NO 誘導IAA 和O2·-積累于側根尖端促進水稻側根生長
2022-06-29 09:31:54朱雄梅蔡慧楠劉瑞琪林佩錦郞雙怡黃悅鄧舒琪莫億偉金晨鐘
農業現代化研究 2022年3期
關鍵詞:水稻
朱雄梅,蔡慧楠,劉瑞琪,林佩錦,郞雙怡,黃悅, 鄧舒琪,莫億偉*,金晨鐘*
(1. 湖南人文科技學院農田雜草防控技術與應用協同創新中心,湖南 婁底 417000;2. 紹興文理學院生命科學學院,浙江 紹興 312000)
植物根系主要由兩部分組成,一種起源于胚根的初生根,另一種在主根上形成側根及生長于成熟組織的不定根。側根和根毛的形成均受到多種植物激素調控[1-2],特別是生長素(IAA)在側根的起始分化和形成中起著關鍵作用[3],而根系的IAA 源于地上部分向根部運輸,當IAA 向根部運輸增多時則利于更多側根形成[4];當IAA 極性運輸受阻,側根的數量則顯著下降[5]。一氧化氮(Nitric oxide,NO)在參與植物逆境調節和側根發育中均起著重要作用,植物體內的NO 合成有兩種途徑,一種是依賴亞硝酸還原途徑依靠硝酸還原酶1(NIA1) 和硝酸還原酶2(NIA2),另一種是依靠L-精氨酸的氧化途 徑[6-7]。有研究表明,提供外源NO 供體硝普鈉(Sodium nitroprusside,SNP)時,可使番茄側根原基及側根數量顯著增多[8];當受NO 合成酶(NOS)抑制劑(N’-硝基-L-精氨酸甲酯鹽酸鹽,L-NAME)處理時,根系NO 含量顯著降低并抑制側根形成[9]。基于IAA 和NO 在側根形成過程中均起到重要的調控作用,但這兩種信號物質相互調節機制尚未弄清,有的研究認為NO 是IAA 的上游信號物質,因為在擬南芥葉片發育中發現,NO 可調控IAA 極性運輸輸出載體PIN1 蛋白積累于葉原基處,從而促進葉片形成[10-11];有的研究則認為NO 是IAA 的下游信號物質,因為用NOS 抑制劑L-NAME 或NO 清除劑(2-(4-羧基苯基)-4,4,5,5-四甲基咪唑啉-1-氧基-3-氧化物鉀鹽,cPTIO)處理后可抑制IBA 對側根的誘導,且NAA 可通過誘導NO 的合成促進番茄側根發育[12]。此外,活性氧(Reactive oxygen species, ROS)作為一種信號分子,在植物生長發育和激素信號調控中起著重要作用,如ROS 增多也可促進側根及根毛形成[13-14]。由此說明,IAA、NO和O2·-在調控側根形成過程中均有著重要作用,但前人的相關研究結果主要來源于雙子葉模式植物擬南芥等,在水稻等主要農作物則研究較少。前人為了更直觀地探討IAA 定位和功能的關系,有研究發現,DR5 比天然的IAA 反應元件(AuxREs)具有更強的IAA 反應能力,表達活性可增加5~10 倍[15], 以DR5 為啟動子與報告基因重組,構建成IAA 響應報告基因DR5::GUS,轉入植物體獲得轉基因植株,已成為研究IAA 的定位及其作用的良好工具[16],利用DR5::GUS 報告基因研究IAA 的分布已在擬南芥[11]、水稻[17]和棉花[18-19]等多種植物得到應用。水稻是重要的糧食作物,側根形成的水平直接影響水稻植株對養分的吸收能力,本研究以DR5-GUS標記轉基因水稻為材料,深入探討IAA、NO 和O2·-對水稻側根形成的影響,以期提高水稻根系的養分利用效率。
1 材料與方法
1.1 實驗材料和處理
DR5::GUS 標記的轉基因水稻(Oryza sativa L.)由華中農業大學王學路教授饋贈,SNP、NO 合成酶抑制劑(L-NAME)、2-(4-羧基苯基)-4,4,5,5-四甲基咪唑啉-1-氧基-3-氧化物鉀鹽(cPTIO)、NO 熒 光 探 針(5,6-Diaminofluorescein diacetat,DAF-2DA)、IAA 檢測的DR5::GUS 報告基因染色液均購自生工生物工程(上海)股份有限公司。水稻種子浸種和催芽后,選擇種子根長約為0.5 cm 生長一致的幼苗,置于含兩層濾紙的培養皿中,加入1/50 MS 微量元素培養溶液(以防單鹽毒害),保持根系置于培養液中進行相應的處理。
1.2 NO 供體及清除劑對水稻種子根根系生長的影響
為研究NO 供體及NO 清除劑對水稻種子根根系生長的影響,設計了如下三組試驗。1)為了探討外源NO 供體對側根形成的影響,設計在1/50 MS 微量元素培養溶液中分別加入0(對照)、5、10、15、20 和25 μmol/L SNP 的處理。2)為了探討NO 合成酶抑制劑對側根形成的影響,設計在1/50 MS 微量元素培養溶液中分別加0(對照)、50、100、200、400、800 μmol/L NO 合成酶抑制劑的處理。3)為了探討NO 清除劑對側根形成的影響,設計在1/50 MS 微量元素培養溶液中分別加0(對照)、50、100、200、400、800 μmol/L cPTIO 的處理。
所有處理組均置于光照培養箱中,每個處理設置4 個生物學重復,每個培養皿放置8 粒種子。光照培養箱的溫度設置為28 ℃,光照強度為 500 μmol/(m2·s),每天13 h 光照,11 h 黑暗,相對濕度為85%。每天補充相應溶液,在處理的第3 d和第5 d,分別測量初生根、側根、冠根的長度及數量。
1.3 NO、IAA 和O2·-在側根起始部位的定位觀察
1.3.1 NO 染色 經上述處理后,第3 d 剪取開始出現側根生長的初生根,參考Sánchez-Vicente 等[10]的方法,用100 mmol/L pH7.2 的磷酸鈉緩沖液配制成10 μM DAF-2DA 溶液,在25 ℃黑暗條件下對根系染色30 min,再用ddH2O 浸泡3 次,每次5 min,以洗掉材料表面的染料,置于顯微鏡下觀察并照相。
1.3.2 GUS 染色 參考胥華偉[20]等用GUS 染色法確定IAA 在側根的定位,具體方法為根系材料用GUS 染色液(100 mmol/L pH7.2 磷酸鈉緩沖液、0.5 mmol/L 高鐵氰化鉀、0.5 mmol/L 亞鐵氰化鉀、1.0 mmol/L X-gluc 和2.0% DMSO)在37 ℃染色2 h 后,經ddH2O 浸泡3 次,每次5 min,以洗掉材料表面的染料,置于顯微鏡下觀察并照相。
1.3.3 NBT 染色 將根系材料置于NBT 染液中(100 mmol/L pH7.2 磷酸鈉緩沖液,NBT 濃度為0.1mg/mL), 室溫染色1.5 h,倒掉染色液,再用ddH2O 浸泡3次,每次5 min,以洗掉材料表面的染料后,置于顯微鏡下觀察并照相。根據染色的程度判斷各處理對NO、IAA 和O2·-在側根起始部位分布的影響。
1.4 SNP 和NO 合成酶抑制劑處理對NO 形成相關基因表達水平的檢測
取上述不同SNP 及NO 合成酶抑制劑處理3 d的幼苗初生根(基部1.5 cm 內能形成側根的部分),通過Takara RNA 提取試劑盒提取RNA,反轉錄成cDNA,-20 ℃保存備用。參考Zhou 等[21]方法,利用實時熒光定量PCR 分析與NO 合成相關的硝酸還原酶1(OsNIA1)和硝酸還原酶2(OsNIA1)表達情況,以水稻Actin1 作為內參基因OsActin1 (LOC_Os03g50890),具體引物設計如下:OsNIA1 引物為:F:5′-CCAATTCTTTCATCGTGTTCT-3′,R:5′-CATG CAGCATTTCGTTTCT-3′;OsNIA2 引物為:F:5′-AC TGGTGCTGGTGCTTCTGG-3′,R:5′-CGGCTGGG TGTTGAGGGACT-3′;Actin1 引物為:F:5′-CAACA CCCCTGCTATGTACG-3′,R:5′-CATCACCAGAGTC CAACACAA-3′。熒光定量PCR 試劑盒為Thermo Scientific DyNAmo ColorFlash SYBR Green qPCR Kit (Thermo),儀器為Stratagene Mx3005P 熒光定量PCR 儀,反應體系及條件參照試劑盒說明書進行。以上實驗至少重復5 次,結果用平均值±SD 表示,用SPSS16.0 軟件進行單因素方差分析。
2 結果與分析
2.1 不同濃度SNP 處理對水稻初生根長度及側根數量的影響
從表1 可知,SNP 處理濃度為5 μmol/L 時,在處理后的第3 d 和第5 d,初生根長、側根和冠根的數量與對照相比無顯著差異(P >0.05);當SNP處理濃度達10 μmol/L 以上時,隨著SNP 處理濃度的增加,初生根長度、側根數量和冠根數量也逐漸增大,各處理初生根長度、側根數量及冠根數量均顯著大于對照處理。其中以SNP 處理為20 μmol/L時效果最佳,在處理后的第3 d,初生根長度、側根數量和冠根數量分別比對照處理提高59.95%、26.89%和67.80%,均達顯著性差異(P <0.05);在處理后的第5 d, 初生根長度、側根數量和冠根數量分別比對照處理提高28.04%、25.37%和65.49%,均達顯著性差異(P <0.05)。但當SNP 濃度達25 μmol/L 時,與20 μmol/L 的使用濃度相比,對初生根長度、側根數量及冠根數量不再有顯著促進作用(P >0.05),結果表明水稻幼苗根系生長需要NO信號的參與。

表1 硝普納處理對初生根長度、側根及冠根數的影響Table 1 The effects of SNP treatment on primary root length, lateral root and crown root number
2.2 NO 合成酶抑制劑對水稻初生根長度及側根數量的影響
從表2 可知,當NO 合成酶抑制劑為50 μmol/L 時,在處理后的第3 d 和第5 d,初生根長、側根和冠根的數量與對照相比無顯著差異(P >0.05);當NO 合成酶抑制劑達100 μmol/L 以上時,隨著NO合成酶抑制劑處理濃度的增加,初生根長度、側根數量和冠根數量也逐漸減小。NO 合成酶抑制劑濃度達100 μmol/L 以上時,各處理的初生根長度、側根數量及冠根數量均顯著低于對照處理。其中以NO 合成酶抑制劑為800 μmol/L 時效果最佳,如在處理后的第3 d,初生根長度、側根數量和冠根數量分別比對照處理降低30.96%、56.94%和55.05%,均達顯著性差異(P <0.05);在處理后的第5 d 時,初生根長度、側根數量和冠根數量分別比對照處理降低51.38%、64.12%和58.59%,也均達顯著性差異(P <0.05)。結果表明NO 合成受到抑制時,也抑制了根系的生長。
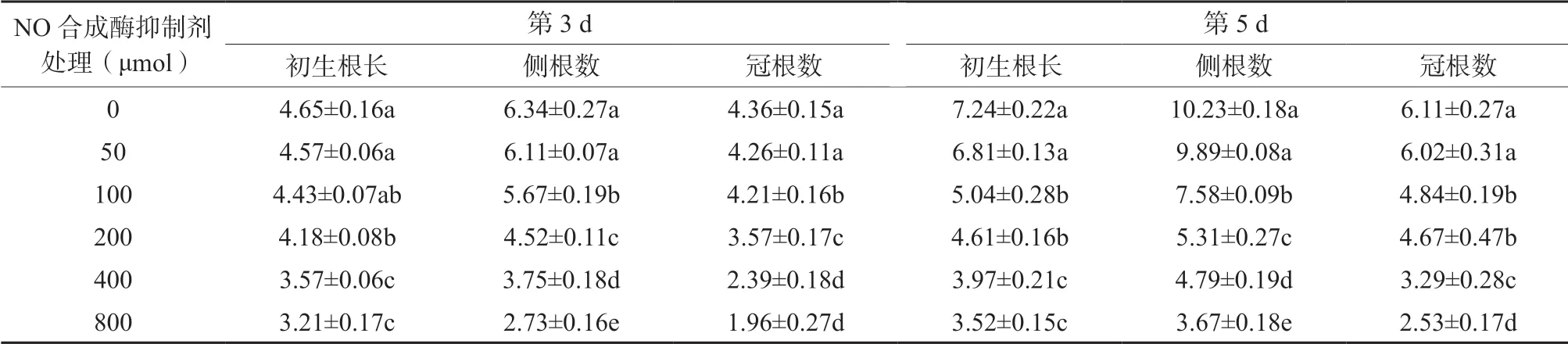
表2 NO 合成酶抑制劑處理對初生根長度、側根及冠根數的影響Table 2 The effects of NO synthetase inhibitor treatment on primary root length, lateral root and crown root number
2.3 NO 清除劑處理對水稻初生根長度及側根數量的影響
從表3 可知,當NO 清除劑為50 μmol/L 時,在處理的第3 d 和第5 d,初生根長、側根和冠根的數量與對照相比無顯著差異(P >0.05);當NO清除劑達100 μmol/L 以上時,隨著NO 清除劑處理濃度的增加,初生根長度、側根數量和冠根數量也逐漸減小。當NO 清除劑cPTIO 濃度達100 μmol/L 以上時,各處理的初生根長度、側根數及冠根數均顯著低于對照處理,其中以800 μmol/L NO 清除劑cPTIO 處理的效果最佳,如在處理后的第3 d,初生根長度、側根數量和冠根數量分別比對照處理降低49.80%、56.26%和76.75%,均達顯著性差異 (P <0.05);在處理后的第5 d 時,初生根長度、側根數量和冠根數量分別比對照處理降低56.67%、56.61%和55.41%,均達顯著性差異(P <0.05)。結果表明,當根系的NO 受到cPTIO 清除后,初生根、側根及冠根的生長均受到抑制作用。

表3 NO 清除劑處理對初生根長度、側根及冠根數的影響Table 3 The effects of NO scavenger treatment on primary root length, lateral root and crown root number
2.4 SNP、NO 合成酶抑制劑及清除劑處理對側根NO 分布的影響
從圖1 可知,經SNP 處理后,結果發現NO 在根表皮及根內大部分細胞均有分布,主要集中分布于側根形成部位和側根根尖部分,且熒光強度越大,表示NO 含量越高。與對照相比,外源SNP 處理后,NO 熒光強度明顯大于對照處理。受NO 合成酶抑制劑和清除劑處理后,染色程度明顯下降。側根清除處理后起始部位的NO 含量明顯減少,且NO 的熒光強度最淺,側根數量少而短(圖1),結果表明當NO 積累減少,側根數量下降。
2.5 SNP、NO 合成酶抑制劑及清除劑處理對側根IAA 分布的影響
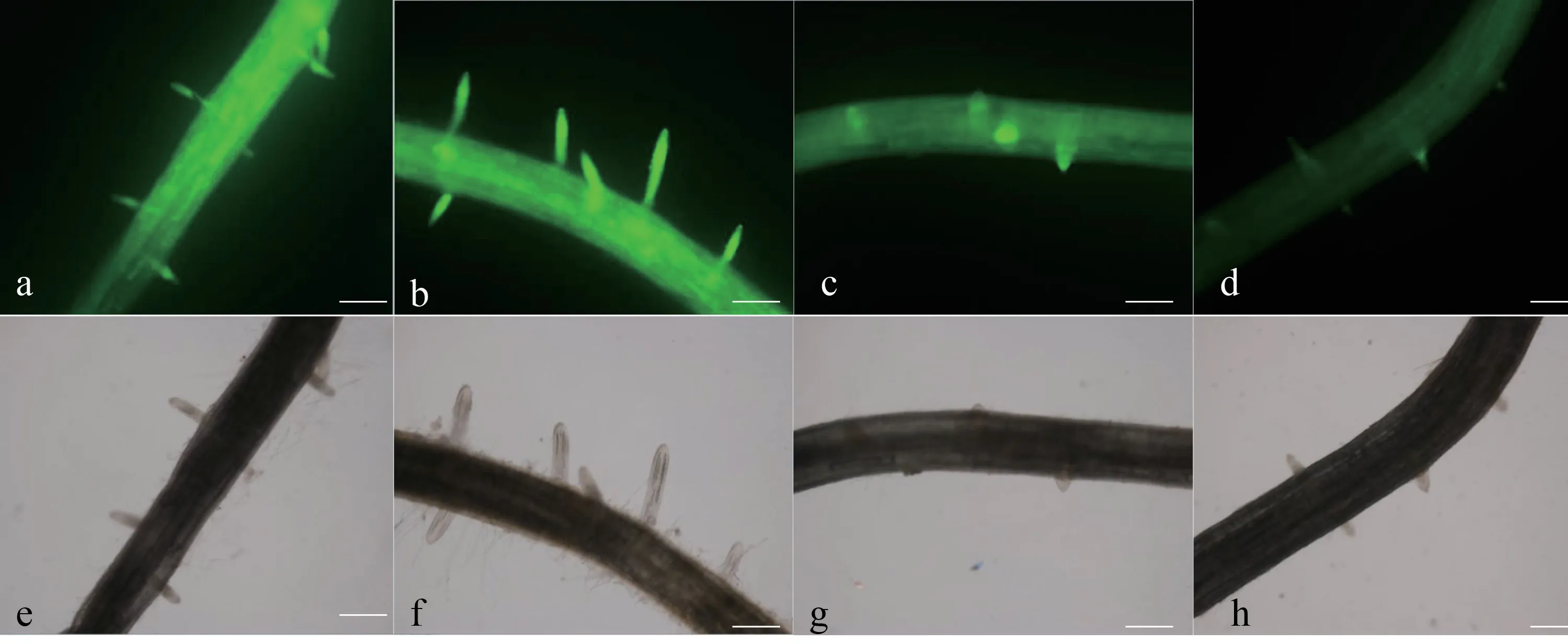
圖 1 SNP、NO 合成酶抑制劑及清除劑處理對NO 分布的影響Fig. 1 The effects of SNP, NO synthetase inhibitor and scavenger treatments on NO distribution
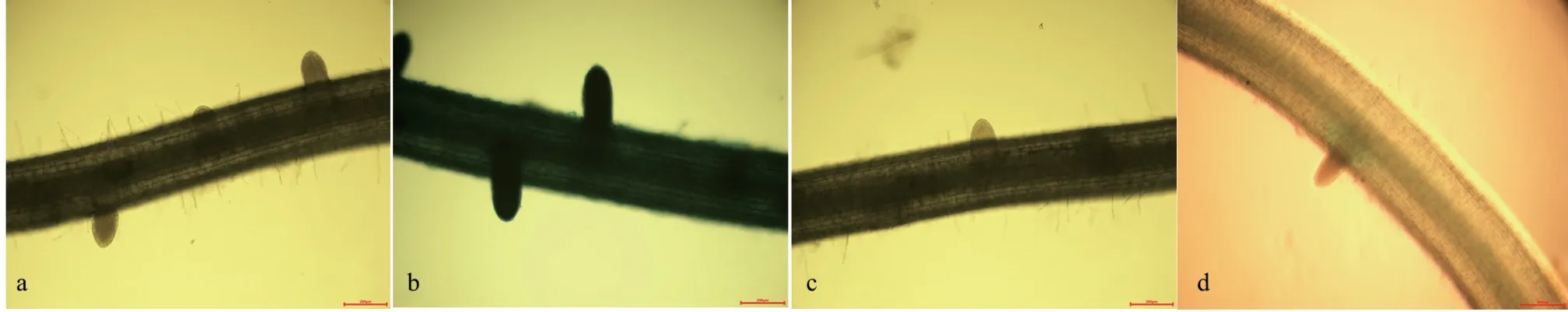
圖 2 SNP、NO 合成酶抑制劑及清除劑處理對IAA 分布的影響Fig. 2 The effects of SNP, NO synthetase inhibitor and scavenger treatments on IAA distribution
從圖2 可知,經SNP 處理后,通過對DR5:: GUS 標記水稻側根起始部位染色,結果發現IAA主要分布在根的維管束部位,極性分布于側根形成部位和側根根尖部分,在初生根的維管束與側根形成起始部位,明顯看到染色較深,說明IAA 含量較多(圖2),與對照相比,外源的SNP 處理后,染色程度明顯大于對照處理。而受到NO 合成酶抑制劑和清除劑處理后,側根數量減少,染色程度明顯下降,以受NO 清除劑處理后,IAA 的GUS染色最淺,結果表明由初生根向側根運輸IAA 含量明顯減少,IAA 極性積累于側根起始形成部位,有益于側根形成,當IAA 積累減少,側根形成量顯著下降。
2.6 SNP、NO 合成酶抑制劑及清除劑處理對側根O2·-分布的影響
經SNP 處理后,用NBT 對水稻側根的起始部位染色,結果發現O2·-主要分布在根的維管束部位,極性分布于側根形成部位及側根根尖部分,在初生根維管束與側根形成起始部位,明顯看到染色較深,表明O2·-含量較多(圖3)。與對照相比,外源SNP處理后,染色程度明顯大于對照處理。受到NO 合成酶抑制劑和清除劑處理后,O2·-染色程度明顯下降。表明O2·-極性積累于側根起始形成部位,有益于側根形成,當O2·-積累減少時,側根的形成量顯著下降。
2.7 外源SNP 對OsNIA1 和OsNIA2 基因表達的影響
從圖4 可知,隨著SNP 處理濃度的增加,對基因OsNIA1 和OsNIA2 相對表達量的促進作用逐漸增大。當SNP 達10 μmol/L 以上時,各處理濃度均可顯著提高OsNIA1 和OsNIA2 基因的表達(P <0.05)。如當SNP 濃度為15 μmol/L、20 μmol/L和25 μmol/L 時,OsNIA1 表達量分別比對照處理增 加 了107.65%、88.82% 和79.41%(P <0.05),OsNIA2 表達量分別比對照處理增加了109.36%、152.71%和146.31%(P <0.05)。表明外源SNP 能促進與NO 合成相關基因OsNIA1 和OsNIA2 的表達,且當SNP 濃度達到15 μmol/L 時,對OsNIA1誘導效果最好,而SNP 濃度超過20 μmol/L 時,對OsNIA2 誘導效果最佳。
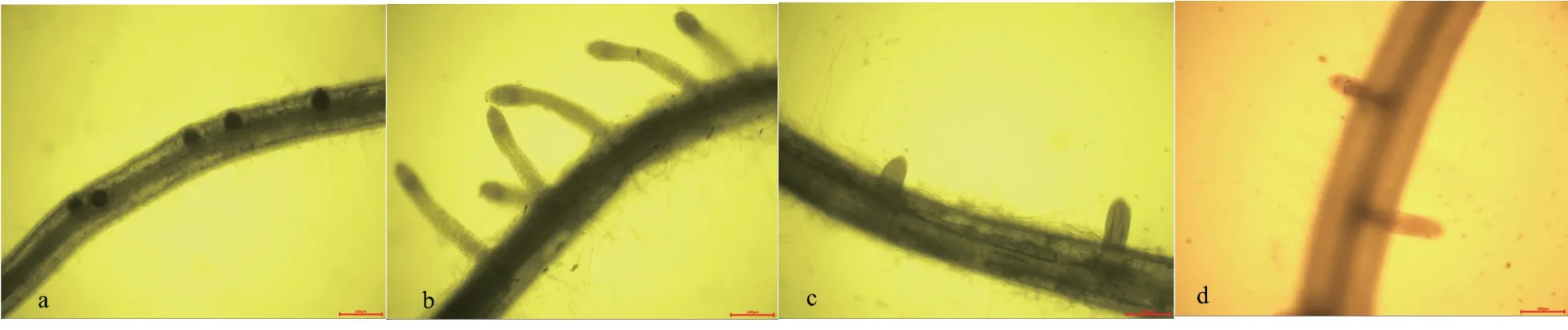
圖 3 SNP、NO 合成酶抑制劑及清除劑處理對側根O2·-分布的影響Fig. 3 The effects of SNP, NO synthetase inhibitor and scavenger treatments on O2·- distribution
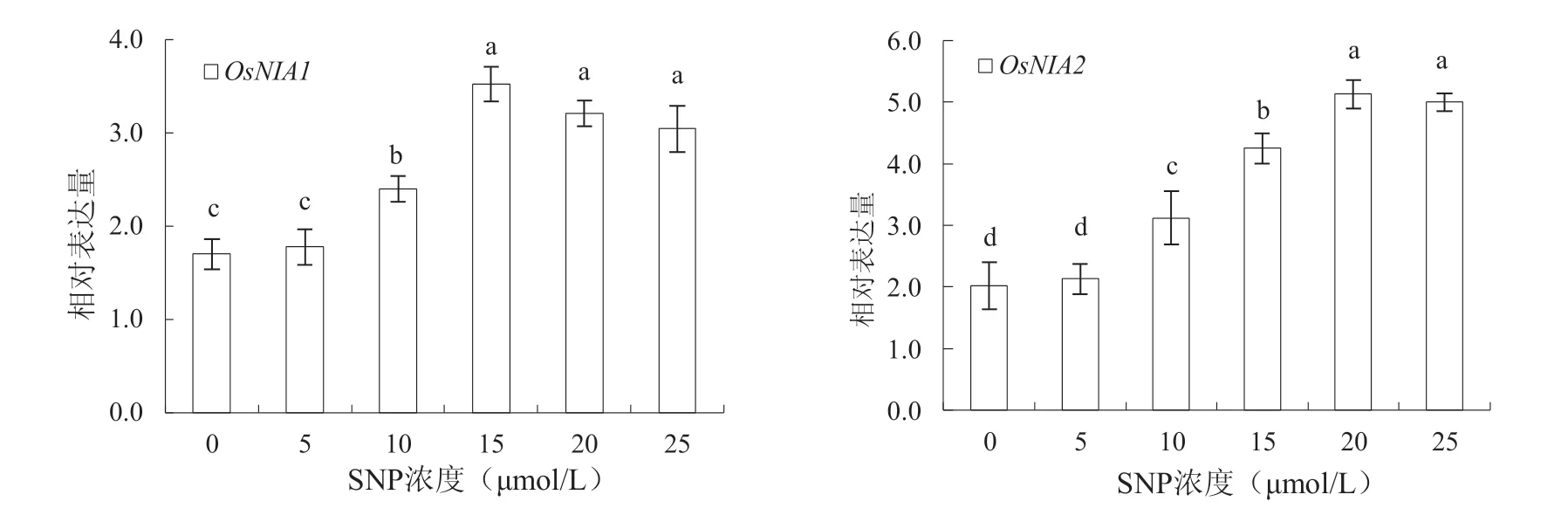
圖4 SNP 處理對OsNIA1 和OsNIA2 基因表達的影響Fig. 4 The effect of SNP treatment on the expression of OsNIA1 and OsNIA2
2.8 外源NO 合成酶抑制劑對OsNIA1 和OsNIA2基因表達的影響
從圖5 可知,隨著NO 合成酶抑制劑處理濃度的增加,對基因OsNIA1 和OsNIA2 相對表達量的抑制作用逐漸增大。當NO 合成酶抑制劑達100 μmol/L 時,可顯著抑制OsNIA1 表達(P<0.05),對OsNIA2 表達沒有顯著影響(P>0.05),當NO 合成酶抑制劑達200 μmol/L 時,可顯著抑制OsNIA2 的表達(P<0.05),當NO 合成酶抑制劑濃度為800 μmol/L 時,對OsNIA1 和OsNIA2 的表達量受到的抑制作用最大,分別比對照下降63.85%和60.58%(P<0.05)。表明外源NO 合成酶抑制劑可抑制與NO 合成相關基因OsNIA1 和OsNIA2 的表達。
2.9 外源SNP 對NO 含量及O2·-產生速率的影響
從圖6 可知,當SNP 處理濃度低于5 μmol/L時,對NO 的含量無促進作用,與對照相比無顯著性差異(P >0.05)。當SNP 達10 μmol/L 以上時,隨著SNP 處理濃度的增加,對NO 含量的促進作用也逐漸增大,如SNP 濃度為10 μmol/L、15 μmol/L、20 μmol/L 和25 μmol/L 時,NO 含量則分別比對照增加了45.99%、78.17%、96.43%和71.42%,均達顯著性差異(P<0.05),其中當SNP 濃度為20 μmol/L處理時,NO 含量最高,當濃度達25 μmol/L 時,NO 含量又有所下降。進一步探究SNP 處理對側根形成部位O2·-產生速率的影響時發現,當SNP 處理濃度低于5 μmol/L 時,對O2·-的產生速率無促進作用,與對照相比無顯著性差異(P >0.05)。當SNP 達10 μmol/L 以上時,隨著SNP 處理濃度增加,對NO 含量和O2·-產生速率的促進作用也逐漸增大,如在SNP 濃度為10 μmol/L、15 μmol/L、20 μmol/L和25 μmol/L 時,O2·-產生速率分別比對照增加了64.07%、131.13%、208.98%和174.65%,均達顯著性差異(P<0.05),當SNP 濃度為20 μmol/L 處理時,O2·-產生速率最大,當濃度達25 μmol/L 時,O2·-產生速率又有所下降。
3 討論
3.1 水稻側根形成需要NO、IAA 和O2·-極性積累于側根起始部位

圖 5 NO 合成酶抑制劑處理對OsNIA1 和OsNIA2 基因表達的影響Fig. 5 The effect of NO synthetase inhibitor treatment on the expression of OsNIA1 and OsNIA2

圖 6 SNP 處理NO 含量及O2·-產生速率的影響Fig. 6 The effect of SNP treatment on NO content and O2·-generation rate
本研究結果證實,水稻側根形成需要NO 信號調控,且需要NO、IAA 和O2·-極性積累于側根的起始部位。當有外源的SNP 供體時,NO 在側根形成部位的熒光強度與對照組相比顯著增強,當受到NO 合成酶抑制劑或NO 清除劑處理后,水稻側根部位的NO 熒光強度顯著減弱,同時IAA 和O2·-在側根起始部位的積累也顯著降低,表明側根數量和長度均受到顯著抑制,當清除NO 時,側根的數量顯著下降[22]。研究結果表明,O2·-和IAA 極性積累于側根起始部位受到NO 信號調節,受NO 供體SNP 處理后,能夠促進IAA 和O2·-極性積累于側根起始部位及根尖,從而誘導側根的發生頻率和快速生長。該研究結果與前人在擬南芥中發現IAA 在側根初始細胞的選擇、根原基發育和側根分生組織激活等過程中均起到關鍵作用的結論基本一致[23]。但這三種信號物質如何選擇主根中的特定中柱細胞作為側根原基并促進側根發育的具體機制,還需深入研究。
3.2 NO 促進IAA 和O2·-積累于側根起始部位利于側根發育
通過對DR5-GUS 標記轉基因水稻IAA 進行GUS 染色發現,IAA 主要分布在初生根中柱部位和側根根尖,而在初生根皮層細胞的含量很少,在無側根發育部位甚至無極性積累。當NO 供體SNP 濃度增加時,側根形成部位NO 含量也隨之增加,同時,IAA 的GUS 染色顯著加深,表明側根形成部位IAA 的積累也相應提高。因此,該研究結果證實了水稻側根發生確實需要IAA 極性積累于側根起始細胞才有利于側根的快速生長,這與前人的研究結果相似[24],且本研究結果也說明IAA 位于NO 的下游信號物質。水稻側根的形成需要NO 調控IAA極性積累于側根起始部位,NO 信號是促進IAA 極性運輸還是加強了IAA 從頭合成再向側根原基的極性積累,還需深入研究。
此外,水稻根受外源NO 供體SNP 處理后,隨著NO 含量增加,側根形成部位及根尖的O2·-極性積累也增加,這與前人發現NO 在調控ROS 在植物體內的代謝平衡有著重要作用的結論一致[25-26]。且有研究表明,NO 通過ONOO-來調控H2O2的信號,這與前人發現水稻種子萌發時受外源O2·-清除劑處理則可顯著抑制初生根生長的結果相似[27-28]。因為已有研究發現,O2·-是植物生長的重要調節信號物質,在生長活躍區都有O2·-產生,當O2·-積累后可誘導SOD 活性增加,SOD 將O2·-轉化成H2O2,再在O2·-和H2O2信號下誘導側根的形成[29]。而且O2·-經SOD 作用產生H2O2的同時也形成.OH 時,.OH可促進細胞壁木糖和果膠的裂解,從而促進細胞壁松動,進而促進細胞生長,當.OH 形成途徑受阻時,玉米根尖伸長區生長明顯受到抑制[30]。所以側根形成及快速生長均需O2·-信號的參與。當水稻幼苗NO 合成受到抑制或受清除時,O2·-在側根的積累量顯著下降,導致側根形成受阻,這與前人在NO 促進其他植物不定根形成的研究結果類 似[31]。當超表達O2·-供體煙酰胺腺嘌呤二核苷酸磷酸氧化酶(Respiratory burst oxidase homologue, Rboh) 時,擬南芥的側根數量明顯增多,當Rboh基因表達受到抑制或突變時,側根數量顯著減少,當利用外源的KI 清除O2·-后,側根數量也顯著降低[14],說明側根形成也需要O2·-積累于側根的起始部位。Orman-Ligeza 等[14]進一步研究表明,外源IAA 和NAA 處理可顯著提高Rboh基因家族表達、Rboh 蛋白在側根原基積累可促進H2O2含量增加,從而誘導側根形成,且外源H2O2還能部分恢復生長素運輸aux1 和lax3 雙突變體的側根數量,由此說明O2·-作為IAA 下游信號物質調控側根的形成,或存在著互作與協同關系[32]。因此,在NO 調控水稻側根形成過程中,可能是NO 先促進IAA 極性積累,再促進O2·-極性積累,從而利于側根形成。但水稻側根形成過程中,NO 信號調控IAA 和O2·-極性積累并促進側根形成的具體生理與分子機制還需要深入研究,特別是中側根原基起始細胞的選擇與NO 調控信號的關系也需深入探討。
4 結論
1)水稻幼苗側根形成需要NO 信號參與,當提供外源的NO 供體SNP 時,可顯著促進IAA 和O2·-極性積累于側根原基起始細胞和側根的尖端,從而促進側根形成。當提供外源NO 合成受到抑制和清除劑時,降低了NO 在側根起始部位的積累,也顯著降低了側根數量。
2)在水稻幼苗側根形成過程中,IAA 可能是NO 的下游信號物質,O2·-則可能是IAA 的下游信號物質,他們共同調控側根的形成。
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
