果也表明,SiSHR 是親水蛋白(圖4)。

表2 谷子SHR 蛋白的理化性質Tab.2 Physical and chemical properties of SiSHR protein in Setaria italica

圖4 SiSHR 蛋白的疏水性預測Fig.4 Hydrophobicity prediction of SiSHR proteins
PRABI 對SiSHR 蛋白的二級結構預測結果如圖5 所示,主要由α-螺旋和無規則卷曲組成,約占85%。對SiSHR 蛋白進行三級結構的建模發現(圖6),谷子SHR 蛋白空間構象(包括二級結構的走向和空間折疊方向)與擬南芥的十分相似,二者均模擬到5b3g.1 模型,為SHORT-ROOT 蛋白模型。

圖5 SiSHR 蛋白二級結構預測Fig.5 Secondary structure prediction of SiSHR proteins

圖6 SHR 蛋白三級結構預測Fig.6 Tertiary structure prediction of SHR proteins
2.4 SiSHR 蛋白互作分析
為探究谷子中SHR 蛋白在谷子生長發育過程中的作用,對SiSHR 蛋白的互作情況進行預測,結果發現,SiSHR1 和SiSHR2 均為根形態建成的核心作用因子(圖7),均可與5 個蛋白互作,CYCLIN家族蛋白(Si032222m)調控細胞周期,C2H2 轉錄因子蛋白(Si005031m)調控分生組織的形成,GRAS 轉錄因子家族的SCL 蛋白(Si032551m)調控根的生長發育,SHBY 家族蛋白(Si013112m)調控細胞分裂,WOX 家族蛋白(Si002900m)調控根尖的生長,Ser/Thr 蛋白激酶(Si016636m)與植物生長發育過程中的多種途徑有關。其中,SHR 蛋白與SCL 蛋白以共表達方式進行互作,其他蛋白與SHR 蛋白具體的互作情況還有待進一步試驗驗證。

圖7 SiSHR 蛋白互作分析Fig.7 Interaction analysis of SiSHR proteins
2.5 SiSHR 基因的時空表達模式分析
以晉谷42 號的不同組織為材料,分析SHR基因在谷子中的時空表達情況,結果發現(圖8),SiSHR1 和SiSHR2這2 個基因在谷子不同生育時期存在不同程度的表達,呈現出時空特異性。這2個基因在萌發3 d 的種子中都有一定的表達;在苗期,2 個SHR基因在根、莖、葉中均有表達,其中根中表達量最高,在莖中表達量最低;在灌漿期,2 個基因在根中高表達,在其余組織中表達量則很低。說明SHR基因在谷子根系的形態建成過程中有重要作用,同時也參與谷子苗期葉的生長。除根以外的其他組織中,SiSHR1的表達量均低于SiSHR2,且除根和葉以外的其他組織中SiSHR1的表達量很低,說明SiSHR基因對谷子的萌發、葉片發育及根的形態建成由SiSHR1、SiSHR2共同調控,而對其他方面的調控主要是SiSHR2在起作用。

圖8 SiSHR1/SiSHR2 基因的組織特異性表達Fig.8 Tissue-specific expression of SiSHR1/SiSHR2 genes
3 結論與討論
谷子有發達的根系,次生根密集。SHR基因介導側根和不定根的發生、維管束細胞的發育以及根尖分生組織的生長,對根的形態構成有重要作用[5,12,16-17]。研究發現,不同物種中存在SHR基因數目的不同,擬南芥中有1 個[6],水稻中有2 個[21],在谷子基因組中鑒定到2 個SHR基因,編碼的蛋白含有VHIID 結構域和PFYYE 結構域,它們是SHR蛋白保持活性的必要結構域,SHR 蛋白在VHIID結構域與其他轉錄因子結合發揮作用[29]。
有研究發現,GA 信號與SHR 共同調控擬南芥根的形成[30];SHR 和SCR 可以促進PIN 蛋白的表達和極性分布,調控根尖生長素轉運[31];生長素響應因子(ARFs)調控SHR 由維管組織向基本組織移動[32],同時它可以促進生長素從胚芽向胚根運輸,為胚根提供生長素的位置信號[33]。在側根原基形成過程中,SHR 啟動子介導mIAA14-GR 影響側根原基中的生長素信號,抑制側根的形成[34]。本研究中,SiSHR基因的啟動子區含有GA、生長素、水楊酸和ABA 信號響應元件,可能通過調控GA、生長素、水楊酸和ABA 信號途徑參與調控谷子根的形態建成;蛋白互作預測結果表明,SiSHR 蛋白與SCL 蛋白的互作方式為共表達,與擬南芥中SHR基因與SCR基因和SCL基因常形成SHR-SCRSCL3 復合體共同影響根輻射的形態形成[35]的研究結果一致。
有研究發現,SHR基因缺失導致植物出現短根的形態[30],在本研究中,SiSHR1/SiSHR2基因在谷子的苗期和灌漿期在根中均有相對較高的表達量,且在苗期根中的表達量高于灌漿期的表達量,由此推測,SiSHR基因在谷子中有調控根系形成的作用。SHR的過表達促進葉脈組織形成,zmshr1突變體的維管組織、葉組織發育不正常(包括葉肉細胞和維管鞘細胞)[11];擬南芥SHR功能缺失抑制細胞分裂、擾亂細胞核內復制過程,導致細胞增殖過程提前停止[36],shr突變體和scr突變體的蓮座葉生長受到抑制[37];OsSHR2在發育的小葉脈中表達,促進氣孔分化,但OsSHR1不表達[38];SiSHR1在葉片中的表達量低于SiSHR2,與水稻中一致,SiSHR基因在谷子苗期葉片中的相對表達量遠高于灌漿期成熟的葉片中的表達量,表明SiSHR基因可能調控谷子葉片的發育。由此推測,谷子的萌發、葉片發育及根的形態建成由SiSHR1、SiSHR2共同調控,而對其他方面的調控主要是SiSHR2在起作用。
本研究從谷子中鑒定出2 個SHR基因,其中,SiSHR1基因位于9 號染色體,編碼635 個氨基酸;SiSHR2基因位于2 號染色體,編碼594 個氨基酸。SiSHR 蛋白均為不穩定的酸性親水性蛋白,二級結構主要由α-螺旋和無規則卷曲組成,三級結構預測與SHORT-ROOT 蛋白模型(5b3g.1)相似。值得注意的是,SiSHR啟動子區含有光、激素(GA、ABA、生長素、水楊酸)等響應元件,SHR 蛋白與CYCLIN、C2H2、SCL、SHBY、WOX 蛋白互作,其中SiSHR 蛋白與SCL 蛋白的互作方式為共表達,故推測SiSHR 蛋白可能通過應答GA、生長素和ABA 信號途徑,及蛋白互作的方式參與調控。進一步對晉谷42 號中SiSHR基因在不同時期組織中的表達進行差異分析,結果發現,SiSHR1/SiSHR2基因在根中的表達量最高;更重要的是,苗期根和葉片中的表達量均顯著高于灌漿期根和葉片中表達量,表明SiSHR基因的表達參與調控谷子根的形態建成和葉片發育,為谷子SiSHR基因在根和葉片中的調控機制研究奠定了理論基礎。
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
