PRL 和O2 對HC11 細胞氧化應激狀態下乳蛋白合成的影響
2022-06-20 10:26:56AlexanderJonathanSpitzerZHAOFengqi
中國飼料 2022年11期
關鍵詞:氧化應激
田 青 , Alexander Jonathan Spitzer, ZHAO Fengqi
(1.江蘇食品藥品職業技術學院,江蘇淮安223003;2.佛蒙特大學,伯靈頓05405)
氧化應激是促氧化物和抗氧化物之間的不平衡所致。Saeidnia 和Abdollahi(2013)研究表明,奶牛圍產期間隨著代謝需求增加, 尤其是氧氣需求的增加會導致活性氧(ROS)產量的增加,尤其是奶牛乳腺,作為乳汁合成的主要部位,其所承擔的代謝壓力更易發生氧化應激。 研究表明, 持續的氧化應激將誘導脂質過氧化, 繼而引起線粒體損傷,造成細胞凋亡,降低機體免疫功能和炎癥應答能力 (Ma 等,2018;Sordillolm 等,2005)。 Ma 等(2019)研究表明,氧化應激對免疫系統和炎性應答的影響可使奶牛對疾病的易感性增加,酮血癥、脂肪肝、卵巢疾病、低血鈣、低血鎂等患病率隨之升高。 研究表明, 奶牛分娩后乳腺代謝活躍而使高產奶牛乳腺產生大量的活性氧(ROS),而過量的ROS 會誘導隱性乳房炎的發生(Ma 等,2018;Mulligan 等,2008)。 同時,Ma 等(2018)研究還發現, 乳腺的密集生長和牛奶的合成也是乳腺產生氧化應激和炎癥的原因。 促炎作用的發揮則主要通過有絲分裂原激活蛋白激酶(MAPK)、絲氨酸/蘇氨酸激酶Akt (AKT) 和核因子kappa B (NFκB)介導,即過量的ROS 首先激活MAPK 和AKT信號通路,隨后NF-κβ 通路被激活,繼而引發炎癥因子的高表達, 炎癥反應程度和進程加重和延長,乳腺細胞開始受損傷,細胞開始凋亡,乳蛋白基因表達受阻, 乳品質變差, 甚至停乳(Abuelo等,2015;Liu 等,2012;Cargnello 等,2011)。 Ma 等(2018)指出,血紅素氧化酶-1(HO-1)和磷酸酰胺腺嘌呤二核苷酸醌氧化還原酶-1(NQO-1)是兩種重要的II 相解毒酶, 可促進細胞的抗氧化能力,硫氧還蛋白還原酶-1(TXNRD1)是一種重要的奶牛抗氧化硒蛋白,XCT 是一種控制細胞內谷脯甘化合成來源的重要轉運載體, 他們是參與機體抗氧化的關鍵基因, 這四種基因的表達受到Nrf2-ARE 的調控。 奶牛泌乳、代謝增強、氧化應激、炎癥損傷往往相伴而生,但又相互影響,甚至會影響奶牛產奶和乳品質, 給奶業生產帶來巨大損失。 氧化應激反應的發生既與代謝中氧氣需求增加有關,又與機體本身抗氧化能力有關,因此,
本文假設如果減少氧氣的供應, 或者激活機體抗氧化基因表達,就會對氧化應激、炎癥反應有一定的緩解作用。 本試驗旨在以體外培養的大鼠乳腺上皮細胞(HC11) 為載體, 探討催乳素和氧氣對HC11 細胞中參與抗氧化的關鍵基因表達的影響。
1 材料與方法
1.1 細胞模型 HC11 細胞來自于妊娠中期BALB/c 大鼠的細胞系,由美國佛蒙特大學提供。
1.2 試驗設計 采用隨機分組試驗設計,將細胞分為2 組,每組4 個處理,每個處理3 個重復,每個重復4 個孔, 研究有氧和缺氧狀態下催乳素對H2O2誘導HC11 細胞乳蛋白相關基因、 葡萄糖轉運因子和細胞因子基因表達的影響。 試驗設計方案見表1。
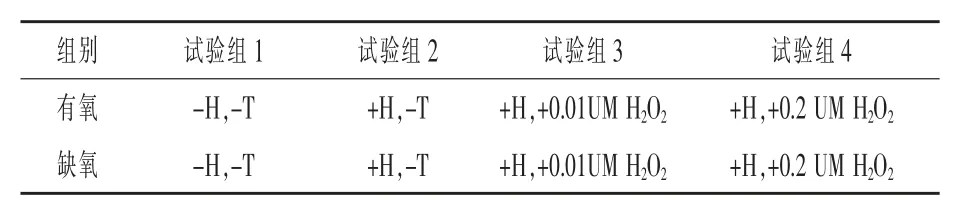
表1 試驗設計
1.3 試驗方法 復蘇后的HC11 細胞以2 ×105個/孔等密度接種于6 孔板, 先用生長培養基CGM(DMEM/F12,10% FBS,100 U/mL 雙抗,5 μg/mL 胰島素,10 ng/mL EGF,所有試劑均來自美國Gibco公司)培養至細胞80%貼壁后,從培養箱取出細胞,用PBS 清洗細胞3 次,換用前激素誘導培養基PIM (10% charcoal-treated 馬血清) 培養24 h后,再用PBS 清洗細胞3 次,再換用激素誘導培養基HIM(0.1 μmol 地塞米松,5 μg/mL 催乳素)分常氧組 (5% CO2培養箱) (0 μg/mL PRL+0 μmol H2O2,5 μg/mL PRL+0 μmol H2O2,5 μg/mL PRL+
0.01 μmol H2O2和5 μg/mL PRL+0.2 μmol H2O2)和缺氧組(缺氧培養箱,O2<6%)(0 μg/mL PRL+0 μmol H2O2,5 μg/mL PRL+0 μmol H2O2,5 μg/mL PRL+0.01 μmol H2O2和5 μg/mL PRL+0.2 μmol H2O2)分別培養3 h 后收獲細胞,用于測定各基因表達。
總RNA 的提取采用RNeasy Plus Kit (Qiagen)試劑盒并按說明書進行,RNA 的轉錄采用Super-ScriptTMIII Reverse Transcriptase (Invitrogen)試劑盒并按說明書進行,以GAPDH、β-actin 和HPRT為內參,采用SYBR green 試劑盒并按說明書對各基因mRNA 表達量進行分析,基因表達量用2-ΔΔCT法進行計算。 各個基因引物信息詳見表2。

表2 基因引物信息
1.4 數據統計分析 所有原始數據用SAS 9.1 中TUKEY 單因素方差分析進行數據處理,結果用“平均值±標準誤”表示。P<0.05 表示差異顯著。
2 試驗結果
2.1 有氧和缺氧狀態下催乳素對HC11 細胞蛋白質相關基因表達的影響 由表3 可見, 在有氧狀態下, 與對照組相比, 催乳素均顯著誘發了CSN2、CSN1S1 和α-LA 基因的表達(P< 0.05),且隨著H2O2濃度的增加, 三種蛋白基因的表達量顯著下降(P<0.05)。 在缺氧狀態下,催乳素顯著誘發了CSN2、CSN1S1 和α-LA 基因的表達 (P<0.05),隨著H2O2濃度的增加,CSN2、CSN1S1 基因的表達量顯著下降(P< 0.05)。但對α-LA 基因而言,隨著H2O2濃度的增加,其表達量反而顯著上升(P< 0.05)。 與有氧狀態相比,在缺氧狀態下,不管有無催乳素作用, 也不管H2O2濃度高低,CSN2和CSN1S1 基因表達量均顯著降低(P< 0.05),而α-LA 基因的表達量則顯著上升(P<0.05)。
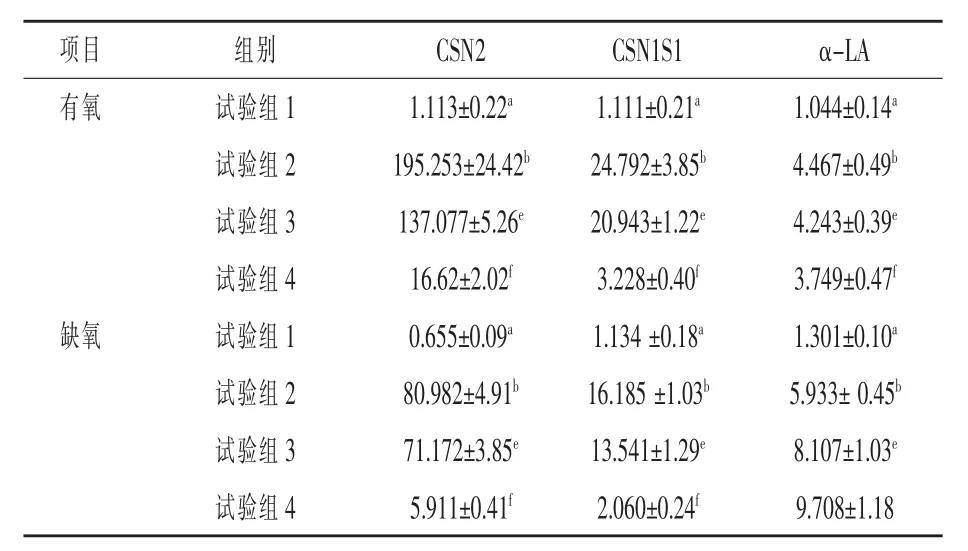
表3 有氧和缺氧狀態下PRL 對HC11 細胞乳蛋白相關基因表達的影響
2.2 有氧和缺氧狀態下催乳素對HC11 細胞抗氧化基因表達的影響 由表4 可見, 與對照組相比,在不使用H2O2的條件下,不管有氧還是缺氧,催乳素對抗氧化基因的表達影響差異不顯著(P>0.05)。 但在有氧且使用H2O2的條件下,與對照組相比,HO-1、XCT、TXNRD1 基因的表達量顯著增加, 且隨著H2O2劑量的增加而顯著增加(P<0.05);對NQO-1 基因,其表達量卻顯著降低(P<0.05), 隨著H2O2劑量的增加其表達量顯著減少(P< 0.05)。 在缺氧并使用H2O2的條件下,XCT、TXNRD1、NQO-1 和HO-1 基因的表達量與有氧狀態下的變化趨勢一致(P< 0.05),即在有催乳素作用的基礎上,與有氧狀態相比,缺氧顯著誘發了抗氧化基因XCT、TXNRD1 和NQO-1 的表達,并呈顯著的劑量效應(P< 0.05),但對TXNRD1 基因,其表達量的變化卻正好相反(P< 0.05)。

表4 有氧和缺氧狀態下PRL 對HC11 細胞氧化應激信號通路相關基因表達的影響
2.3 有氧和缺氧狀態下催乳素對HC11 細胞GLUTs 和炎癥因子基因表達的影響 由表5 可見,在有氧狀態下,催乳素對GLUTs 基因的表達影響差異不顯著(P> 0.05),卻顯著降低了炎癥因子IL-1β 的基因表達量(P< 0.05);當有氧并使用H2O2的條件下,GLUT8 基因的表達量顯著降低, 且有明顯的H2O2劑量效應 (P< 0.05),但GLUT1 基因表達變化不明顯(P> 0.05)。 對于炎癥因子,催乳素顯著降低了IL-1基因表達量,且其表達量隨著H2O2劑量的增加而增加 (P<0.05)。 在缺氧狀態下,催乳素顯著增加了葡萄糖轉運因子GLUT1 和GLUT8 基因的表達 (P<0.05), 也顯著增加了IL-1 基因的表達 (P<0.05)。 但在有氧并使用H2O2的條件下,低劑量的H2O2顯著增加了GLUT1 基因的表達量, 高劑量卻顯著降低了其表達量 (P< 0.05);H2O2均顯著增加了GLUT8 基因的表達量,且隨著劑量的增加而增加(P< 0.05)。 對于炎癥因子,H2O2顯著增加了IL-1基因表達量,且均隨著H2O2劑量的增加而增加(P< 0.05)。

表5 有氧狀態下PRL 對HC11 細胞GLUTs和炎癥因子基因表達的影響
3 分析與討論
3.1 有氧和缺氧狀態下催乳素對HC11 細胞蛋白質相關基因表達的影響 本試驗在有氧和缺氧狀態下采用不同濃度的H2O2對HC11 細胞誘導培養3 h,結果發現在有氧狀態下,與未經H2O2誘導組相比, 催乳素顯著誘發了CSN2、CSN1S1 和α-LA 基因的表達。當HC11 細胞被誘導處于氧化應激狀態下時,其CSN2、CSN1S1 和α-LA 基因的表達依然顯著高于未添加PRL 組,這說明催乳素在泌乳和乳蛋白合成方面的作用明顯。 但常氧狀態下隨著H2O2濃度的增加,三種蛋白基因的表達量也顯著下降, 說明氧化應激的確是奶牛養殖業和奶業生產中的一個重要問題, 雖然催乳素的存在對泌乳和乳蛋白的合成有一定的保護作用,但也不能完全抵擋氧化應激本身造成的損傷。 研究表明,PRL 是雌性動物乳腺生長發育、啟動和維持泌乳必不可少的激素。在乳腺中,PRL 發揮作用主要通過JAK/Stat 和PI3K-mTORC2 信號通路調控乳蛋白的合成(丁洛陽等,2016)。金亞亞等(2021)研究發現,100 ~ 400 μg/L 催乳素可以促進BMECs 增殖。200 μg/L 催乳素對BMECs 酪蛋白合成相關基因表達具有較好的促進效果。 100 μg/L催乳素在翻譯水平上對促進酪蛋白合成效果最好(Lee 等,2015)。 這和本試驗結果基本一致。 由此可見, 催乳素對乳腺蛋白質合成的積極作用是基本得到定論的。
泌乳期奶牛需氧量增大, 導致活性氧產量的增加,進而導致奶牛產生氧化應激的產生,而減少氧氣的攝入是否會緩解氧化應激反應所產生的氧化損傷鮮見報道, 因此本試驗同時開展了缺氧狀態下的研究。 研究表明,在缺氧狀態下,催乳素顯著誘發了CSN2、CSN1S1 和α-LA 基因的表達,但隨著H2O2濃度的增加,CSN2、CSN1S1 基因的表達量也顯著下降,這與有氧狀態下的趨勢一致。與有氧組相比,不管是在對照組還是催乳素誘導組,也不管是在低濃度還是高濃度的H2O2誘導組,其CSN2 和CSN1S1 基因表達量均顯著降低。 這說明, 缺氧狀態下催乳素雖然可以緩解氧化應激帶來的損傷,但是緩解的能力非常有限,氧氣不足更加劇了氧化應激造成的損傷, 對乳腺泌乳和乳蛋白合成的影響更大。 這一點可以由本試驗中得到的在缺氧和氧化損傷狀態下α-LA 基因的表達量的顯著上升來解釋,氧化損傷也嚴重,機體為了對抗損傷, 就需要動員諸如α-LA 等具有免疫功能的基因來參與損傷的修復, 進而盡可能的保持乳腺健康和正常泌乳功能。
3.2 有氧和缺氧狀態下催乳素對HC11 細胞氧化應激信號通路相關基因表達的影響 郭詠梅(2020)研究發現,圍產期和泌乳前期的高產奶牛代謝水平明顯增強,機體抗氧化功能顯著降低,很容易誘發氧化應激反應而導致疾病易感性增加,如乳腺炎、酸中毒、酮癥等。因此,除了泌乳激素調控, 如何動員機體的抗氧化基因來對抗氧化應激而維持機體的代謝平衡也很關鍵。
抗氧化系統主要包括酶類抗氧化劑和非酶類抗氧化劑, 兩者協同保護細胞免受自由基誘導的氧化損傷。 Lee 等(2015)研究表明,TrxR1 是細胞質中一種重要的硒蛋白,在氧化應激條件下,Nrf2快速地易位入核并與EpRE、ARE 結合,EpRE、ARE 存在于其目標抗氧化酶 (HO-1,NQO-1、GCLC 和TrxR1 等) 基因的啟動子區域并誘發其轉錄。本試驗研究表明,不管在有氧還是缺氧狀態下,催乳素對抗氧化基因的表達影響不大。但在氧化應激狀態下, 催乳素顯著增加了HO-1、XCT、TXNRD1 基因的表達量,且隨著H2O2劑量的增加而顯著增加。 說明當機體受到氧化應激損傷時,HO-1、XCT、TXNRD1 基因表達就會被激活,從而提高機體抗氧化應激的能力, 這與李前輝等(2021)的研究結果一致。
Abd 等(2021)試驗還發現,NQO-1 基因的表達量顯著降低,且隨著H2O2劑量的增加而顯著減少。 研究表明,NQO-1 參與抗氧化活性和應激下關鍵調控蛋白的穩定, 對于本試驗中NQO-1 基因的表達量顯著降低,且隨著H2O2劑量的增加而顯著減少的結果與前人的研究結果不太一致,其原因可能是氧化應激狀態造成了NQO-1 基因的損傷,也有可能與NQO-1 基因的多態性有關,同樣的矛盾結果也有報道。 據報道,NQO1 基因缺失,易使小鼠受到氧化應激的影響,發展為腫瘤;但也有報道稱NQO1 敲除可減少膠質母細胞瘤細胞的增殖, 而過度表達則增加細胞增殖(Dinkova等,2010)。除了酶活性、抗氧化活性和應激下關鍵調控蛋白的穩定作用之外,已有證據表明NQO-1在癌癥生物學中具有雙重效應, 既可以作為腫瘤抑制因子,也可以作為腫瘤促進因子,因此,產生此結果的具體原因還有待進一步研究。
本試驗研究結果還發現, 在缺氧并處于氧化應激狀態下,HC11 細胞中HO-1、XCT、TXNRD1和NQO-1 基因的表達量均顯著上調, 且隨著劑量的增加而顯著增加。在有催乳素作用的基礎上,與有氧狀態相比, 缺氧顯著誘發了抗氧化基因XCT、TXNRD1 和NQO-1 的表達, 并呈顯著的劑量效應,說明缺氧更加誘發了氧化應激損傷,為了對抗這種損傷,機體會動員抗氧化基因的表達,進而增強機體的抗氧化能力。
3.3 有氧或缺氧狀態下催乳素對HC11 細胞GLUTs 和炎癥因子基因表達的影響 本試驗研究結果表明, 在常氧狀態下, 催乳素顯著增加了GLUTs 基因的表達量,卻顯著降低了炎癥因子的基 因 表 達 量 (P< 0.05)。 H2O2均 顯 著 降 低 了GLUT8 基因的表達量(P< 0.05)。 葡萄糖轉運因子(GLUTs)是一類調控細胞外葡萄糖進入細胞內的跨膜蛋白家族,參與糖代謝、炎性反應和免疫應答等過程。 張青玲 (2020) 研究表明,GLUT1 和GLUT8 轉運蛋白在乳腺發育過程中表達增加,泌乳時GLUT1 mRNA 表達增加20 倍,GLUT8 蛋白表達增加2 倍。說明在未發生氧化應激的乳腺中,PRL 有利于增加葡萄糖的轉運能力而為乳腺提供能量,此時炎癥因子處于低表達狀態;而在機體產生氧化應激時,炎癥基因表達逐漸增強,說明氧化應激狀態下炎癥反應同步開始。本試驗中,常氧狀態下, 在存在PRL 的前提下,GLUT1 表達差異不顯著,GLUT8 在氧化應激狀態下表達量顯著降低;催乳素顯著降低了IL-1基因表達量,但隨著H2O2劑量的增加而增加,這也正好說明在氧化應激狀態下,葡萄糖的轉運受阻,炎癥反應加強,機體的主要機能由泌乳和乳成分合成向抗炎轉變。這一推測在缺氧狀態下表現的更為明顯。 本試驗結果中,在缺氧狀態下,催乳素顯著誘導了葡萄糖轉運因子GLUT1 和GLUT8 基因的表達, 低劑量的H2O2顯著增加了GLUTs 基因的表達量, 高劑量GLUTs 表達量與低劑量相比開始顯著降低;H2O2均顯著增加了GLUT8 基因的表達量, 且隨著劑量的增加而增加。 H2O2顯著增加了IL-1β 基因表達量,且均隨著H2O2劑量的增加而增加(P<0.05)。 這說明缺氧加劇了機體對抗炎癥時對能量的消耗,但隨著氧化應激的持續加劇,機體功能開始紊亂,炎癥反應持續增強,組織開始損傷。 本試驗中GLUT1 和GLUT8 的比例關系的改變和對于炎癥因子基因表達的持續增加就說明了這一點。
IL-1β 又稱淋巴細胞活化因子, 也是由活化的單核巨噬細胞產生, 在組織炎癥狀態下發揮著重要功能。 Dinkova 等(2010)研究結果表明,不同濃度LPS 刺激奶牛乳腺上皮細胞24 h 后,細胞中IL-1β、IL-6 和IL-8 的mRNA 相對表達水平隨著LPS 刺激濃度的增加而增加,說明IL-1β、IL-6 在炎癥反應中會被激活以起到抗炎作用, 這也進一步驗證了本試驗結果及其相關推測。
4 結論
催乳素有利于改善氧化應激對乳腺乳蛋白基因表達的影響, 其作用是通過增加抗氧化基因的表達、 激活炎癥因子的表達和維持葡萄糖轉運因子的正常運轉而實現的。
猜你喜歡
中成藥(2021年5期)2021-07-21 08:39:04
世界科學技術-中醫藥現代化(2020年2期)2020-07-25 02:05:56
中成藥(2018年6期)2018-07-11 03:01:24
中成藥(2018年5期)2018-06-06 03:11:43
天然產物研究與開發(2016年6期)2016-06-05 10:29:26
西南軍醫(2016年6期)2016-01-23 02:21:19
新疆醫科大學學報(2015年10期)2015-12-26 12:33:30
吉林大學學報(醫學版)(2015年4期)2015-12-17 07:48:13
實用中西醫結合臨床(2015年7期)2015-02-28 16:30:23
癌變·畸變·突變(2015年3期)2015-02-27 06:15:09
