肝癌細胞SMMC-7721中CXCL3表達與氧化應激的相關性研究
2022-06-16 02:34:30毛麟宇鹿士振王偉群張春斌張麗娜
腫瘤基礎與臨床 2022年2期
毛麟宇,鹿士振,王偉群,張春斌,張麗娜
(1.佳木斯大學附屬第一醫院檢驗科,黑龍江 佳木斯154003;2.佳木斯大學基礎醫學院生物學教研室,黑龍江 佳木斯154007;3.佳木斯大學基礎醫學院生理學教研室,黑龍江 佳木斯154007;4.漳州衛生職業學院醫學技術系、轉化醫學檢測應用技術協同創新中心,福建 漳州 363000;5.大慶油田總醫院檢驗科,黑龍江 大慶 163000)
肝癌是我國常見的惡性腫瘤之一,具體發病分子機制尚有待研究。相關研究稱感染性疾病和慢性炎癥約占致癌因素的25%。慢性炎癥時炎癥細胞和上皮細胞均可產生反應性氧(reactive oxygen species,ROS)或反應性氮(reactive nitrogen species,RNS),引起DNA的損傷、突變,進而可能引發惡性腫瘤的發生。而乙肝、丙肝導致的慢性肝炎的長期不愈,反復發作,將導致肝臟的病變:肝纖維化及肝硬化,并最終發展為肝癌。趨化因子3(C-X-C motif chemokine ligand 3,CXCL3)作為炎性細胞因子的一種,在肝細胞癌中保持較高的表達水平,正符合惡性腫瘤發生發展的炎性微環境理論。
長期炎癥狀態下,氧化應激失衡,ROS對核酸、蛋白、脂質等的氧化作用促進了惡性腫瘤的發生,損傷的DNA進入細胞質后,又會誘導干擾素介導的先天免疫反應,從而促進ROS的產生,保持著腫瘤的炎癥環境。適量的ROS在惡性腫瘤的進程中激活了對細胞有利的信號,可以促進細胞的增殖,存活和抗氧化應激能力。而過高的ROS會導致腫瘤細胞的持續突變和基因組不穩定性。癌癥的發展中也需要依賴于內源性抗氧化劑來減輕氧化應激,使其保持在一定的水平。
有相關研究發現CXCL3在肝細胞癌中表達高于癌旁組織且跟腫瘤的炎癥環境相關[1-2],但關于肝癌中CXCL3和氧化應激之間的關系尚未有人研究。本課題的目的在于研究CXCL3表達水平發生改變后,細胞氧化應激水平變化情況,驗證在肝癌細胞中CXCL3與氧化應激水平的相關性。
1 材料與方法
1.1 實驗材料肝癌細胞系來源于美國模式培養物集存庫;CXCL3病毒轉染試劑購自廣州易錦生物技術有限公司;BeyoRTTMⅡ cDNA第一鏈合成試劑盒、RIPA裂解液(強)購自中國碧云天生物技術有限公司;總RNA提取試劑購自生工生物工程(上海)股份有限公司;熒光定量試劑購自羅氏診斷產品(上海)有限公司公司;血紅素加氧酶1(heme oxygenase-1,HO-1)抗體、總超氧化物歧化酶(superoxide dismutase,SOD)測試盒(羥胺法)、丙二醛(malondialdehyde,MDA)測定試劑盒(硫代巴比妥酸法)購自萬類生物有限公司;辣根過氧化物酶標記二抗購自北京中杉金橋生物技術有限公司。
1.2 細胞轉染對數生長期的SMMC-7721細胞按5×104個/mL的濃度鋪入24孔板中;將載有CXCL3高表達和CXCL3敲除及各自對照的帶有嘌呤霉素抗性及相應熒光蛋白質粒的慢病毒顆粒與完全培養基按11混合;按轉染試劑操作說明轉染到SMMC-7721細胞中(依次命名為:vector-CXCL3和sh-CXCL3);培養12~16 h后,加入嘌呤霉素(終濃度為2 μg/mL)進行篩選;純度達到90%以上后,進行后續實驗。
1.3 細胞增殖細胞按1.5×103個/孔接種于96孔培養板中;細胞在37 ℃培養24 h、48 h和72 h后,分別向設定孔中加入10 μL CCK-8試劑;37 ℃培養箱中孵育1 h后用酶標儀讀取各孔450 nm處的吸光度值。
1.4 Western Blotting法將30 μg的細胞總蛋白經質量分數12%SDS聚丙烯酰胺凝膠電泳處理后,轉移至聚偏氟乙烯膜;質量分數5%脫脂乳封閉后,用兔抗HO-1多克隆抗體(11 000稀釋)在4 ℃孵育過夜;TBST洗滌3次×5 min;辣根過氧化物酶偶聯的山羊抗兔二抗(15 000稀釋)在37 ℃孵育1 h;TBST洗滌4次×10 min;ECL試劑孵育膜,使信號可視化;用全自動化學發光圖像系統進行曝光、拍照,并對圖像進行量化和分析。
1.5 氧化應激處理將等量細胞鋪到6孔板的4個孔,待細胞長滿80%時按最終為0、100、200、400 μmol/L的濃度梯度將過氧化氫加入6孔板,正常培養24 h,提取4個孔的總RNA,反轉錄,熒光定量檢測各自的CXCL3的表達量。
1.6 SOD、MDA測定SOD、MDA的測定采用酶聯免疫吸附試驗,具體操作按照說明書進行。
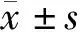
2 結果
2.1 CXCL3的表達情況本研究通過對TCGA數據庫的生物信息學分析發現,CXCL3在肝癌組織的轉錄水平與正常組織比較差異無統計學意義(t=0.864,P=0.075)。CXCL3高表達患者與CXCL3低表達患者相比,總生存率低(P=0.001)。見圖1、2。

圖1 TCGA數據庫中CXCL3在不同肝臟組織中表達的比較
2.2 CXCL3與氧化應激的關聯分析
2.2.1 過氧化氫處理后CXCL3的表達 過氧化氫濃度為100 μmol/L時,CXCL3的表達與對照組差異無統計學意義(t=0.716,P=0.494)。當濃度達到200 μmol/L時,CXCL3的表達下降(t=3.275,P=0.004)。當濃度達到400 μmol/L時,CXCL3的表達水平下降更加明顯(t=5.162,P<0.001)。見圖3。
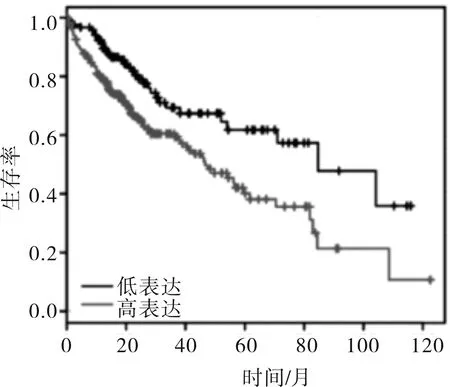
圖2 CXCL3表達對肝癌患者生存率的影響
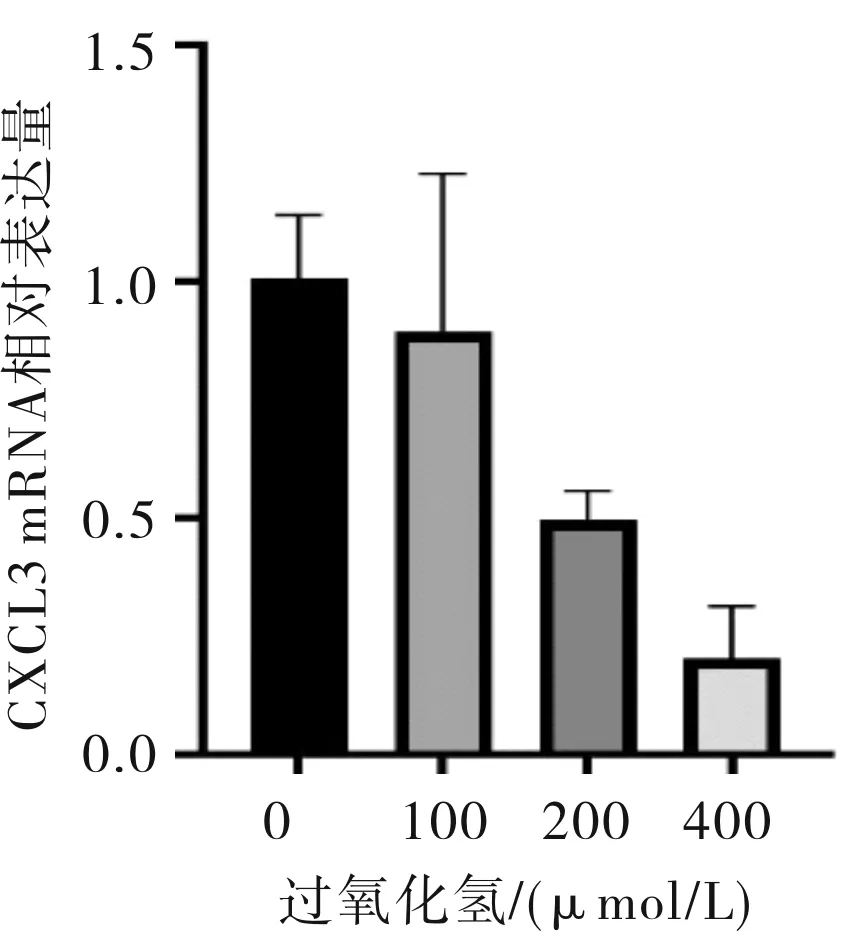
圖3 過氧化氫處理后肝癌細胞中CXCL3表達情況分析
2.2.2 CXCL3與氧化應激基因的關聯分析 生物信息學分析結果所示,肝癌組織中有7個與CXCL3表達相關的氧化應激基因。包括1個負相關基因PTGS2(COX2);6個正相關基因,分別為CYBB、SOD3、NCF1、NCF2、UCP2和HMOX1。見圖4。
圖4 與CXCL3表達相關的氧化應激基因及其相關系數
2.3 熒光定量PCR驗證轉染效果為了驗證CXCL3表達對肝癌細胞氧化應激狀態的影響,本研究設計了高表達CXCL3細胞和低表達CXCL3細胞。穩定轉染CXCL3的SMMC-7721細胞中CXCL3的表達接近其對照的2倍(t=3.432,P=0.034);RNAi干擾SMMC-7721細胞的CXCL3表達是對照組的20%左右(t=9.957,P=0.014)。見圖5。
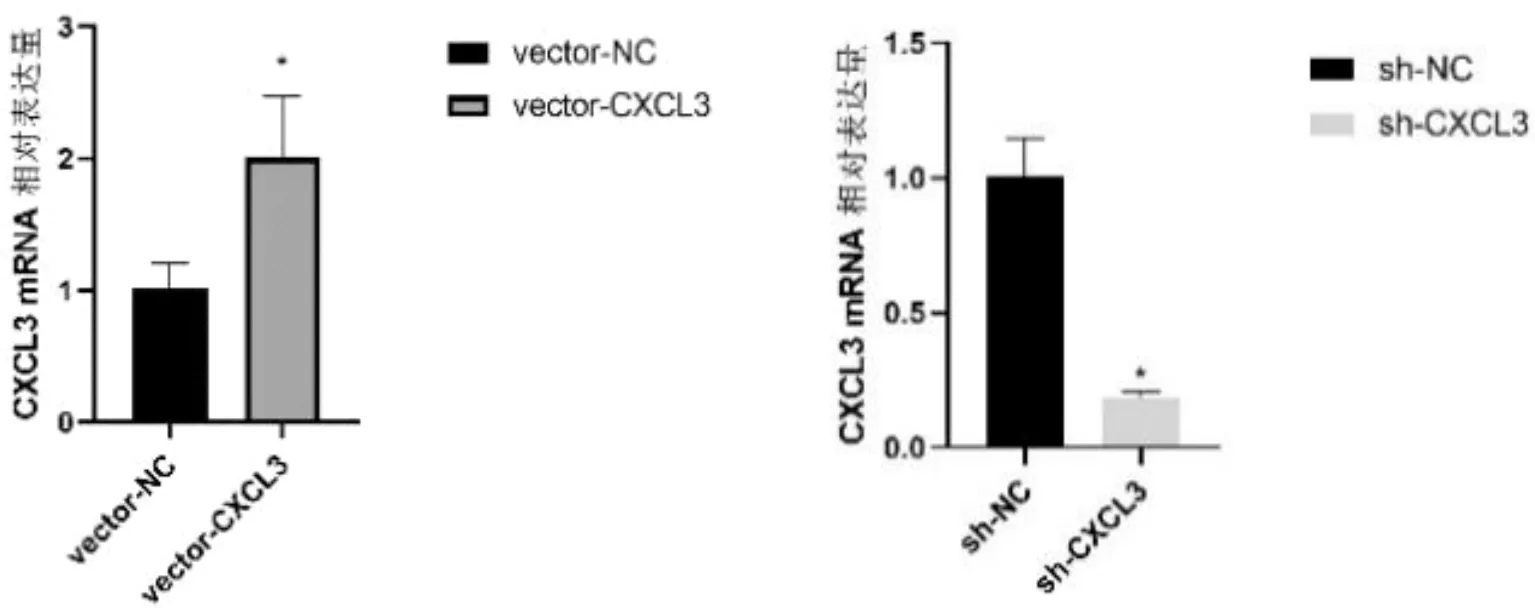
圖5 轉染后SMMC-7721細胞中CXCL3表達情況
2.4 CCK-8增殖情況SMMC-7721細胞中,穩定高表達CXCL3的細胞增殖能力強于對照(t=4.180,P<0.001);低表達CXCL3的細胞增殖能力低于對照組(t=60.201,P<0.001)。見圖6。
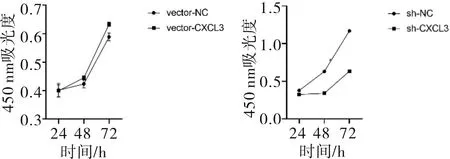
圖6 CXCL3表達對SMMC-7721細胞增殖的影響
2.5 SOD測定結果SMMC-7721細胞中,高表達CXCL3的細胞SOD水平高于對照組(t=253.301,P<0.001);低表達CXCL3的細胞SOD水平低于對照組(t=253.782,P<0.001)。見圖7。
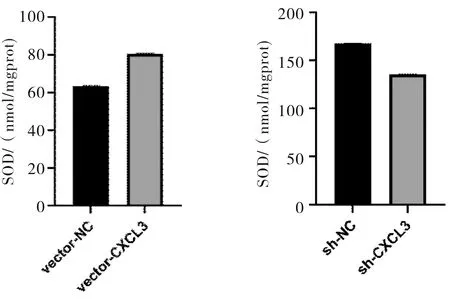
圖7 CXCL3表達對SMMC-7721細胞中SOD水平的影響
2.6 MDA測定結果SMMC-7721細胞中,高表達CXCL3的細胞MDA含量高于對照組(t=19.485,P<0.001);低表達CXCL3的細胞MDA含量低于對照組(t=11.265,P<0.001)。見圖8。
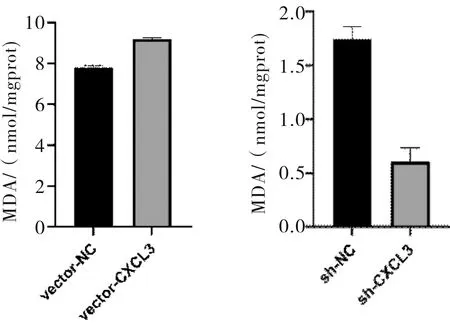
圖8 CXCL3表達對SMMC-7721細胞MDA水平的影響
2.7 HO-1表達情況SMMC-7721細胞中,高表達CXCL3的細胞HO-1相對表達量高于對照組(t=5.592,P<0.001);低表達CXCL3的細胞HO-1相對表達量低于對照(t=13.064,P<0.001)。見圖9、10。
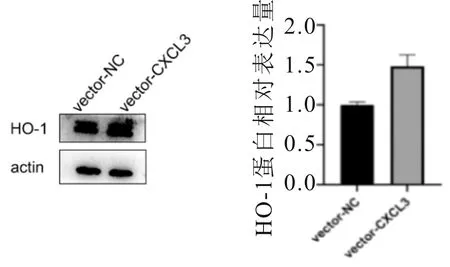
圖9 高表達CXCL3 SMMC-7721細胞中HO-1表達情況
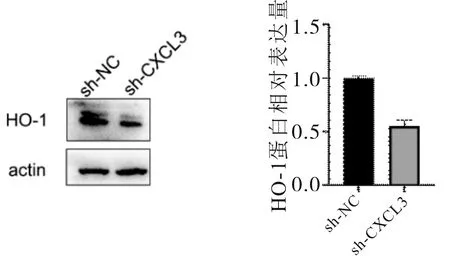
圖10 低表達CXCL3 SMMC-7721細胞中HO-1表達情況
3 討論
3.1 CXCL3的表達水平影響肝癌的惡性表型CXCL3是一種生長因子蛋白,與CXCR2結合,并作為中性粒細胞的趨化劑。作為一種重要的趨化因子,CXCL3在多種腫瘤中有一定的表達。在宮頸癌中,腫瘤組織高表達CXCL3[3-4];CXCL3通過調節細胞外調節蛋白激酶通路相關基因的表達促進宮頸癌細胞增殖和遷移、抑制其凋亡[5]。
在研究頭頸部癌時發現,CXCL3在口腔鱗癌組織中的表達高于正常組織,與腫瘤分期及淋巴結轉移密切相關;CXCL3的高表達促進了裸鼠移植瘤的生長;細胞外調節蛋白激酶阻斷劑PD98059可減弱CXCL3誘導的增殖和遷移效應[6]。CXCL3在頭頸部鱗癌中的表達高于正常患者,CXCL3高表達患者組總生存率較低,其表達水平會影響凋亡、Toll樣受體、Nod樣等多種信號通路基因[7]。
Han等[2]建立的裸鼠模型中,與相鄰正常組織相比,肝癌組織中CXCL3的mRNA和蛋白表達上調。同樣,Zhang等[8]報道CXCL3的高表達表明預后不良,通過細胞外調節蛋白激酶的磷酸化增強了肝癌干細胞的維持和腫瘤發生。
本研究通過對TCGA數據庫的生物信息學分析發現CXCL3在肝癌組織的轉錄水平高于正常組織。高表達CXCL3的患者比低表達CXCL3患者總生存率低。這與Wang等[9]的數據分析結果一致,說明CXCL3的表達水平也會影響肝癌的發病進程。
3.2 CXCL3與氧化應激基因的關聯分析有研究[10]表明,氧化應激的失衡可以加速細胞惡性衍變過程,并且影響腫瘤預后的效果。但是,迄今為止,仍不清楚腫瘤細胞中氧化應激分子調控及其作用機制的完整網絡,影響肝癌細胞中的氧化應激失衡的關鍵分子也不清楚。
我們先利用過氧化氫誘導肝癌細胞SMMC-7721,使其處于氧化應激狀態,來檢測CXCL3的表達情況。結果顯示,過氧化氫濃度達到200、400 μmol/L時,CXCL3的表達水平下降,并且隨著過氧化氫濃度增加,下降幅度逐漸增大。此結果可以說明,CXCL3的表達與肝癌細胞的氧化應激狀態密切相關。
為了探討肝癌細胞中的氧化應激相關基因表達情況,本研究利用cBioPortal數據庫,分析了肝癌細胞中與CXCL3基因共表達的氧化應激基因,利用DAVID數據庫(https://david-d.ncifcrf.gov/,6.7版本)對其進行了功能富集分析。結果顯示,當P<0.001時,在與CXCL3基因相關的基因功能注釋中,獲得了7個與氧化應激和腫瘤發生相關的基因,包括1個負相關基因:PTGS2(COX2);6個正相關基因:CYBB、SOD3、NCF1、NCF2、UCP2和HMOX1。
值得關注的是,與CXCL3負相關基因COX2,在大多數正常組織都未能找到。最近,其已被證明可在多種惡性腫瘤患者體內檢出,顯示在腫瘤病患中起重要作用[11-13]。這是一個誘導酶,功能是激活巨噬細胞或其他細胞,充斥于炎癥組織[14]。這與我們前期研究CXCL3影響炎癥、腫瘤的進程結果吻合。
3.3 CXCL3表達對肝癌細胞氧化應激狀態的影響為了驗證CXCL3表達水平對肝癌細胞氧化應激狀態的影響,我們設計了高表達CXCL3細胞株和低表達CXCL3細胞。
3.3.1 CXCL3表達對細胞增殖的影響 本研究中,高表達CXCL3的SMMC-7721細胞增殖能力強于對照組;低表達CXCL3的細胞增殖能力低于對照組。由此可以說明,CXCL3的表達水平與SMMC-7721細胞增殖密切相關,而氧化應激水平的改變很可能就是由增殖能力的變化引起的。
3.3.2 CXCL3表達對SOD的影響 SOD是機體內天然存在的超氧自由基清除因子,是一種能夠催化超氧化物通過歧化反應轉化為氧氣和過氧化氫的酶,而體內的其他酶會立即將過氧化氫分解為完全無害的水。因此,SOD能專一清除體內有害的自由基,以解除自由基氧化體內的某些組成成分而造成的機體損害。
本研究中,高表達CXCL3的SMMC-7721細胞SOD高于對照組;低表達CXCL3的細胞SOD低于對照組。這也與生物信息學預測中CXCL3與SOD基因呈正相關結果一致。
SOD在機體的氧化與抗氧化平衡中起到至關重要的作用。高表達CXCL3能引起SOD增加,這說明在一定范圍內增加CXCL3可以增強細胞的抗氧化能力,保護細胞免受氧化應激損傷。這與Wang等[15]發現人參皂苷抗氧化機制相類似。
3.3.3 CXCL3表達對MDA的影響 MDA是細胞膜脂過氧化作用的產物之一,其產生還能加劇膜的損傷。因此,MDA的多少能夠代表膜脂過氧化的程度。
本研究結果顯示,在SMMC-7721細胞中,高表達CXCL3的細胞MDA高于對照組;低表達CXCL3的細胞MDA低于對照組。由此可以說明,CXCL3的表達水平與SMMC-7721細胞氧化應激損傷密切相關。這與Li等[16]從枸杞中提取的枸杞多糖-4a對MDA的作用類似。
在很多研究中,SOD與MDA呈負相關,通常給以氧化應激刺激后,SOD降低而MDA升高。例如,檢測急性心肌梗死患者外周血[17]和噪聲暴露情況下大鼠耳蝸皮質組織[18]都屬于此類情況。但本研究中,高表達CXCL3的肝癌細胞中MDA升高可以說明細胞氧化應激狀態處于更高水平,而相對于CXCL3正常表達水平時,細胞長期處于增殖增加的狀態,這說明MDA等過氧化狀態對細胞損傷影響得以消除,即細胞的抗氧化能力的到了提高,SOD水平的升高正對應這一結論,反之亦然。
3.3.4 CXCL3表達對HO-1的影響 HO在細胞應激反應和體內平衡中起著關鍵作用。其表達或活性的失調與多種人類疾病的病理生理學有關,包括心血管疾病、糖尿病、肥胖、肺病、胃腸疾病、腎功能障礙、皮炎、癌癥等。因此,血紅素加氧酶被認為是許多人類疾病的預防、治療干預中最重要的目標之一[19]。
本研究結果顯示,在SMMC-7721細胞中,高表達CXCL3細胞的HO-1表達高于對照組;低表達CXCL3細胞HO-1表達低于對照組。由此可以說明,CXCL3的表達水平與HO-1表達密切相關,這與增加HO-1表達可能導致一些疾病的進展,如神經退行性變、致癌的觀點吻合[20]。
當前的惡性腫瘤相關研究已不再局限于進行某一因子的調控機制,而是強調整個的腫瘤微環境的作用。本課題根據惡性腫瘤的炎癥環境對惡性腫瘤發生、發展的影響,首次提出了在肝癌細胞中,CXCL3表達與細胞氧化應激水平具有相關性的觀點。但由于本課題只涉及了CXCL3與肝癌的相關性,沒有進行具體的調控關系及調控網絡的研究,且炎癥細胞因子僅選用了CXCL3,不能系統地詮釋炎癥相關因子與氧化應激的關系。這些問題將在課題組今后的研究中逐漸解決。
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
世界科學技術-中醫藥現代化(2020年2期)2020-07-25 02:05:56
天津醫科大學學報(2019年3期)2019-08-13 06:53:08
人大建設(2019年12期)2019-05-21 02:55:32
西南軍醫(2016年6期)2016-01-23 02:21:19
腫瘤預防與治療(2015年1期)2015-09-26 07:26:20
中國當代醫藥(2015年16期)2015-03-01 02:03:11
癌變·畸變·突變(2015年4期)2015-02-27 06:15:25
西南軍醫(2015年2期)2015-01-22 09:09:37
食品科學(2013年15期)2013-03-11 18:25:48
