中國典型生態系統GPP的季節變異及其影響要素
2022-06-25 01:24:42姜鵬秦美歐李榮平孟瑩楊霏云溫日紅孫沛方緣
生態環境學報 2022年4期
關鍵詞:生長
姜鵬 ,秦美歐,李榮平,孟瑩,楊霏云,溫日紅,孫沛,方緣
1.中國氣象局沈陽大氣環境研究所,遼寧 沈陽 110016;2.中國氣象局氣象干部培訓學院遼寧分院,遼寧 沈陽 110016;3.沈陽區域氣候中心,遼寧 沈陽 110016;4.中國氣象局氣象干部培訓學院,北京 100081
總初級生產力(GPP)是單位時間內生物(主要是綠色植物)通過光合作用途徑所固定的有機碳量,決定著可供陸地生態系統生物利用的初始物質和能量(Wang et al.,2012)。作為大氣和陸地生態系統之間的最大碳通量,GPP受全球氣候變化的影響最為顯著,全球氣候變化通過影響生態系統自養呼吸和異養呼吸驅動生態系統呼吸(ER)的變異,而自養呼吸和異養呼吸共同決定了陸地生態系統的碳匯能力(Beer et al.,2010)。相對于ER,不同類型生態系統的研究均發現,GPP主導了生態系統的碳匯能力(Li et al.,2017b;Chen et al.,2020a),從而成為評估碳循環的重要指標(Beer et al.,2010)。中國陸地生態系統碳循環是全球碳收支的重要組成部分,對氣候變化非常敏感(He et al.,2019)。因此,深入探討中國陸地生態系統GPP的變化規律及其影響機制,將有助于提高對碳循環過程的理解,也將有助于精確評估及預測生態系統碳循環對氣候變化的響應及反饋。
基于點尺度的模擬氣候變化實驗-生態系統尺度的通量觀測-區域尺度的遙感監測的相關研究表明,輻射、溫度、水分等要素是GPP季節或年際變異的重要影響因子。模擬實驗主要關注GPP變異的內在影響機制,相關研究已揭示增溫等情境下的GPP對植被物候(Zhu et al.,2017)、群落結構(Li et al.,2020)以及葉片氮含量(Li et al.,2017a)等因子的響應過程及機制;通量和遙感觀測的研究主要探究環境因子與GPP之間的相關關系及相關變量的解釋權重,研究發現水分是草地生態系統GPP的主導因子(Zhang et al.,2018),而溫度和輻射與森林生態系統GPP年際變化的相關性較高(Yu et al.,2008)。目前已有證據表明,不同類型生態系統GPP對環境因子的響應過程和機制存在明顯的差異性,內蒙古溫帶草原和高寒草原GPP季節或年際變異的主導因子是水分,而高寒灌叢—草甸GPP變異的主導因子則是溫度(Zhang et al.,2016;柴曦等,2018;孟瑩等,2020)。中國東部季風區的森林生態系統研究發現,環境因子對GPP的驅動過程和機制在不同類型森林生態系統間存在著明顯的差異性(Yu et al.,2008),雖然基于全球FLUXNET數據的研究也發現了上述的異質性,但其研究的站點沒有覆蓋中國區域(尹茜茜等,2020)。然而,鮮有研究系統闡明中國草地、灌叢和森林生態系統GPP的變化規律、影響要素以及甄別生態系統間的差異性。
中國東部季風區沿水熱量梯度構成了一條完整的南北森林樣帶(North-South Transect of Eastern China,NSTEC),包含從溫帶落葉針葉林到亞熱帶常綠闊葉林等多種生態系統類型;沿著東西水熱梯度分布的草地樣帶(China Grassland Transect,CGT)覆蓋內蒙古溫帶草原、高寒灌叢-草甸和高寒草地等不同生態系統類型(于貴瑞等,2014),這為揭示大空間尺度下的 GPP變化規律及其影響因素提供了理想的天然實驗平臺。渦度相關法可在高時間分辨率下測量陸地生態系統和大氣之間的CO2、水和能量交換,基于此技術,可以獲取完整而可靠的生態系統碳水通量和氣象數據。中國陸地生態系統通量觀測研究網絡(ChinaFLUX)沿著NSTEC和CGT布設了多個通量站點,對生態系統碳交換已經開展了20余年的觀測(于貴瑞等,2014)。本研究以中國典型生態系統為研究對象,在NSTEC和CGT中選取3個森林站點、1個灌叢站點和2草地站點,在機器學習等多種統計分析的支撐下,揭示GPP的季節變化特征及其影響要素,量化影響要素的解釋權重,以及甄別生態系統類型間的差異性,為生態系統碳循環的模型模擬提供理論參考,為陸地生態系統對氣候變化的響應和反饋提供理論依據。
1 材料與方法
1.1 通量站點描述
沿著明顯的水熱組合梯度,NSTEC和CGT跨越了多個氣候帶和植被類型,兩條樣帶的直線距離均超過3000 km,包含多個具有完整獨特森林和草地景觀特征的連續植被帶。ChinaFLUX在NSTE和CGT兩條樣帶上沿著不同水熱梯度設立通量站點,利用渦度相關技術展開長期而連續的通量觀測。本研究沿著NSTEC和CGT選取6種具有區域代表性的生態系統為研究對象,分別為長白山溫帶針闊混交林、千煙洲亞熱帶常綠針葉人工林、鼎湖山亞熱帶常綠針闊混交林、內蒙古溫帶草原、海北高寒灌叢-草甸和當雄高寒草地。基于ChinaFLUX,在上述生態系統中選取相對應的通量站點,分別為長白山站、千煙洲站、鼎湖山站、內蒙古站、海北站和當雄站,各觀測站點地理位置等信息如表1所示,詳細的站點信息見參考文獻(Yu et al.,2008;Zhang et al.,2016)。
1.2 數據來源
1.2.1 通量及氣象數據來源
6個通量站數據來源于國家生態科學數據中心(http://www.cnern.org.cn/),目前這個數據平臺共享的通量數據時間范圍為2003—2010年。本研究主要關注生長季主要時期的 GPP變化特征及其影響要素。參考之前的相關研究,海北站和當雄站的研究時間范圍為6—9月(Chen et al.,2019);內蒙古站和長白山站為5—9月(Li et al.,2020);千煙洲站和鼎湖山站的生長季為全年,其中,將千煙洲站的7—10月定義為季節性干旱時期(Jiang et al.,2020),其余月份為正常時期,將鼎湖山站的4—9月定義為濕季,其余月份為旱季(Yu et al.,2008)。研究時間段內的GPP累積量超過其全年總量的80%。
常規氣象數據包含土壤含水量(CSW;5 cm深度)、土壤溫度(ts;5 cm深度)、空氣溫度(ta)、空氣相對濕度(Hr)、飽和水汽壓差(Dvp)、光合有效凈輻射(RPA)、太陽輻射(Rd)、凈輻射(Rn)和降水量(P),其中Dvp是指在一定溫度下,飽和水汽壓與空氣中的實際水汽壓之間的差值,DVP應用ta和Hr依據公式(1)求得。
通量和氣象數據的觀測儀器均由開路式渦度相關和常規氣象觀測系統組成,其中渦度相關系統包含紅外氣體分析儀(LI-7500,LI-COR Lincoln,USA)、三維超聲風速儀(CSAT3,Campbell Scientific,Logan,USA)和數據采集器(CR5000,Campbell Scientific,Logan,USA)。常規氣象觀測系統的日尺度數據采集時間段(Model CR23X,Campbell Scientific,Logan,USA)與通量數據相同。更為詳細的數據觀測介紹可以參考之前的研究(柴曦等,2018)。
1.2.2 葉面積指數數據獲取
本研究使用的葉面積指數(leaf area index,LAI)數據來源于美國國家航空航天局(NASA)開發的MODIS LAI產品,時間分辨率為4 d,空間分辨率為500 m。在此基礎上,獲取各個站點的經緯度坐標,采樣選擇以通量站點為中心,向南北和東西延伸各0.5 km的范圍,最終的LAI值為每個站點周圍3×3像素窗口內LAI的平均值(https://modis.ornl.gov/cgibin/MODIS/global/ subset.pl)。
1.3 統計方法
本研究使用4種統計分析方法量化 GPP季節變異的影響要素以及變量的相對貢獻,分別為Pearson相關分析、全子集回歸、隨機森林算法和方差分解分析。全子集回歸分析可以檢驗所有可能的模型,通過調整r2和Mallows CP統計量等準則選取“最佳”模型,在此基礎上,使用相對權重分析明確不同變量的解釋權重(Chen et al.,2020b)。在隨機森林算法中,mtry為節點值,可確定每次迭代的變量抽樣數值,用于決定二叉樹的變量數量,本研究選取最小殘差平方均值對應的mtry值作為最優值;ntree為隨機森林所包含的決策樹數目,本研究選取殘差平方均值穩定時對應的ntree值作為最優值,其中本研究中的mtry值為 2或者3,ntree值主要分布在400—600之間。方差分解通過對每種環境因子進行偏分析后,即可量化每種環境因子對GPP季節變異的貢獻。上述統計分析均在R 4.0.3中完成,其中全子集回歸、隨機森林算法以及方差分解分析分別是基于 leaps包中的 regsubsets()函數、randomForest包中的randomForest()函數和vegan包中的varpart()函數。
2 結果與分析
2.1 典型生態系統GPP及環境因子的季節變化特征
中國草地、灌叢和森林生態系統中環境因子和GPP的季節變化特征呈現出明顯的區域分異。6個通量站點的GPP季節變異整體呈單峰分布,千煙洲站GPP在6月達到生長季最大值,而后(7—8月)出現明顯的下降,而鼎湖山站的GPP在5—6月達到生長季峰值后保持至9—10月,其余站點的GPP在7—8月達到生長季最大值,森林站點的GPP生長季總量明顯高于灌叢和草地站點(圖1)。站點間Dvp的生長季變化特征差異較大,千煙洲站、鼎湖山站和內蒙古站的Dvp明顯高于其他站點,其中千煙洲站和內蒙古站的Dvp在 7—8月達到生長季最大值,海北站、長白山站和當雄站的Dvp在春季達到生長季最大值,而鼎湖山站的Dvp在6—7月和9—10月呈現出兩個峰值(圖1)。海北站、內蒙古站、千煙洲站和長白山站的CSW均在4—5月達到生長季最大值,其中千煙洲站的CSW在7—8月出現明顯的降低,當雄站的CSW在7—8月達到最大值,而鼎湖山站的CSW在4—10月保持相對穩定,長白山站、鼎湖山站和海北站的CSW高于其余站點(圖1)。各站點的ta季節變化特征較為一致,均呈單峰分布,在7—8月達到生長季最大值,其中鼎湖山站和千煙洲的ta高于其余站點。生長季RPA整體呈單峰分布,在6—8月達到生長季最大值,鼎湖山和千煙洲站的RPA在4—5月出現明顯的降低趨勢,草地和灌叢站點的RPA明顯高于森林站點(圖1)。除千煙洲站和鼎湖山站外,其余站點的P均在7—8月達到生長季最大值,千煙洲站的P在4—6月達到生長季峰值,而后明顯下降,呈現出“雨季—旱季”交替的變化趨勢,而鼎湖山站的P主要集中在4—9月,表現出明顯的旱季和雨季的分化(圖1)。
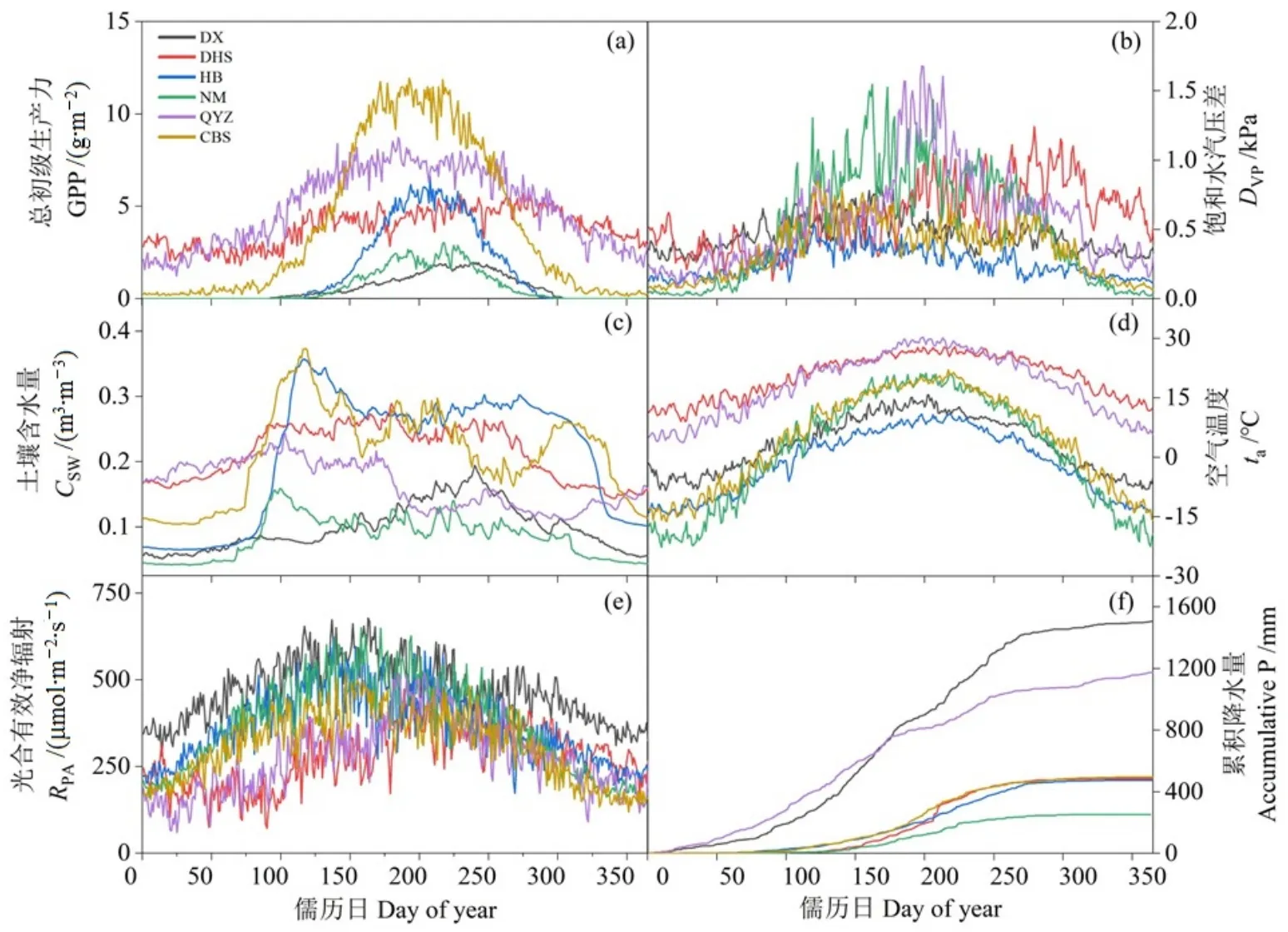
圖1 6個通量站點GPP、Dvp、CSW、ta、RPA和P的季節變化特征Figure 1 Seasonal dynamics of GPP, Dvp, CSW, ta, RPA and P
2.2 典型生態系統生長季GPP的影響要素分析
在中國草地、灌叢和森林生態系統中,生長季GPP的影響要素存在明顯的差異性。除鼎湖山站的GPP和CSW無顯著性相關關系外(P=0.069),其余站點GPP與其驅動要素在0.05、0.01或0.001的顯著性水平上均存在相關關系(圖2)。Pearson相關分析結果表明,草地生態系統站點的GPP與Dvp存在顯著的負相關關系,但與CSW和LAI表現出正相關關系,而與ta和RPA之間的相關系數小于上述3個因子(圖2)。與草地生態系統相比,灌叢和森林生態系統站點的ta、RPA和Dvp與GPP存在顯著的正相關關系,且相關系數高于其他因子與GPP之間的相關系數(圖2)。長白山站、鼎湖山站和海北站的CSW與GPP表現出負相關關系或相關系數不顯著,但千煙洲站的CSW和 GPP存在顯著的正相關關系(圖2)。
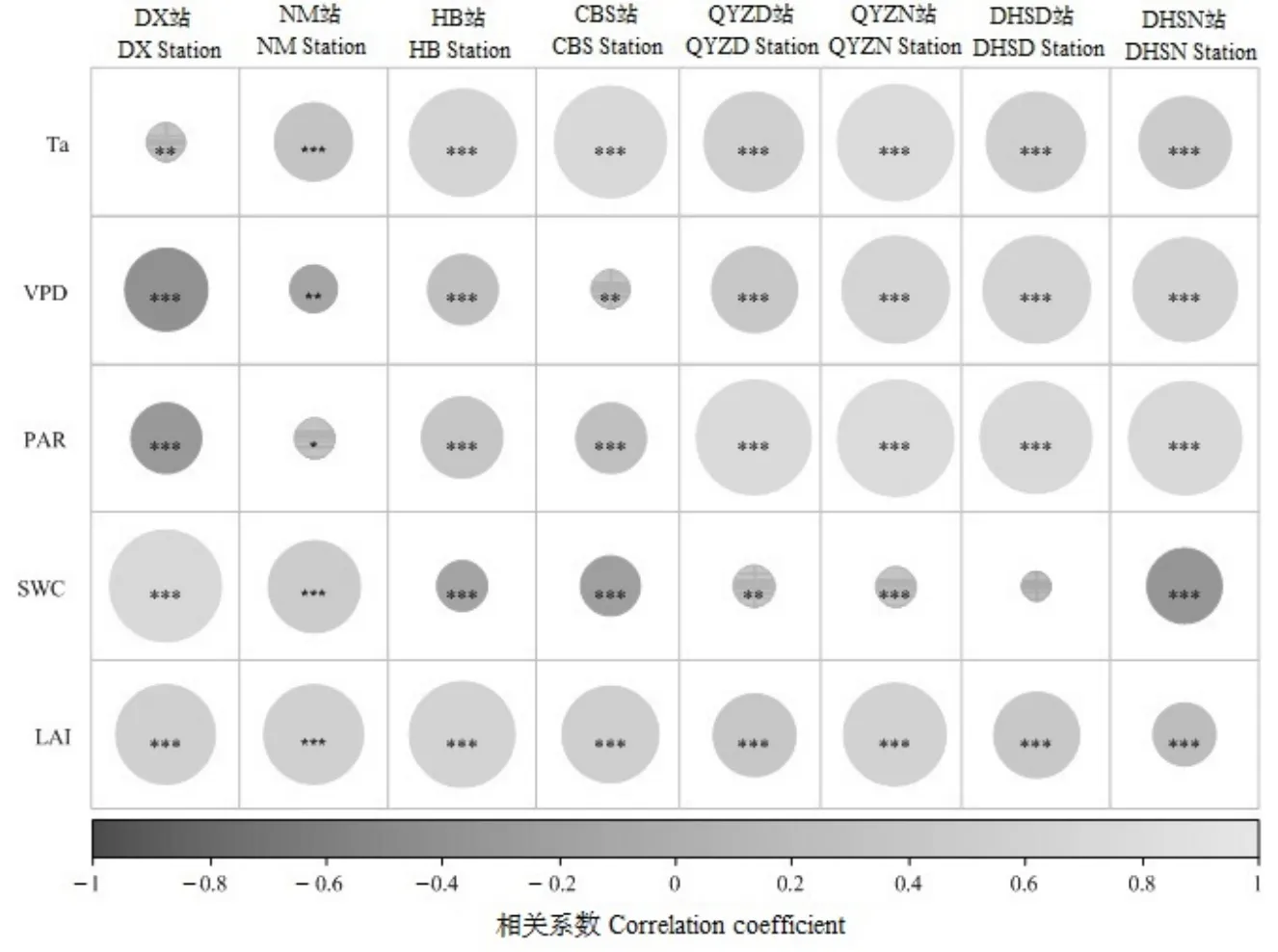
圖2 基于皮爾遜相關分析的生長季Dvp、CSW、ta、RPA和P與GPP之間的關系Figure 2 Correlations between Dvp, CSW, ta, RPA, LAI and GPP basing on Pearson correlation analysis
全子集回歸結果表明,草地生態系統站點GPP主要受CSW(P=0.000)和LAI(P=0.000)的正向影響,但GPP與Dvp(內蒙古站:P=0.000;當雄站:P=0.006)則表現出顯著的負相關關系(表2)。灌叢和森林站點中海北站和鼎湖山站的RPA和ta正向影響GPP的生長季變異(海北站:P=0.000和P=0.000;鼎湖山站:P=0.000和P=0.000);長白山站GPP受到CSW(P=0.004)和Dvp(P=0.000)的負向影響以及ta(P=0.000)、RPA(P=0.000)和 LAI(P=0.000)的正向影響;千煙洲站的ta、RPA和CSW與GPP表現出正相關關系(P=0.000),但旱季時的Dvp負向影響 GPP(P=0.000)(表2)。
2.3 典型生態系統生長季GPP變異的主要環境影響因子分析
相對權重分析結果表明,當雄站和內蒙古站的CSW主導 GPP的生長季變異,其中當雄站CSW對GPP變異的解釋比重高于60%;長白山站和海北站的ta能解釋超過或接近50%的GPP生長季變異;千煙洲站和鼎湖山站的RPA對生長季GPP變異的解釋權重超過50%(表2)。隨機森林算法結果發現,CSW是當雄站和內蒙古站 GPP變異的主要影響要素,解釋的比重超過40%;長白山站和海北站中的環境因子(ta和ts)主導GPP的季節變異,解釋權重均超過 20%;千煙洲站和鼎湖山站 GPP的主導因子均為RPA,其解釋的比重接近或者高于30%(圖3)。本研究將ta和ts作為溫度因子,CSW和Dvp作為水分因子以及RPA、Rd和Rn作為輻射因子,使用方差分解分析揭示不同因子對生長季 GPP變異的解釋權重。結果發現草地站點GPP生長季變異的主導因子為水分,而灌叢和森林站點GPP生長季變異的主導要素為溫度或輻射(圖4)。3種分析方法結果均表明,千煙洲站季節干旱時期的水分因子對GPP的解釋比重高于其他灌叢和森林站點(表2;圖3—4)。
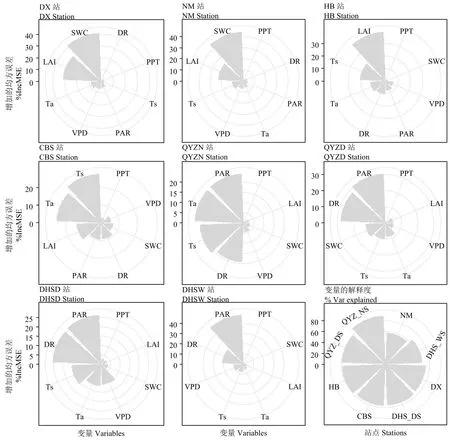
圖3 基于隨機森林算法的GPP生長季變異主導要素分析Figure 3 Driving factors of the variability in GPP based on random forest algorithm

圖4 基于方差分解算法的GPP生長季變異主導要素分析Figure 4 Driving factors of the variability in GPP based on variance partitioning analysis
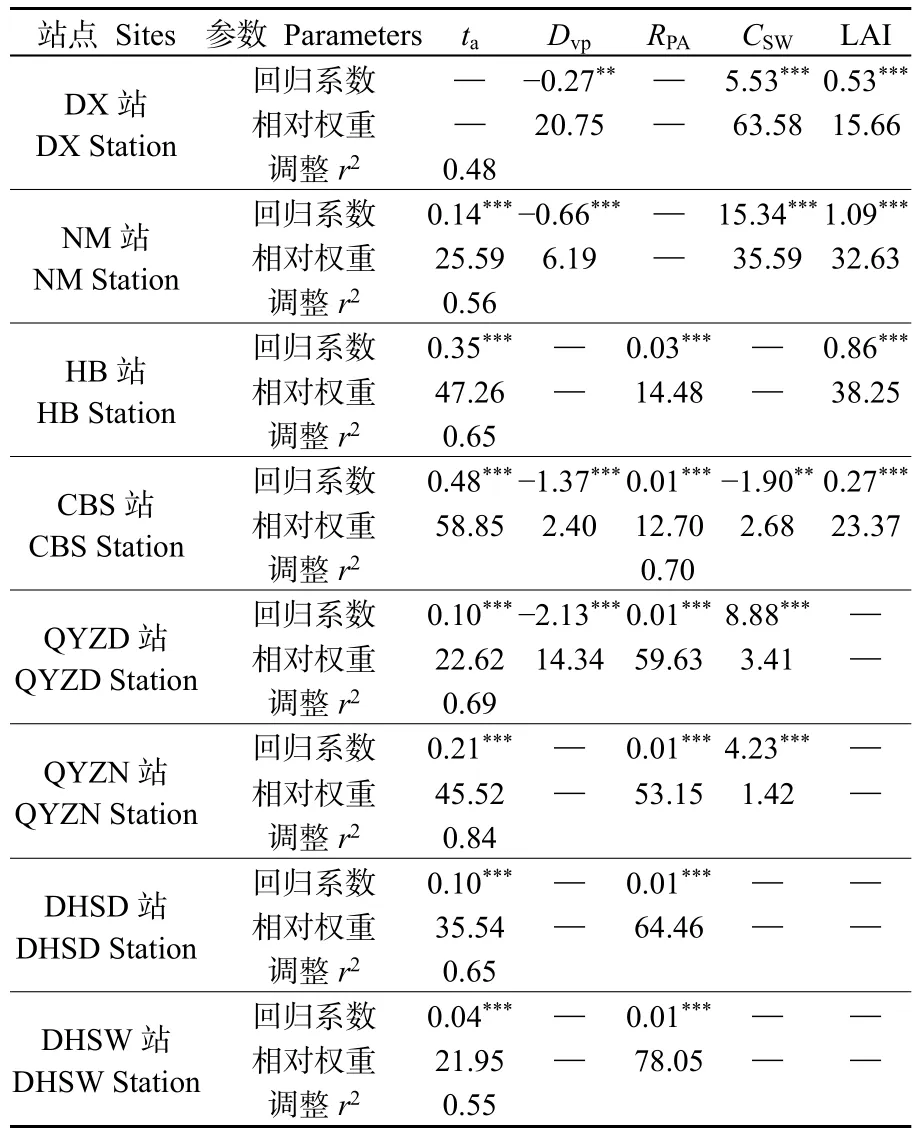
表2 基于全子集回歸分析生長季Dvp、CSW、ta、RPA和P與GPP之間的關系Table 2 Correlations between Dvp, CSW, ta, RPA, LAI and GPP based on full subset regression
3 討論
3.1 森林和灌叢生態系統GPP季節變異的影響因素
本研究與之前相關研究結果均表明,中國亞熱帶森林生態系統 GPP生長季變異的主導因子為輻射(Yu et al.,2008)。千煙洲站和鼎湖山站位于亞熱帶季風氣候區,生長季擁有充沛的降水和適宜的溫度,但RPA明顯低于其他生態系統的,尤其是在雨季時期,植被生長容易受到光照的限制。證據表明,亞熱帶森林生態系統雨季時期的凈碳吸收明顯低于旱季,主要是由于頻繁的大霧和陰雨天導致光照不充足(Yu et al.,2008)。區域內土壤類型主要為紅土,紅土具有粘性強、持水能力強等特點,結合充沛的降水量,這將削弱亞熱帶森林遭受水分限制的風險。即使在干旱時期,森林龐大的根系能夠吸取深層土壤水而維持葉面水勢,通過氣孔開放進行光合作用的收益將高于關閉氣孔保持的水分,表現出“開源”而不是“節流”的水分利用策略(Konings et al.,2017;Jiang et al.,2020;Deng et al.,2021)。但亞熱帶森林的水分利用策略也與干旱事件的強度相關,例如2009、2010年中國西南部的極端干旱事件明顯降低森林的固碳能力(Li et al.,2019)。
中國溫帶森林和高寒灌叢草甸生態系統 GPP季節變異的主要影響要素為溫度。長白山站和海北站位于中國中高緯和高海拔地區,冬季寒冷而漫長,生長季降水量在500—700 mm范圍內,生長季輻射較為充足,溫度明顯低于亞熱帶地區。區域內土壤有機質和黏土含量較高,加之低溫誘導的低蒸發量,使得土壤持水能力較強(Li et al.,2016;Yu et al.,2008)。此外,兩個站點位于中國季節性積雪的主要分布區,春季積雪融水能進一步補充土壤水分,致使其生長季CSW接近甚至超過亞熱帶地區(Yu et al.,2008)。研究甚至發現,海北站的CSW和P無顯著性相關關系,進一步證明了該區域土壤具有含水量高和持水能力強等特征(Li et al.,2016)。上述自然環境特征導致溫度成為GPP季節變異的主導因素,其驅動機制如下:(1)溫度通過影響春季植被返青期、生長季長度而驅動 GPP的季節變異(Zu et al.,2018);(2)溫度能夠通過促進葉綠體膜的穩定性,提升核酮糖-1,5-二磷酸羧化酶/加氧酶(Ribulose-1,5-bisphosphate carboxylase/Oxygenase,通常簡寫為RuBisCO)的速率和活性,提高電子傳遞能力、熱穩定性等途徑直接影響 GPP(Booth et al.,2012);(3)溫度通過調控土壤微生物活性以及土壤有機質分解,進而影響土壤氮礦化速率和菌根氮吸收速率,這些過程將影響葉片氮濃度以及CO2的固定速率進而影響GPP(Wohlfahrt et al.,2008)。
總體上,本研究發現溫度和輻射主導了溫帶和亞熱帶森林生態系統GPP的季節變異,但在季節干旱期間,水分因子對千煙洲站GPP存在著不可忽略的影響。與其他站點雨熱同期相比,千煙洲站所在區域每年7—10月受副熱帶高氣壓帶控制,出現“雨熱不同期”的現象,該段時間P和CSW出現明顯的降低趨勢,但溫度卻達到了生長季峰值(Xu et al.,2017),進而導致該區域中水分因子對GPP變異的貢獻要高于其他森林或灌叢生態系統中的。除CSW外,“雨熱不同期”的高溫少雨天氣明顯增加了Dvp,而過高的Dvp通過限制植被氣孔開放程度抑制植被光合能力,且之前的研究發現,干旱時期GPP對Dvp的響應敏感性要強于其他情境(Sulman et al.,2016)。因此,即使亞熱帶森林生態系統擁有較高的土壤含水量和持水能力以及龐大的根系系統,但在季節性干旱期間,GPP仍可能受到水分的限制,尤其是受到過高Dvp的抑制。基于目前我們在投稿的文章,在季節性干旱期間,當Dvp高于0.8 kPa時,Dvp與GPP呈現出顯著的負相關關系。
3.2 草地生態系統GPP季節變異的影響因素
在中國溫帶草原和高寒草地生態系統中,相關研究一致認為水分是植被生長和碳通量生長季變異的主要驅動要素(李夏子等,2013;耿曉東等,2017;Zhang et al.,2018)。傳統觀點認為青藏高原地區溫度是植被生長的限制因子,但高原中西部高寒草地區域的通量以及降水-溫度耦合模擬實驗結果均發現,水分為GPP的主導因子,而不是溫度或者輻射(Fu et al.,2018;Zhang et al.,2018)。中國草地生態系統主要分布在半干旱地區,生長季降水量在400—500 mm之間,區域內輻射及風力較強,加之土壤中砂石含量較高,植被覆蓋率低,導致土壤持水能力較差(Chen et al.,2009;Peng et al.,2013)。草地生態系統的優勢種多為淺根物種,與高大植被相比,矮小植被受到的水分限制更大(Matheny et al.,2014),這是由于淺根物種的生長主要依靠淺層土壤水分,但自然環境特征導致草地生態系統淺層土壤的持水能力較差,進一步增強水分因子的效應。水分因子對草地生態系統GPP生長季變異的驅動機制主要表現在以下3個方面:(1)CSW通過驅動植被返青期和生長季長度而影響生長季GPP,尤其是群落中淺根物種的物候期對水分變化更為敏感(Zhu et al.,2017);(2)水分變化通過改變群落結構而影響GPP的生長季動態(Liu et al.,2016)。研究發現,降水充沛條件下,CSW通過增加群落中淺根優勢種的蓋度而增加生長季GPP(Chen et al.,2019);(3)植物木質部或根系通過根部脫落酸濃度感知土壤水分狀況,進而驅動葉片氣孔狀態,實現對光合作用和蒸騰作用的調控(Martínez-Vilalta et al.,2014)。
目前多數研究只關注水分因子中的CSW對GPP的影響,而Kimm et al.(2020)研究發現,Dvp同樣對GPP的季節變異產生了重要的影響,其效應甚至強于CSW。氣孔既是植物光合作用吸收CO2的入口,又是蒸騰作用水分散失的出口,在平衡優化植物光合與蒸騰關系中發揮著重要作用。而Dvp是植被蒸騰作用的重要調控因子,當Dvp過高時,植物傾向于關閉氣孔,以防止水分流失,進而削弱植被光合能力(Monteith,1995)。本研究發現,草地生態系統生長季Dvp與GPP存在顯著的負相關關系,這表明過高的Dvp將抑制GPP。雖然草地生態系統中的Dvp小于森林生態系統,但其較低的土壤含水量和持水能力以及優勢種多為淺根系植物,共同決定了較低的土壤水勢和葉片水勢,削弱葉片氣孔對高Dvp的抵抗能力,進而導致草地生態系統GPP容易受到Dvp的限制(Konings et al.,2017;Ding et al.,2018)。這與孟瑩等(2020)研究基本一致,即相對于森林等生態系統,草地生態系統GPP受Dvp的抑制作用更強。
4 結論
本研究基于中國草地、灌叢和森林生態系統中的6個ChinaFLUX通量站點數據,結合隨機森林等統計分析方法,系統闡明了中國典型生態系統GPP的季節變化特征、影響因素以及生態系統類型間的差異性。本研究結果表明,6個通量站點生長季GPP總體呈現單峰分布,且森林生態系統生長季GPP高于灌叢和草地生態系統。除千煙洲站外,其余站點的環境因子在生長季呈現出雨熱同期的現象,而千煙洲站受副熱帶高氣壓帶控制,在 7—10月經歷季節性干旱,出現“雨熱不同期”的現象。草地站點GPP的季節變異與CSW、LAI呈正相關,與Dvp呈負相關,與溫度和輻射的相關性較弱,而森林和灌叢站點的 GPP主要受ta和RPA的正向影響,與水分因子的關系較弱。整體上,水分因子主導草地生態系統GPP的季節變異,而溫度或輻射因子主導灌叢和森林系統GPP的季節變異,但在季節性干旱期間,水分因子對千煙洲站GPP的季節變異的解釋比重高于對其他灌叢和森林站點的。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
