鹽堿脅迫對(duì)甘藍(lán)型油菜生理及分子機(jī)制的影響
2022-07-11 08:07:50楊明煊劉志文
華北農(nóng)學(xué)報(bào) 2022年3期
李 班,呂 瑩,楊明煊,宋 婷,于 放,劉志文
(大連工業(yè)大學(xué) 生物工程學(xué)院,遼寧 大連 116034)
土壤鹽堿化作為世界各國正在面臨著的重大環(huán)境問題,對(duì)于農(nóng)作物的產(chǎn)量和質(zhì)量有著嚴(yán)重的影響,據(jù)不完全統(tǒng)計(jì),全世界鹽堿地的面積為9.5億hm2,其中我國為9 913萬hm2,而且土壤鹽堿化趨勢(shì)也正逐年增加[1-2]。鹽堿脅迫包括鹽脅迫、堿脅迫以及鹽堿共同脅迫,其中,鹽脅迫主要是由中性鹽NaCl和Na2SO4引起,而堿脅迫則是由堿性鹽NaHCO3和Na2CO3引起[3-7]。鹽堿脅迫對(duì)植物帶來的傷害包括滲透脅迫、離子毒害、活性氧脅迫等[8]。在受到鹽堿脅迫的時(shí)候,植物一般會(huì)通過2種途徑來調(diào)節(jié)自身滲透平衡,一種是植物細(xì)胞會(huì)通過吸收外界環(huán)境中的Na+、K+、Cl-等無機(jī)鹽離子來調(diào)節(jié);另一種是植物細(xì)胞通過自身合成的脯氨酸、可溶性糖、甜菜堿、抗氧化酶等物質(zhì)來調(diào)節(jié)[9];而甜菜堿作為植物體內(nèi)無毒的滲透調(diào)節(jié)物質(zhì),它的合成以膽堿為底物,經(jīng)過2步催化形成,一是膽堿加氧酶(CMO),催化膽堿氧化成甜菜堿醛;二是甜菜堿醛脫氫酶(BADH),催化甜菜堿醛形成甜菜堿[10]。在植物受到鹽堿脅迫時(shí),葉綠素含量變化可以反映光合作用變化以及植物對(duì)鹽堿脅迫的響應(yīng)[11]。另外,在受到鹽堿脅迫時(shí),植物體內(nèi)會(huì)產(chǎn)生一種有毒物質(zhì)—丙二醛,丙二醛的過量積累會(huì)影響植物細(xì)胞的正常生理活動(dòng),因此,丙二醛可以作為衡量植物受到鹽堿脅迫強(qiáng)弱的指標(biāo)之一[12]。
甘藍(lán)型油菜(BrassicanapusL.)作為我國重要油料作物,具有長勢(shì)強(qiáng)、產(chǎn)量高、營養(yǎng)豐富、經(jīng)濟(jì)價(jià)值高、耐鹽堿能力強(qiáng)等諸多優(yōu)點(diǎn)[13]。此外,土壤上層所積累的大量沉積性有機(jī)物及土壤中的部分難溶物,都會(huì)在甘藍(lán)型油菜根系所分泌的有機(jī)酸的作用下加速溶解,這不僅能改善土壤,還能減輕地力消耗,兼有養(yǎng)地的作用,因此具有極大的種植潛力[14]。
一直以來,關(guān)于植物耐鹽堿方面的報(bào)道多集中在單一鹽、單一堿對(duì)植物脅迫后其生理生化指標(biāo)方面的研究[15-17],關(guān)于鹽堿共同脅迫及其分子機(jī)制的研究較少,并且在較多研究中,鹽堿脅迫被統(tǒng)稱為鹽脅迫,缺少一定的科學(xué)性。實(shí)際上,在我國大面積的鹽堿環(huán)境中,植物是受到了鹽、堿共同脅迫的影響[18],因此,鹽堿共同脅迫是目前生產(chǎn)實(shí)際中存在的主要問題。本試驗(yàn)主要利用人工模擬鹽脅迫、堿脅迫以及鹽堿脅迫處理甘藍(lán)型油菜,從生理及分子機(jī)制兩方面研究甘藍(lán)型油菜對(duì)鹽堿脅迫的響應(yīng),旨在為提高甘藍(lán)型油菜耐鹽堿能力以及增加甘藍(lán)型油菜產(chǎn)量提供重要的理論依據(jù)。
1 材料和方法
1.1 試驗(yàn)材料
試驗(yàn)材料為甘藍(lán)型油菜華油雜62,由華中農(nóng)業(yè)大學(xué)提供。
1.2 試驗(yàn)方法
1.2.1 混合鹽堿溶液的配制和處理 試驗(yàn)中所用復(fù)合鹽成分為NaCl、Na2SO4,按照1∶1混合;復(fù)合堿成分為Na2CO3、NaHCO3,按照1∶1混合;復(fù)合鹽堿成分為NaCl、Na2SO4、Na2CO3、NaHCO3,按照1∶1∶1∶1混合。共設(shè)置處理組3個(gè)(復(fù)合鹽、復(fù)合堿、復(fù)合鹽堿),對(duì)照組1個(gè)(ddH2O)。每個(gè)處理組設(shè)置3個(gè)梯度,其中復(fù)合鹽包括:Y25(含鹽量25 mmol/L)、Y50(含鹽量50 mmol/L)、Y75(含鹽量75 mmol/L);復(fù)合堿包括:J25(含堿量25 mmol/L)、J50(含堿量50 mmol/L)、J75(含堿量75 mmol/L);復(fù)合鹽堿包括:YJ25(含鹽堿量25 mmol/L)、YJ50(含鹽堿量50 mmol/L)、YJ75(含鹽堿量75 mmol/L)。
1.2.2 種子發(fā)芽率測(cè)定 將50粒籽粒飽滿、大小一致的華雜62種子播種到含有不同人工模擬的復(fù)合鹽、復(fù)合堿和復(fù)合鹽堿溶液的平板上培養(yǎng),萌發(fā)期間規(guī)律性噴灑相應(yīng)溶液,以保證種子發(fā)芽期需要的水分,7 d 后計(jì)算發(fā)芽率,每個(gè)處理設(shè)3 次重復(fù)。
1.2.3 生理指標(biāo)的測(cè)定 為減少試驗(yàn)誤差,本試驗(yàn)中所用幼苗均由一顆華雜62種子經(jīng)NaClO消毒后再經(jīng)植物組織培養(yǎng)進(jìn)行大量擴(kuò)培,隨后將生長30 d且長勢(shì)一致的組培苗移入營養(yǎng)土中,待生長14 d后,取長勢(shì)一致的油菜幼苗進(jìn)行不同濃度的復(fù)合鹽、復(fù)合堿和復(fù)合鹽堿溶液處理,分別處理7,14,21 d后,收集葉片,迅速存于-80 ℃超低溫冰箱中,用于不同生理指標(biāo)的測(cè)定,每個(gè)處理設(shè)3次重復(fù)。
葉綠素含量測(cè)定采用分光光度法,脯氨酸含量測(cè)定采用酸性茚三酮法;可溶性糖含量測(cè)定采用蒽酮比色法,SOD采用氮藍(lán)四唑(NBT)光還原法測(cè)定[19],丙二醛含量測(cè)定采用硫代巴比妥酸法,CAT活性采用過氧化氫法測(cè)定,POD活性采用愈創(chuàng)木酚法測(cè)定[20],甜菜堿含量測(cè)定方法參考文獻(xiàn)[21]進(jìn)行。
1.2.4 實(shí)時(shí)熒光定量分析 通過TRIzol法提取經(jīng)不同條件處理后的甘藍(lán)型油菜幼苗葉片中的總 RNA,利用反轉(zhuǎn)錄酶將所提取的RNA反轉(zhuǎn)錄成cDNA,以PP2A作為內(nèi)參進(jìn)行實(shí)時(shí)熒光定量分析。利用甘藍(lán)型油菜中甜菜堿合成途徑關(guān)鍵酶基因CMO的全序列,通過Primer Premier 5軟件根據(jù)引物設(shè)計(jì)原則設(shè)計(jì)特異性引物,引物序列如表1所示。

表1 引物序列Tab.1 Primer sequences
實(shí)時(shí)熒光定量?jī)x器為LightCycle?480,使用 SYBR GREEN Ⅰ熒光染料,程序如下:95 ℃預(yù)變性5 min;95 ℃變性20 s,50 ℃退火15 s,72 ℃延伸15 s,40 個(gè)循環(huán);72 ℃延伸10 min。反應(yīng)體系:SYBR GREEN Ⅰ 10 μL;Primer 1 0.2 μL;Primer 2 0.2 μL;cDNA 2 μL;ddH2O 7.6 μL,總體系20 μL。將得到的Ct值使用2-ΔΔCt方法計(jì)算,每個(gè)基因測(cè)3組平行,計(jì)算其平均數(shù)以及標(biāo)準(zhǔn)誤差。
1.3 數(shù)據(jù)處理與分析
通過Microsoft Excel 2010軟件進(jìn)行數(shù)據(jù)的計(jì)算和整理,采用 SPSS 軟件對(duì)數(shù)據(jù)進(jìn)行方差分析和最小顯著差異性檢驗(yàn),通過Origin 2018進(jìn)行圖表的繪制。
在所有圖表中,誤差線表示3個(gè)生物學(xué)重復(fù)的標(biāo)準(zhǔn)誤差,不同的小寫字母表示在P<0.05 水平上各試驗(yàn)處理間差異顯著。
2 結(jié)果與分析
2.1 鹽堿脅迫對(duì)甘藍(lán)型油菜種子發(fā)芽率的影響
隨著鹽堿溶液濃度的升高,甘藍(lán)型油菜的發(fā)芽率有所下降,當(dāng)油菜種子受到中高濃度(YJ50、YJ75)鹽堿脅迫影響時(shí),發(fā)芽率為0,其中復(fù)合鹽處理的種子發(fā)芽率與CK組相比變化差異較小,說明復(fù)合鹽對(duì)油菜種子發(fā)芽率影響最小,其次為復(fù)合堿的傷害,而復(fù)合鹽堿對(duì)種子發(fā)芽率影響程度最大,且達(dá)顯著水平(P<0.05)(表2)。
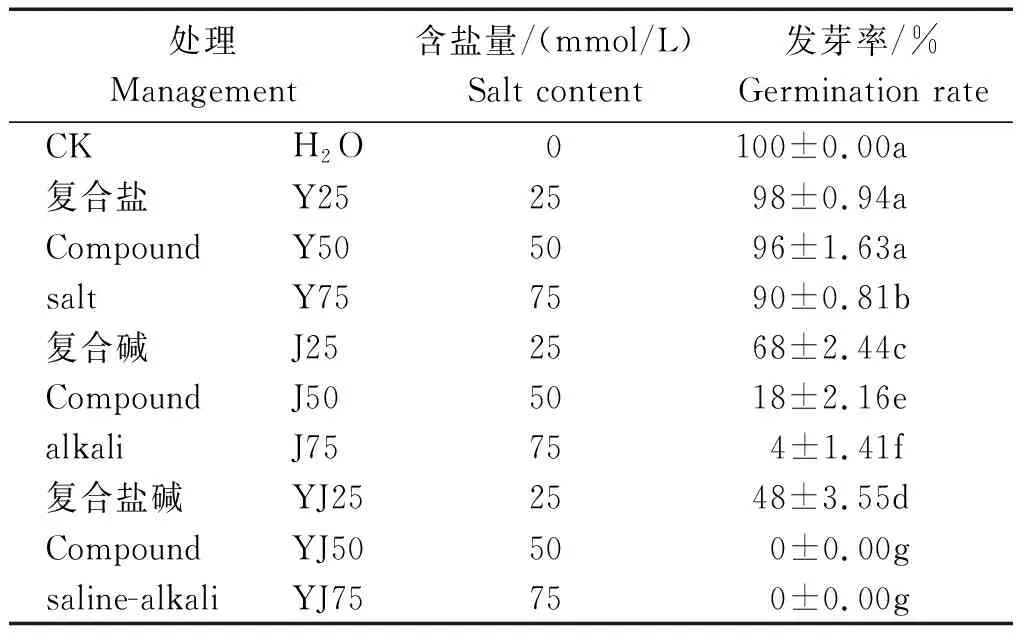
表2 鹽堿脅迫對(duì)種子發(fā)芽率的影響Tab.2 The effect of saline-alkali stress on the seed germination
2.2 甘藍(lán)型油菜耐鹽堿機(jī)制
2.2.1 葉綠素含量 如圖1所示,在經(jīng)低中復(fù)合鹽(Y25、Y50)處理油菜幼苗時(shí),葉綠素含量隨處理時(shí)間延長先升高后降低,當(dāng)處理時(shí)間為14 d,葉綠素的含量最高,分別為16.53,16.67 mg/g,在經(jīng)過高復(fù)合鹽(Y75)、復(fù)合堿、復(fù)合鹽堿溶液處理后,葉綠素含量隨著處理濃度增加與處理時(shí)間的延長而下降。因此,低濃度鹽堿溶液會(huì)促進(jìn)葉綠素的積累,而高濃度鹽堿溶液會(huì)抑制葉綠素的積累,其中以復(fù)合堿與復(fù)合鹽堿對(duì)葉綠素的影響最為顯著(P<0.05)。

不同小寫字母表示差異顯著(P<0.05)。圖2—11同。Different lowercase letters indicate significant difference(P<0.05).The same as Fig.2—11.
2.2.2 脯氨酸和可溶性糖含量 如圖2,3所示,隨著鹽堿溶液濃度升高與處理天數(shù)的增加,葉片中脯氨酸及可溶性糖含量顯著增加,當(dāng)脅迫處理21 d,中高復(fù)合鹽(Y50、Y75)與高復(fù)合鹽堿(YJ75)處理組中脯氨酸含量急劇升高,分別是對(duì)照組的12.19,12.97,65.99倍,差異顯著(P<0.05);高復(fù)合鹽堿(YJ75)組中可溶性糖大量積累,脅迫21 d可溶性糖積累量高達(dá)40.60 mg/g,為對(duì)照組的5.21倍。

圖2 復(fù)合鹽、復(fù)合堿、復(fù)合鹽堿對(duì)甘藍(lán)型油菜葉片脯氨酸含量的影響Fig.2 Effect of compound salt,compound alkali,compound saline-alkalion the proline content in leaves of Brassica napus L.

圖3 復(fù)合鹽、復(fù)合堿、復(fù)合鹽堿對(duì)甘藍(lán)型油菜葉片可溶性糖含量的影響Fig.3 Effect of compound salt,compound alkali,compound saline-alkalion the soluble sugar content in leaves of Brassica napus L.
2.2.3 丙二醛含量 如圖4所示,在復(fù)合鹽、復(fù)合堿、復(fù)合鹽堿溶液脅迫油菜7 d,其體內(nèi)丙二醛含量變化差異較小,在中高復(fù)合鹽(Y50、Y75)處理幼苗14,21 d,丙二醛含量要高于對(duì)照組,且差異顯著;脅迫21 d,高復(fù)合堿(J75)處理油菜幼苗后的丙二醛含量要高于高復(fù)合鹽(Y75)處理后的丙二醛含量;復(fù)合鹽堿對(duì)油菜幼苗丙二醛含量的影響趨勢(shì)與復(fù)合鹽、復(fù)合堿的影響趨勢(shì)相似。
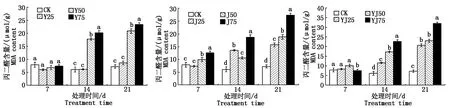
圖4 復(fù)合鹽、復(fù)合堿、復(fù)合鹽堿對(duì)甘藍(lán)型油菜葉片丙二醛含量的影響Fig.4 Effect of compound salt,compound alkali,compound saline-alkalion malondialdehyde content in leaves of Brassica napus L.
2.2.4 抗氧化酶活性 由圖5可知,復(fù)合鹽處理可顯著降低SOD活性;在復(fù)合鹽處理7 d,POD的活性隨著鹽濃度的升高而升高,與空白對(duì)照相比,高鹽(Y75)處理后的POD活性提高了3.03倍,且隨處理時(shí)間不同,POD活性先升后降,在第14 天達(dá)到最高值;在復(fù)合鹽處理7 d,處理組中CAT活性均高于對(duì)照組,差異顯著(P<0.05),在脅迫14 d,CAT活性隨著鹽濃度呈先降后升趨勢(shì),且隨處理時(shí)間的增加,CAT的活性降低。

圖5 鹽脅迫對(duì)甘藍(lán)型油菜葉片抗氧化酶活性的影響Fig.5 Effect of salt stress on antioxidant enzyme activity in leaves of Brassica napus L.
如圖6所示,復(fù)合堿處理后,處理組中SOD活性均低于對(duì)照組;在脅迫14 d,復(fù)合堿處理后的POD活性達(dá)到最大值,分別為286.5,349.25,302.12 U/(g·min);復(fù)合堿處理后的CAT活性隨處理時(shí)間延長與處理濃度增加而顯著降低(P<0.05)。

圖6 堿脅迫對(duì)甘藍(lán)型油菜葉片抗氧化酶活性的影響Fig.6 Effect of alkali stress on antioxidant enzyme activity in leaves of Brassica napus L.
如圖7所示,在處理第7天,處理組SOD活性要高于對(duì)照組,隨著復(fù)合鹽堿溶液濃度及處理天數(shù)增加,SOD活性降低;復(fù)合鹽堿處理提高了POD活性,與對(duì)照相比,高鹽堿(YJ75)處理21 d后的POD含量提高了2.26倍,差異顯著(P<0.05);處理組中,CAT活性隨著復(fù)合鹽堿的升高而呈先升后降的趨勢(shì),而隨著脅迫時(shí)間的持續(xù),CAT的活性呈先降后升的趨勢(shì),處理組中CAT活性均低于對(duì)照組,差異顯著。

圖7 鹽堿脅迫對(duì)甘藍(lán)型油菜葉片抗氧化酶活性的影響Fig.7 Effect of saline-alkali stress on antioxidant enzyme activity in leaves of Brassica napus L.
各項(xiàng)生理指標(biāo)結(jié)果均表明,鹽堿脅迫對(duì)油菜幼苗的傷害程度大小表現(xiàn)復(fù)合鹽堿>堿>鹽。
2.3 甜菜堿合成途徑中關(guān)鍵酶基因表達(dá)量
由圖8可知,復(fù)合鹽處理7 d,高鹽(Y75)處理后的CMO基因表達(dá)量達(dá)到最高值,為空白對(duì)照的3.14倍,而在處理14,21 d,CMO基因的表達(dá)量隨著鹽濃度的升高而上升,差異顯著(P<0.05);圖9,10可以看出,復(fù)合堿與復(fù)合鹽堿溶液顯著提高了CMO基因的表達(dá)量;在復(fù)合鹽、堿、鹽堿溶液處理21 d后,高鹽(Y75)、高堿(J75)、高鹽堿(YJ75)處理組中CMO基因相對(duì)表達(dá)量分別提高7.21,9.23,13.76倍,差異顯著(P<0.05);同濃度的復(fù)合鹽堿處理后的葉片中的CMO基因表達(dá)量要高于復(fù)合鹽與復(fù)合堿處理組。

圖8 鹽脅迫對(duì)合成甜菜堿關(guān)鍵酶基因相對(duì)表達(dá)量的影響Fig.8 Effect of salt stress on relative expression of key enzyme genes in betaine synthesis
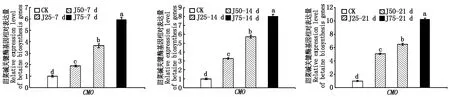
圖9 堿脅迫對(duì)合成甜菜堿關(guān)鍵酶基因相對(duì)表達(dá)量的影響Fig.9 Effect of alkali stress on relative expression of key enzyme genes in betaine synthesis

圖10 鹽堿脅迫對(duì)合成甜菜堿關(guān)鍵酶基因相對(duì)表達(dá)量的影響Fig.10 Effect of saline-alkali stress on relative expression of key enzyme genes in betaine synthesis
在油菜幼苗受到鹽堿脅迫后,關(guān)鍵酶基因CMO的相對(duì)表達(dá)量隨著復(fù)合鹽、堿、鹽堿溶液濃度的升高而上升,且脅迫時(shí)間越長差異越顯著。
2.4 甜菜堿積累量
如圖11所示,甜菜堿的含量隨復(fù)合鹽、堿、鹽堿的濃度及脅迫天數(shù)的增加而上升,且差異顯著(P<0.05)。在高濃度的鹽(Y75)、高濃度的堿(J75)、高濃度的鹽堿(YJ75)脅迫最高天數(shù)(21 d)時(shí),油菜葉片中的甜菜堿的積累量達(dá)到了最高值,分別為7.99,9.61,15.00 mg/g。

圖11 甘藍(lán)型油菜葉片中甜菜堿含量Fig.11 The betaine content in leaves of Brassica napus L.
3 結(jié)論與討論
土地鹽堿化是世界范圍內(nèi)普遍存在的一個(gè)問題,嚴(yán)重影響著植物的生長和發(fā)育[22]。種子萌發(fā)代表著植物生命的開始,在種子萌發(fā)階段,其發(fā)芽率與植物耐鹽堿能力密切相關(guān)。本試驗(yàn)中,油菜種子的發(fā)芽率隨著各溶液濃度的升高呈下降趨勢(shì),其中中高濃度的復(fù)合鹽堿脅迫(YJ50、YJ75)對(duì)油菜種子的傷害最大,而復(fù)合鹽溶液對(duì)種子發(fā)芽率的影響最小,說明鹽堿脅迫對(duì)種子的傷害程度遠(yuǎn)大于單鹽、單堿對(duì)種子的傷害。
可溶性糖和脯氨酸可維持細(xì)胞代謝平衡、細(xì)胞內(nèi)穩(wěn)態(tài)和細(xì)胞膜穩(wěn)定性,當(dāng)植物受到鹽堿脅迫時(shí),脯氨酸、可溶性糖、甜菜堿等滲透調(diào)節(jié)物質(zhì)發(fā)揮了極其重要的作用[23]。本試驗(yàn)中,脯氨酸、可溶性糖、甜菜堿均隨著鹽堿溶液濃度及脅迫天數(shù)的增加而呈上升的趨勢(shì)。
本試驗(yàn)中,葉綠素的含量對(duì)中低濃度復(fù)合鹽溶液(Y25、Y50)表現(xiàn)為隨處理時(shí)間延長呈現(xiàn)先升高后降低的趨勢(shì),可能是因?yàn)榇藭r(shí)油菜葉片中的脯氨酸、可溶性糖、甜菜堿等滲透調(diào)節(jié)物質(zhì)發(fā)揮了重要作用,修復(fù)了油菜葉片所受到的損傷,有效抑制了葉綠素的降解,在經(jīng)過復(fù)合堿、復(fù)合鹽堿及高復(fù)合鹽(Y75)溶液處理后,葉綠素含量隨著處理濃度與處理時(shí)間的升高而下降。
有研究表明,丙二醛含量是隨著鹽濃度或脅迫時(shí)間的增加呈上升趨勢(shì)[24],這一觀點(diǎn)與本研究結(jié)果一致。本試驗(yàn)中,鹽堿脅迫顯著提高了油菜葉片丙二醛含量,而在鹽堿脅迫7 d,丙二醛含量變化差異較小,說明短時(shí)間的鹽堿脅迫對(duì)甘藍(lán)型油菜幼苗所產(chǎn)生的傷害較低,可能是因?yàn)楫?dāng)油菜幼苗因丙二醛的積累而受到活性氧的傷害時(shí),SOD、CAT、POD作為酶類保護(hù)體系開始增加,其中POD發(fā)揮作用最大。
甜菜堿合成途徑中的關(guān)鍵酶基因CMO的相對(duì)表達(dá)量隨著鹽堿溶液的濃度升高以及脅迫時(shí)間延長而上升,CMO會(huì)將膽堿氧化成甜菜堿,因此甜菜堿的含量也會(huì)隨之升高,在復(fù)合鹽、堿、鹽堿溶液處理21 d后,與對(duì)照組相比,高鹽(Y75)、高堿(J75)、高鹽堿(YJ75)處理組中CMO基因的相對(duì)表達(dá)量顯著提高,且此時(shí)甜菜堿的積累量也達(dá)到了最高值,因此在甘藍(lán)型油菜受到鹽堿脅迫時(shí),甜菜堿會(huì)大量積累以減輕油菜所受的傷害。
綜上所述,復(fù)合鹽、堿、鹽堿溶液對(duì)甘藍(lán)型油菜的傷害程度大小表現(xiàn)為復(fù)合鹽堿>堿>鹽;低濃度復(fù)合鹽溶液對(duì)甘藍(lán)型油菜種子的發(fā)芽率影響較低,且短時(shí)間復(fù)合鹽處理對(duì)葉綠素含量有所增加,說明適當(dāng)濃度鹽溶液對(duì)甘藍(lán)型油菜的生長具有一定的促進(jìn)作用;鹽堿溶液會(huì)提高甘藍(lán)型油菜幼苗中的可溶性糖、脯氨酸、甜菜堿等滲透調(diào)節(jié)物質(zhì)以降低鹽堿脅迫對(duì)油菜所帶來的傷害;丙二醛的積累會(huì)使甘藍(lán)型油菜細(xì)胞器發(fā)生損傷,影響其生長,此時(shí)抗氧化酶起到了重要作用:清除活性氧,提高植物的耐鹽堿性,其中以POD發(fā)揮的作用最大;CMO作為甜菜堿合成途徑中的關(guān)鍵酶基因,它的增加促進(jìn)了甜菜堿的積累,而抵御甘藍(lán)型油菜所遭受的鹽堿脅迫所帶來的傷害。以上結(jié)果為甘藍(lán)型油菜耐鹽堿品種選育和改良提供了一定的理論基礎(chǔ)。
