工廠化養殖紅羅非魚攝食耗氧規律
2022-07-07 02:36:30張瑜霏張宇雷
漁業現代化 2022年3期
張瑜霏,張宇雷
(1上海海洋大學水產與生命學院,上海 201306;2 農業農村部漁業裝備與工程技術重點實驗室,中國水產科學研究院漁業機械儀器研究所,上海 200092)
溶氧是溶解在水中的分子態氧,是魚類賴以生存的必要條件之一。以陸基工廠化為主的養殖生產方式環境相對封閉[1],魚類呼吸代謝所消耗的溶氧主要依靠向水中通入空氣或純氧來補充平衡[2-3]。研究表明,魚類耗氧會受到自身和環境因素的影響而變化,導致魚池溶氧產生波動[4-6]。喬德亮等[7]通過測量白斑狗魚(Esoxlucius)耗氧率發現了白斑狗魚屬于“夜伏晝出”的魚類,日均耗氧顯著高于夜均耗氧,耗氧量隨體質量的增加而升高,隨溫度的上升而增加。戴慶年等[8]發現青石斑魚(Epinephelusawoara)晝夜耗氧差異較大,屬于“晝伏夜出”的魚類,主要在夜間取食。宋協法等[9]發現在20℃、25℃、30℃下斑石鯛(Oplegnathuspunctatus)的耗氧率晝夜差異不明顯,耗氧率在投飼后1~3 h達到峰值,隨后呈現逐漸下降的趨勢。王資生等[10]發現鹽度在26.76時,半滑舌鰨(Cynoglossussemilaevis)的耗氧率最低,鹽度在20.08時,半滑舌鰨的耗氧率最高,鹽度對半滑舌鰨耗氧率有極顯著影響。湯洪芬等[11]研究表明,鲇魚(Silurusasotus)幼魚采用降低代謝水平的方式來適應饑餓環境,饑餓時間越久代謝率越低。強笑野等[12]研究表明,羅非魚應激狀態下耗氧率較試驗前有明顯的上升,其差異隨體質量的增加而減小。總體而言,魚類自身的饑餓、攝食、應激以及外界溫度、鹽度、晝夜交替的變化都會對魚的耗氧造成顯著影響。其中,魚類攝食后耗氧率的變化相對更為明顯和重要, Merino等[13]發現加利福尼亞比目魚(ParalichthysCalifornicus)的攝食耗氧率隨個體的增大而減少,3.2~14.2 g規格的幼魚在喂食期間耗氧量達到峰值,但這一影響不會超過24 h。 Royer等[14]在研究虹鱒(Oncorhynchusmykiss)精準養殖方法指出虹鱒魚攝食后耗氧量是平時的2倍以上,針對這一短期的耗氧變化進行預測,可以實現具有成本效益的供氧自動控制。
為了提高工廠化養殖效率,降低養殖成本,減少養殖風險,本研究對紅羅非魚在不同溫度和投飼率條件下的攝食耗氧量及其恢復情況進行了研究,旨在進一步了解和掌握魚類耗氧規律,為水產養殖溶氧精準控制提供理論依據。
1 材料與方法
1.1 材料
本試驗在中國水產科學研究院漁業機械儀器研究所如東中試基地進行,試驗系統主要由養殖池、豎流沉淀器、泵池、移動床生物濾器、紫外殺菌器、自動投飼機、自動增氧裝置、pH調控裝置以及溫控設備組成,系統工藝如圖1所示。養殖池為直徑1.5 m的圓池,水體量1.2 m3。自動投飼機(自制)投飼精度±1 g。自動增氧裝置(自制)控制精度±0.2 mg/L。pH調控裝置(自制)控制精度±0.2。溫控設備購于貝爾美智能科技有限公司,控溫精度±0.2℃。水溫、溶氧和pH傳感器均安裝在魚池內。為避免大氣交換對試驗參數造成影響,采用硅膠墊覆蓋魚池。試驗魚為紅羅非魚,養殖密度為30 kg/m3,規格為50±2 g,總尾數700尾。
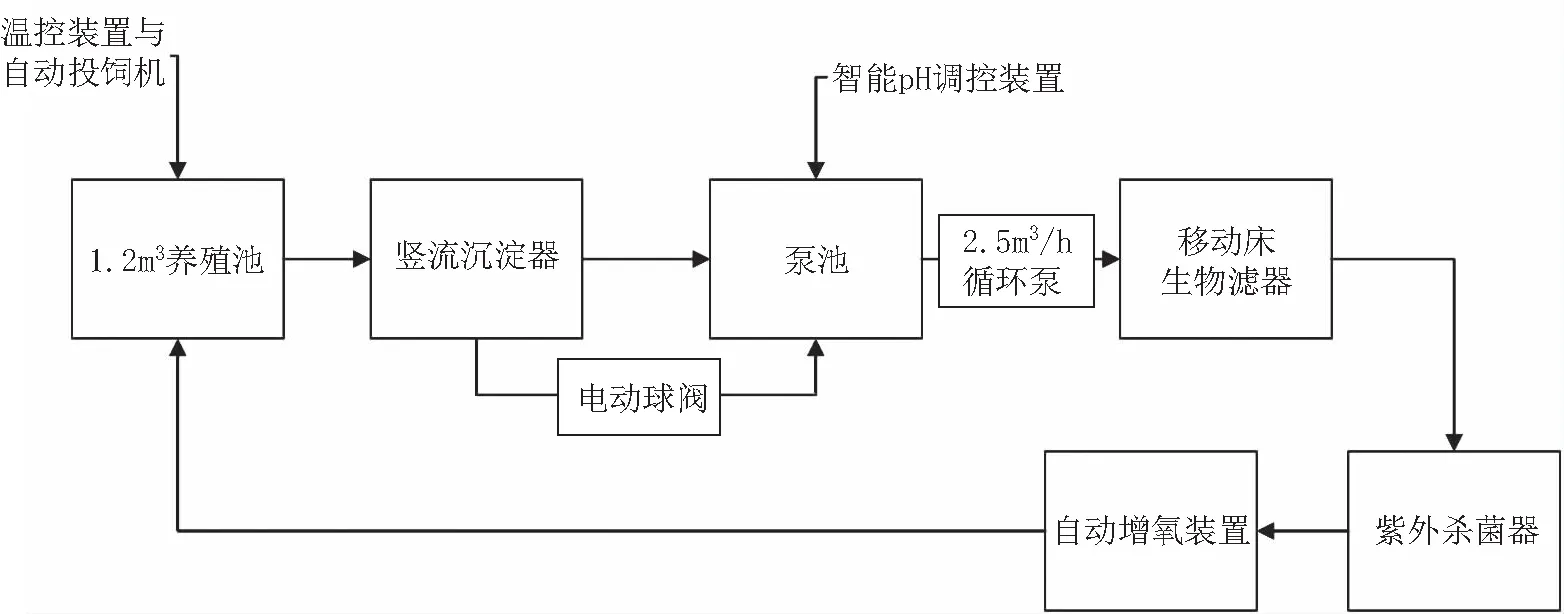
圖1 循環水養殖系統Fig.1 Recirculating aquaculture system
1.2 方法
該試驗使用1套循環水系統,1個養殖池,試驗連續39 d,由于紅羅非魚生長比較緩慢,本研究忽略羅非魚體質量對耗氧量的影響。為避免養殖對象在試驗過程中產生應激,影響數據準確性,本試驗采用靜水法(臨時關閉系統循環),通過記錄養殖池內溶氧變化計算得到羅非魚耗氧量。在關閉系統循環前,控制溶氧質量濃度(7.5±0.2)mg/L、pH(7.2±0.2)。試驗中出現死魚時根據死魚數量及時補充至初始尾數。
試驗設置26℃、29℃、32℃共3個溫度處理組和0.4%、0.8%、1.2%、1.6%、2.0%共5個投飼率的處理組和不投飼的對照組。將不同的溫度和投飼率兩兩組合,測量投飼時和投飼后一段時間的耗氧量,試驗共設30個處理組,每個處理組設置3個重復,研究不同溫度和投飼率組合對魚類攝食耗氧量和攝食后2 h耗氧恢復情況的影響。
每次試驗時間為上午9:00,每天只進行一次試驗以確保魚消化完全。當溶氧從7.5 mg/L降至4.5 mg/L時停止記錄,采樣頻率為1 min,計算攝食耗氧量。繼續開啟系統循環,將溶氧重新調控至7.5 mg/L,2 h后關閉系統,溶氧從7.5 mg/L降至4.5 mg/L時停止記錄,采樣頻率為1 min,計算攝食后2 h的耗氧量。每組試驗完成后將溶氧探頭進行校準。試驗前,提前3 d將水溫調整至試驗水溫,正常投飼使羅非魚適應試驗水溫,待羅非魚游動、攝食正常后用于試驗。
1.3 數據處理
攝食耗氧量:
Mo=(A1-A2) ×V/(M×T)
(1)
式中:Mo—攝食耗氧量(mg/ind.h),A1—試驗前溶氧(mg/L),A2—試驗后溶氧(mg/L),V—魚池水體積(L),M—試驗魚尾數(ind),T—試驗前后時間差值(h)。
以上所得數據導入Microsoft Excel進行整理和作圖,并通過SPSS26.0軟件進行描述性統計、雙因素方差分析、多重比較(Duncan法),顯著性水平為0.05。統計值采用平均值±標準差(Mean±SD)表示。
2 結果與分析
2.1 溫度對羅非魚攝食前耗氧量的影響
分別在26℃、29℃、32℃3個溫度組下對紅羅非魚攝食前耗氧量進行測量,測得3組耗氧量的平均值:16.842±0.425、20.218±0.250、22.606±0.799 mg/ind.h。結果表明不同溫度組耗氧量差異極顯著(P<0.001)。如圖2所示,紅羅非魚攝食前耗氧量隨水溫的升高而增加,32℃組顯著高于26℃組和29℃組,溫度和攝食前耗氧量具有線性關系,可用線性方程y=0.960 7x-7.970 7(n=3,P<0.001,R2=0.990 3)表示。

圖2 不同溫度下的攝食前耗氧量Fig.2 Pre-feeding oxygen consumption at different temperatures
魚類屬于變溫動物,水溫的變化會直接影響魚類的新陳代謝,在合適溫度范圍內,溫度越高,魚類的生理代謝水平也越高,耗氧水平也隨之增強,當超過一定溫度后,組織代謝會進入麻痹甚至停止的狀態,耗氧水平也隨之降低。楊斯琪等[15]運用封閉流水式試驗方法發現,大口黑鱸幼魚(Micropterussalmoides)的耗氧量隨溫度的升高而增加,29℃時到達頂峰,33℃開始下降。李利等[16]通過控制不同的溫度水平測量吉富羅非魚(GIFT)的呼吸情況,證明25℃以上時羅非魚呼吸頻率變化起伏較大,耗氧量與溫度呈線性關系(R2=0.954 9),這與本研究中的現象一致。26℃、29℃、32℃這3個溫度組下,耗氧量隨水溫升高而增加,屬于紅羅非可以適應的水溫,耗氧水平呈現線性關系。
2.2 溫度和投飼率對攝食耗氧量和攝食2 h耗氧量的影響
雙因素方差分析溫度和投飼率對羅非魚攝食耗氧量和攝食后2 h耗氧量的影響見表1。溫度和投飼率對羅非魚攝食耗氧量及攝食后2 h耗氧量有交互作用,且溫度和投飼率對羅非魚攝食耗氧量及攝食后2 h耗氧量的影響有極顯著性差異(P<0.001)。
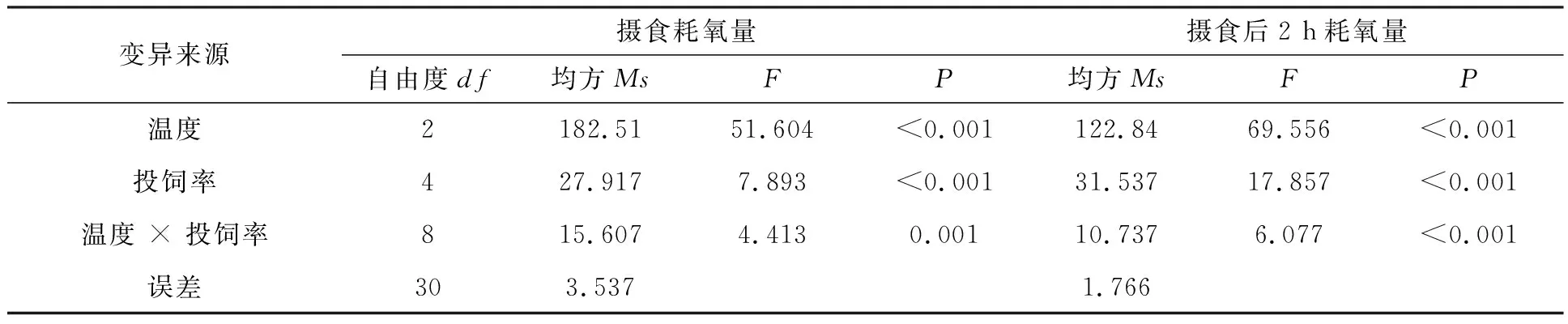
表1 雙因素方差分析溫度和投飼率對紅羅非魚攝食耗氧量和攝食2h耗氧量的影響Tab.1 Two-factor analysis of variance on the effect of temperature and feeding rate on oxygen consumption of red tilapiaand 2 hours after feeding
溫度和投飼率對羅非魚攝食耗氧量及攝食后2 h耗氧量有交互作用,因為溫度是影響魚類食欲和呼吸的主要影響因素之一,在合適的水溫范圍內,溫度越高魚體的生理代謝水平就越高,魚類的攝食耗氧量會隨著溫度的升高而增加,在攝食后其耗氧量的增幅就越高[17-19]。陳松波[20]測量了不同溫度下鯉魚(Cyprinidae)的日攝食率,發現鯉魚成魚在低溫狀態下,攝食率較小,攝食率不隨溫度的升高而改變。幼魚則隨著溫度的升高呈現逐步上升的趨勢,在28℃時達到頂峰,鯉魚成魚和幼魚皆在30℃時攝食率發生驟降。本研究溫度設置在26~32℃之間,耗氧量隨著溫度的升高,耗氧量顯著升高,在投飼率增加的情況下,攝食耗氧量也呈現上升的趨勢,說明26~32℃在羅非魚的適溫范圍內,溫度和投飼率的變化都能對攝食耗氧量產生顯著影響。溫度的升高加快了魚類的攝食代謝,提升了魚類的攝食欲望,從而產生魚類攝食耗氧量隨溫度的升高而增大的趨勢。
圖3顯示不同溫度和投飼率下紅羅非魚攝食耗氧量和攝食2 h耗氧量的測量結果,紅羅非魚攝食后2 h耗氧量顯著低于攝食耗氧量,說明在不同溫度和投飼率下,紅羅非魚在攝食完成2 h后耗氧量均恢復了一定水平。在26℃條件下,攝食耗氧量隨投飼率的增加而上升,到1.6%投飼率時達到頂峰,隨后發生下降,2 h后耗氧量呈現同樣趨勢。在29℃條件下,攝食耗氧量與26℃時趨勢相同,2 h耗氧量隨投飼量呈現逐步上升趨勢未出現下降現象。32℃條件下,耗氧量和投飼率的關系較不穩定,耗氧量呈現先高再降接著逐漸上升的趨勢,0.4%投飼率與1.2%和1.6%差異均不顯著(P>0.05),2 h后耗氧量也比較異常未呈現隨投飼率上升的趨勢。4%投飼率處理下,紅羅非魚攝食耗氧量均呈現隨溫度升高而上升的趨勢,各溫度組之間差異極顯著(P<0.05),2 h后耗氧量呈現同樣趨勢。隨著投飼量的增加,各溫度間的差異逐漸減少,在投飼率1.2%處理下,攝食耗氧量受溫度的影響不顯著(P>0.05),1.6%和2%也未出現耗氧量隨溫度影響呈完全顯著的現象,2 h后耗氧量同樣表現不規律。

圖3 不同溫度和投飼率下紅羅非魚攝食耗氧量 和攝食2h耗氧量Fig.3 Red tilapia postprandial oxygen consumption and2 h postprandial oxygen consumption under differenttemperatures and feeding rates
魚類的饑餓程度決定其代謝強度,當魚類饑餓狀態時會出現和低溫類似的代謝狀態,從而減少能量支出,在出現大量食物時魚類又會處于補償生長期拼命地攝食[21-23]。在以往耗氧量測量中,學者們試驗前通常將試驗魚饑餓3 d,此時魚類耗氧量將低于正常狀態,本試驗在正常運行的循環水系統中測量耗氧量,更接近生產時的魚類耗氧水平。近期有研究報道采用熒光傳感器測量溶氧相對比Winker法誤差更小[24],考慮到Winker法步驟較為煩瑣,試驗采用熒光溶氧傳感器測量魚池溶氧。本研究中的試驗魚,一天只進行一次投飼以保證每次投飼試驗魚都能將飼料吃完,魚類在投飼前皆保持著饑餓狀態,當投飼發生時,魚群搶食激烈,耗氧量較平時上升了1.5~2倍,這一現象在De la Gándara等[25]的報道中也相同體現,黃尾鰤(SeriolaDumerili)在1天2次的投飼頻率下,耗氧量上升顯著,2 h內較投飼前耗氧量上升近2倍。近期蒲德永等[26]對鱸鯉(Percocyprispingipingi)的在不同溫度下的攝食耗氧量進行了研究,其發現在15℃組中,1%與2%的攝食后耗氧量接近,而25℃組中攝食耗氧量隨投飼率的增加而表現增大趨勢,這與本研究的試驗結果基本一致。雷思佳[27]測量了臺灣紅羅非魚的耗氧率,其52.41 g體質量組的耗氧量低于本研究中的攝食耗氧量。金一春等[28]測量的“GTC”羅非魚 287 .40~391 .90 g組耗氧量接近于本研究中紅羅非魚攝食后的耗氧量,紅羅非魚攝食后可以暫時達到大規格魚的水平。本研究中有部分試驗組攝食耗氧量隨投飼率的增加而減小,其原因是投飼率超過了羅非魚最適的攝食溫度所能夠產生的最大攝食率,魚在攝食一段時間后發生了少量嘔吐現象。唐道君等[29]發現黑鱾(Girellamelanichthys)在32℃時,攝食量顯著低于其他溫度組,攝食后的耗氧率也呈現不規律的狀態。在高密度養殖環境下,溶氧難控制,主要是魚類攝食后的特殊動力作用造成的,因此研究魚類攝食耗氧規律對高密度養殖環境下溶氧的穩定具有重大意義。
魚類在攝食完成后耗氧量先呈現急劇上升的趨勢,到達頂峰后逐漸開始下降,直至恢復投飼前的水平。邢道超等[30]的報道中指出云龍石斑魚(Epinephelusgroupers)幼魚的特殊動力作用主要是由蛋白質的代謝作用引起的。在蒲德永等[26]的報道中,鱸鯉的耗氧量在攝食后2~8 h達到頂峰在30 h后耗氧量恢復至投飼前水平,其投飼率越高耗氧量恢復的速度越慢,這與本研究中的研究結果一致,本研究中采集的投飼后2 h的耗氧量,投飼率較少的試驗組攝食耗氧恢復率已經接近投飼前的耗氧量,而較高的試驗組2 h后的耗氧量與投飼前的耗氧量還存在較大的距離。這一現象說明投飼率越高,魚類消化食物的時間越久,耗氧量恢復至投飼前水平所需要的時間越長。
3 結論
溫度變化和投飼是造成循環水系統魚池溶氧波動的主要誘因,魚類攝食前耗氧量與溫度呈線性關系而投飼后則不具備線性關系,但耗氧量仍保留隨溫度升高和增加的特性。魚池溶氧呈現3種狀態:魚類攝食前耗氧狀態、魚類攝食狀態、魚類恢復狀態。這意味著高密度養殖環境下,純氧的輸出流量無法只停留在單一的幅度,而是隨著魚類攝食的狀態而隨之變化。鑒于此,下一步的溶氧智能調控裝備的研發重點是解決溫度變化以及投飼后帶來的溶氧擾動,應用魯棒性較高的智能控制算法通過溫度、溶氧、投飼量等參數的建模和反饋實現主動控制,提前預知溫度變化和投飼參數,及時做出控制響應。
□
