海南清瀾港紅樹植物海蓮種群結構與分布格局研究
2022-07-12 07:23:24李茂金葉儲民陳玉凱
熱帶林業 2022年2期
關鍵詞:紅樹林
李茂金,葉儲民,陳玉凱
1.海南省林業集團有限公司,海南海口 570205;
2.熱帶島嶼生態學教育部重點實驗室,海南師范大學生命科學學院,海南海口 571158
種群結構不僅可以反映種群個體屬性特征的數量分布狀況,也可以反映種群數量動態及種群的發展趨勢,同時可以在很大程度上反映種群與生境間的相互作用關系,以及在群落中的生態作用和生態位。種群的分布格局與種群自身的特性有極大的關系,同時也受種群間效應、生境條件的直接影響。通過對群落中種群分布格局的調查,可以揭示群落空間結構的特征,有助于了解種群的某些生物學和生態學特性,以及解釋一定環境因子對種群行為或者種群生存的作用。調查得出的分布格局結論和數量特征指標,可以作為確定抽樣方式、數據處理提供理論基礎。在生產和開發利用上,對于紅樹林的經營管理以及混交林的配置具有重要參考價值[1-2]。
海蓮隸屬紅樹科木欖屬,常綠喬木,是紅樹林東方類群樹種,在中國境內,海蓮僅分布在海南省,近年來引種于廣東省深圳海灘;在世界上它分布于印度、越南、泰國、緬甸、菲律賓、印度尼西亞等國家的濕熱海灘。
目前對海蓮的研究已有較多的報道,如,從生理生態方面的研究有:鄭文教[3]等研究了鹽度對海蓮幼苗的生長和某些生理生態特性的影響;馬麗[4]等研究了鋁脅迫對海蓮幼苗保護酶系統及脯氨酸含量的影響。從種群和群落方面的研究有:王佳燕[5]等對海南東寨港海蓮群落林隙結構及其更新特征的研究,結果表明:海南東寨港河港海蓮群落最主要的干擾類型是樹木的大量枯死;林隙幼苗密度低于林下幼苗的密度,而林下幼苗的高度低于林隙幼苗的高度,由此可知,適當的林隙可以促進林分更新;廖寶文[6]等對海南島東寨港幾種紅樹植物種間生態位的研究表明:作為紅樹科的植物種之一,海蓮有其特殊的生長環境要求,只適生于底質較為硬實的半泥沙質的高潮灘涂,海蓮生態位寬度值最小,高潮灘灘涂植物海蓮、角果木與其它樹種的生態位重疊值均相對較低(其他樹種的生境可能與海蓮、角果木的生境比較相似的原因),與實際情況相符。海蓮這一紅樹植物為嗜熱窄布種,一般生于潮水較淺、土質比較硬實、陽光充足的泥沙灘涂生境,主要分布在熱帶紅樹林林區的高潮灘涂帶,與中低潮灘的紅樹植物占據相近或相似生態空間的程度不大,所隱含的競爭現象也比較小;鄭松發[7]等對海蓮群落和木欖群落主要種群分布格局的研究表明:海蓮種群在某種程度上具有集群性,但是強度較低,可視作隨機分布和集群分布的過渡類型,分布格局類型主要與各自的群落內生境的微地形變化有關。
海蓮種群是海南省文昌市清瀾港紅樹林林區內的主要優勢種群之一,該研究的目的,是通過探討清瀾港紅樹林林區內優勢植物海蓮種群的結構特征、分布狀況及其發展趨勢,闡明海蓮種群空間分布格局與環境關系,以期為海蓮及其他優勢紅樹種群的人工經營及種群的合理保護提供參考依據。
1 研究區概況
海南清瀾港紅樹林自然保護區是中國最大的紅樹林區之一,保護區總面積2948hm2,其中紅樹林面積1223.3hm2,位于海南省東北部文昌市界內,地理坐 標 為19°15′N ~20°09′N,110°30′E ~110°02′E,屬熱帶季風海洋性氣候[8]。區域內的清瀾港港灣深入內陸,形成了內寬口窄的漏斗狀,在文昌江和文教河匯入灣內,沿岸積累的淤泥深厚,風浪微弱,為典型的瀉湖-河口濕地生境,很適宜紅樹林的生長。
文昌清瀾港紅樹林保護區主要分布在八門灣的沿岸灘涂帶,并且斷斷續續地分布到港灣外,港灣沿岸陸地為沿海臺地地貌,原生的植被類型是熱帶雨林,但受人類經濟活動干擾,絕大部分已開發改成為蝦塘、椰樹、桉樹林、木麻黃林及其他果樹林[8]。
2 研究方法
2.1 取樣方法與野外調查
2020 年8 月到9 月,根據海南文昌清瀾港紅樹林自然保護區海蓮種群的結構現狀及其生境特點,共選擇了3 個樣地進行調查研究,編號分別為P1、P2、P3,樣地概況見(表1)。其中,樣地1 位于八門灣內與清瀾鎮交界處,樣地2 位于八門灣內與東郊鎮交界的沿海臺地,樣地3 位于文昌北部的鋪前鎮珠溪河沿岸。該研究以海蓮種群為目標物種進行調查,在調查區域內,由于紅樹林多為帶狀分布,植物種類也較少,樣方面積不好做大,因此三個樣地取的樣面積為900m2,共2700m2,將每個樣地分成50m2的棋盤格子樣方,各18 個,并采用每木調查法,測量每個樣地內樹種的個體的相對坐標、胸徑、冠幅、高度等特征。
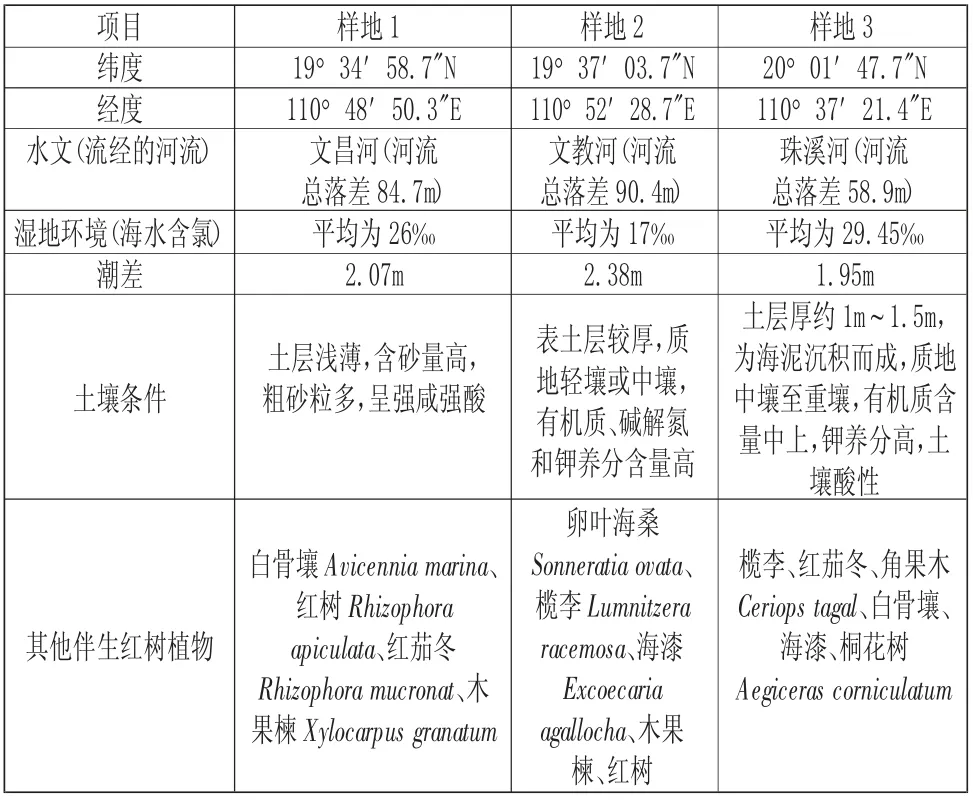
表1 樣地概況Tab.1 Plot Characteristics
2.2 種群結構的研究方法
根據海蓮種群的生長特點,對各樣地中海蓮種群每木調查所得的數據進行分組統計,其中冠幅直徑的組間距為1.0m,高度的組間距為2.0m[9-10],由于種群徑級結構的相對穩定性類似于年齡結構的穩定,所以許多學者都采用胸徑結構來代替年齡結構進行分析喬木種群的結構及其動態特征[11]。該文對海蓮的胸徑結構進行3 個年齡級的分析,即(幼齡)0<胸徑≤7.5cm、(中齡)7.5<胸徑≤22.5cm、(老齡)胸徑>22.5cm 以上。
2.3 種群樹高、胸徑和冠幅的相關性分析
通過對樣地進行每木調查,得到清瀾港紅樹林海蓮種群冠幅直徑、胸徑、樹高的基本實驗數據,對數據進行處理,首先通過利用excel 分別作出冠幅直徑與胸徑、樹高與胸徑的散點圖,然后根據所得各散點圖結合5 種研究模型[12-13],對冠幅直徑與胸徑、樹高與胸徑進行相關性分析;利用SAS 軟件對各組調查所獲得的數據進行模型擬合后,得到各方程的待定參數及各回歸方程及其方程決定系數R2,以決定系數R2大小來評價其擬合程度大小,R2小,說明擬合程度差,模型則不夠可靠。反之,R2大說明擬合程度好,模型可靠。
2.4 種群分布格局分析
以各樣地內海蓮個體數據為依據,對三個樣地調查得到的數據進行方差均值 (亦稱擴散系數)的t 檢驗,以二項分布的聚塊指數(m*/m)、平均擁擠度(m*),Morisita 指數的F 檢驗,判定海蓮種群不同樣地和不同年齡級的空間分布格局[14-15]。
3 結果與分析
3.1 海蓮種群結構
由表2 可知,海蓮種群的個體高度主要集中在0m~6m,占67%,其中各樣地的優勢高度以及個體比例分析結果如下:在樣地P1 中,高度在6m~8m 范圍的個體占65%;在樣地P2 中,高度在2m 以下的個體占58%;在樣地P3 中,高度在2m 以下的個體占26%,2m<高度≤4m、4m<高度≤6m 和6m<高度≤8m 的個體所占比例也較多,分別是24%、22%和22%。這三個樣地的自然環境不同,各樣地的整體外觀有所差異,樣地P1 處于中齡的植株較多,樣地P2 處于幼齡的植株較多,所以兩地的高度有一定的差異,樣地P3 各個年齡的植株都有一定的數量,高度差異介于樣地P1 和P2 兩者之間。
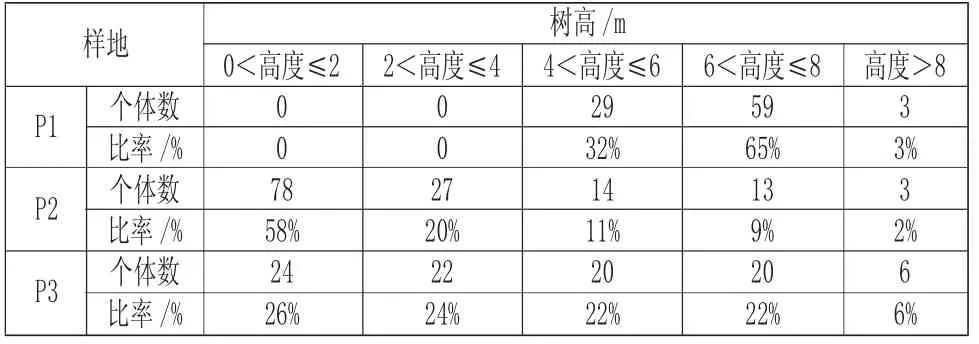
表2 各樹高級分布Tab.2 Distribution of Different Tree Height Grades
由表3 可知,海蓮種群的個體胸徑主要集中在2.5cm~7.5cm,占63%,經過分析,各樣地的優勢胸徑及個體比例如下:在樣地P1 中,胸徑大小在7.5cm~22.5cm 范圍內的個體占54%;在樣地P2 中,胸徑大小在7.5cm 以下的個體占84%;樣地P3 中,胸徑大小在7.5cm 以下的個體占59%。由此可知,在三個調查樣地中,樣地P1 的中齡海蓮植株所占比例是最大的,說明在過去一段時間內,P1 樣地林分發生變化,P1 樣地中海蓮種群呈現了增長趨勢,老齡和幼齡植株相對較少,種群的年齡結構格局趨向于中間大兩頭小,P2 樣地中海蓮種群的自然更新能力最強,樣地中存在大量的幼齡植株,老齡和中齡植株數量很少,說明樣地P2 的環境相對較適合海蓮的更新和發育,樣地P3 的中老齡植株和幼齡植株比例相差不大,種群表現出較穩定的發展趨勢。
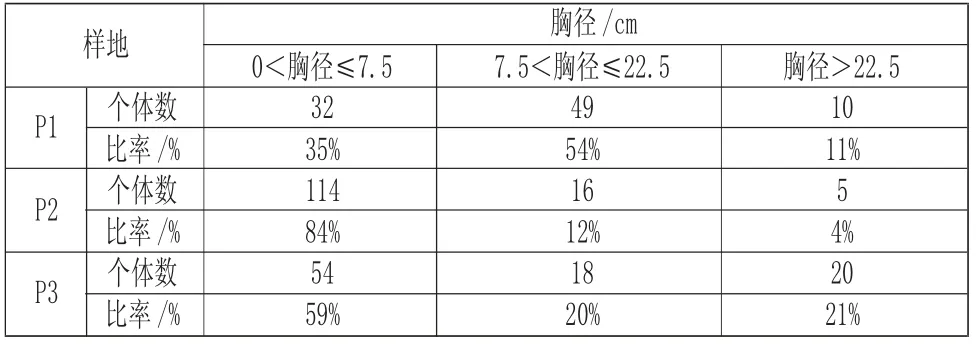
表3 各胸徑級分布Tab.3 Distribution of Different DBH Grades
由表4 可知,海蓮種群的個體冠幅直徑主要集中在0m~3m,占66%,其中各樣地的優勢冠幅直徑及個體比例如下:樣地P1,4<冠幅直徑≤5m 和冠幅直徑>5m 以上的個體較多,均占30%;樣地P2,1m 以下的個體占54%;樣地P3,1m~2m 的個體占52%。樣地P1、P2、P3 的冠幅直徑最大值的個體分別是6.5m、7m、5m;樣地P1 和P3 分別缺失0m~1m 和4m~5m 的個體。
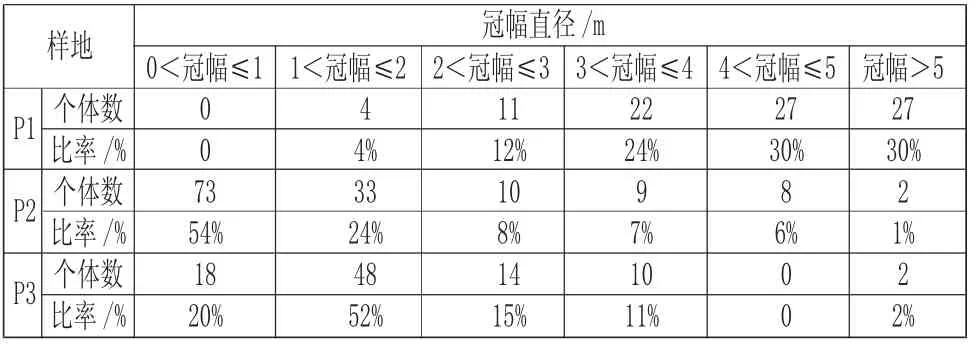
表4 各冠幅直徑級分布Tab.4 Distribution of Different Crown Diameter Grades
由此可知,樣地P2 的自然環境條件更有利于海蓮種群的自然更新,而樣地P1 的自然環境條件對海蓮種群發展不利,有趨向中間大兩頭小的格局,這也說明了不同生境條件和不同立地條件對海蓮種群的作用強度以及種群適應特征有所不同,海蓮種群結構的差異與區域環境特點有較大的相關關系,由此反映出生境差異對種群結構的影響作用。
3.2 海蓮種群樹高、冠幅直徑和胸徑的相關性分析
3.2.1 種群樹高與胸徑的相關性分析
由圖1 分析可以看出,海蓮的樹高具有隨著胸徑增大而增大的現象,觀察數據可知,樹高高度主要集中在3m~6m 的范圍內,相應的胸徑大小范圍在10cm 以內,說明所調查的海蓮林分年齡偏低;經相關數據分析,海蓮樹高與胸徑之間有顯著相關性,相關系數為0.6829,而由表5 中海蓮樹高與胸徑各模型擬合結果可知,五種模型中y=2.4202Ln (x) +0.1578、y=0.2013x+3.2385 和y=2.8341+0.2722x-0.0020x2模型的決定系數相差不大,但模型y=2.8341+0.2722x-0.0020x2擬合效果最好,決定系數最高,利用此模型對海蓮胸徑估算其樹高結果可靠,此模型可為調查海蓮種群提供一定依據。由于紅樹林潮灘通行困難,此模型也可以減輕野外測高工作量,并可了解和進一步掌握海蓮種群的生長發育過程。
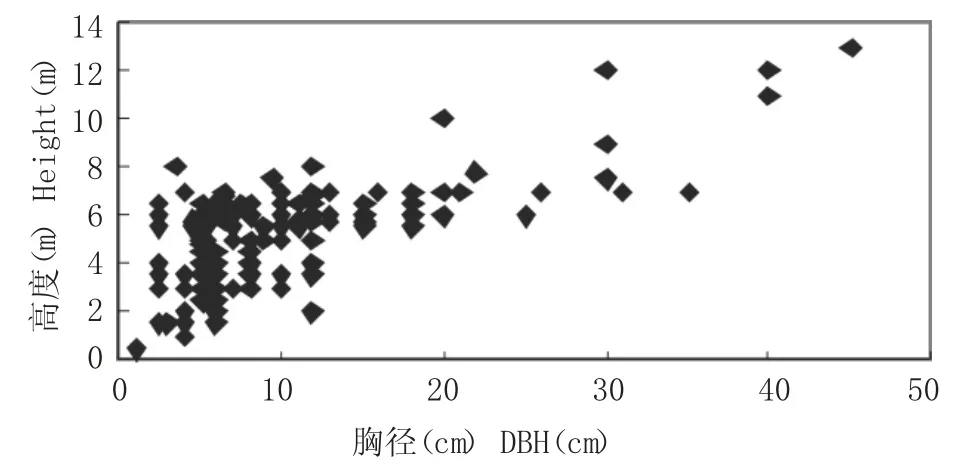
圖1 樹高與胸徑散點Fig.1 Scatter of Height and DBH
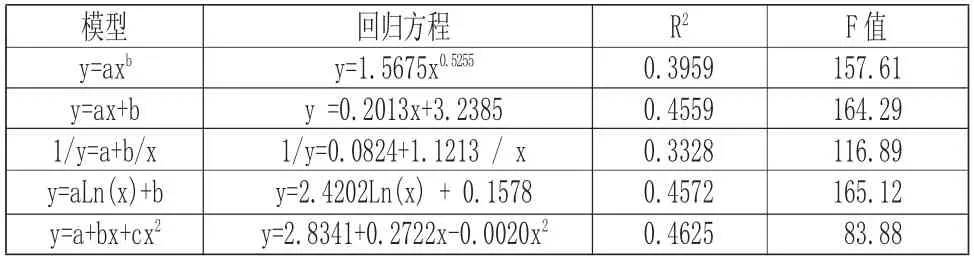
表5 樹高與胸徑關系數學模型及顯著性檢驗Tab.5 Mathematical Model and Test of Significance of Height and DBH
3.2.2 種群冠幅直徑與胸徑的相關性分析
由圖2 分析可以看出,海蓮種群的冠幅直徑具有隨胸徑的增大而增大的現象,觀察數據可知,冠幅直徑以1m~3m 的范圍內最為密集,相應的胸徑在10cm 范圍內,經相關分析,海蓮胸徑與冠幅直徑有顯著相關性,相關系數為0.5492。根據表6 中海蓮冠幅直徑與胸徑各模型擬合結果可知,五種模型中w=0.7218x0.5936和1/w=0.0997+2.7214/x 模 型 的 決定系數相差不大,但模型1/w=0.0997+2.7214/x 擬合效果最好,決定系數最高,利用此模型對海蓮胸徑故算其冠幅直徑結果可靠。根據海蓮種群的胸徑與冠幅直徑大小的相關關系,可以編制林分經營密度表,并且可以適時合理的對海蓮林分進行撫育間伐。并通過對海蓮種群的冠幅直徑與胸徑生長因子之間的相互關系分析,為進一步研究海蓮林種群結構、自然演替和生長發育規律等生態學規律提供可靠的依據。
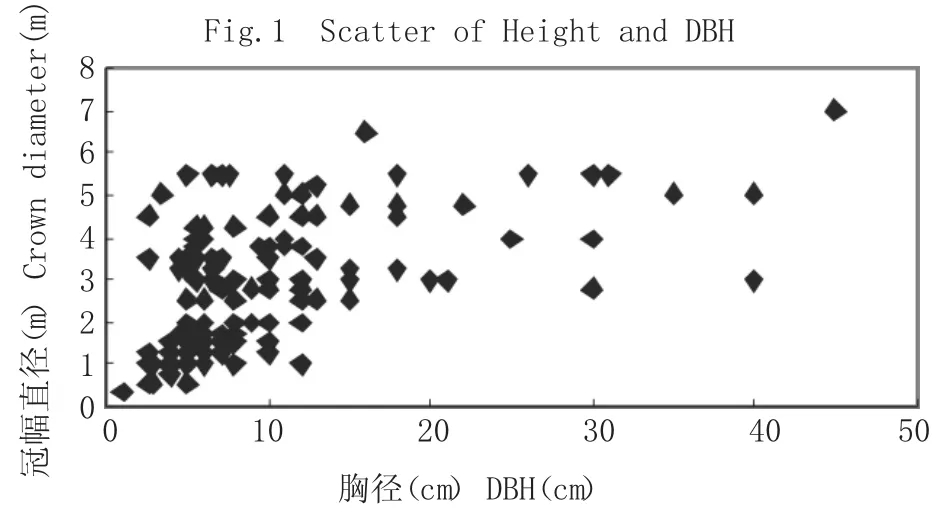
圖2 冠幅直徑與胸徑散點Fig.2 Scatter of Crown Diameter and DBH
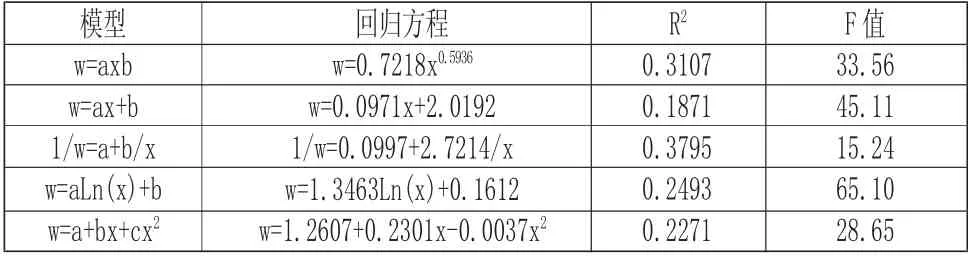
表6 冠幅直徑與胸徑關系數學模型及顯著性檢驗Tab.6 Mathematical Model and Test of Significance of Crown Diameter and DBH
3.3 海蓮種群分布格局
種群的空間格局也是種群結構的重要特征之一,它反映的是種群在水平空間上的配置格局或者是分布狀況,其形成原因是由種群特性、種間關系以及種群所在的環境條件共同起作用所決定。
由表7 可知,所調查研究的三個樣地中,海蓮的種群都呈現出集群分布的狀況,其中樣地P2 的平均擁擠度以及聚塊指數都達到了最高,說明樣地P2 的這種分布格局在一定程度上有利于種群形成幼苗生長的小環境,同時這種分布格局也可以增強種群抵抗其它植物入侵的抗性,增加種群在群落中的競爭力以及種群對營養資源和生存空間資源的占據與鞏固。
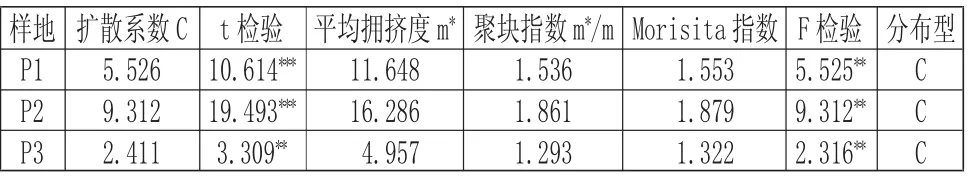
表7 不同樣地海蓮種群分布格局的測定結果Tab.7 Result of Spatial Distribution Patterns of Bruguiera sexangula Population in Different Plots
由表8 可知,海蓮種群在不同發育階段,種群的分布格局大體上呈現出集群型向隨機型轉變的趨勢,并且各集群強度由大變小,趨向于減弱,種群中幼齡樹集群分布,且聚集程度很高;中齡樹為集群分布,聚集程度有所下降,老齡樹逐漸趨于隨機分布。但樣地P3 的幼齡聚集強度較弱,明顯低于P1 和P2,這是因為樣地P3 已受到人為的干擾,而幼齡個體更易受人類活動的影響,因此有趨向隨機分布的可能,而P2 的幼齡聚集強度非常高,明顯高于P1 和P3,這種格局的形成是因為樣地P2 的幼齡植株個體數量比較多,樣地P2 中可以經常見到十幾株簇擁生長。
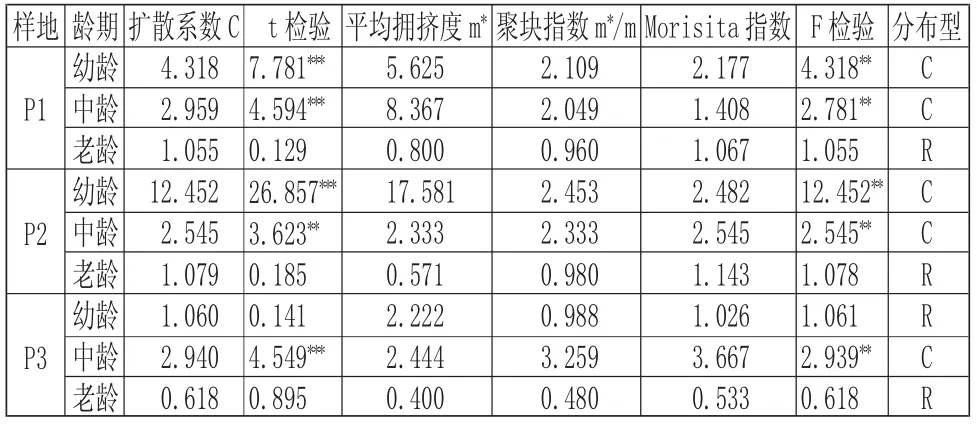
表8 不同發育階段海蓮種群分布格局的測定結果Tab.8 Result of Spatial Distribution Patterns of Bruguiera Sexangula Population at Different Age Stages
4 結論與討論
4.1 結論
海南清瀾港紅樹林林區內的海蓮種群各級樹高的個體數分布較均勻,但不同樣地間的各級高度個體數分布仍有一定的差異;海蓮最大冠幅直徑的個體是7m,66%個體的冠幅直徑大小在0.5m~3m 范圍內;63%的海蓮個體的胸徑大小在2.5cm~7.5cm 范圍內,種群整體的自然更新能力較強。運用5 種研究模型對海蓮胸徑與樹高和冠幅直徑之間的關系進行擬合,均達到了極顯著水平;目前海蓮種群都基本屬于聚集型分布,不同發育階段種群的分布格局大體上表現為集群型向隨機型轉變;但人為活動干擾對海蓮種群幼齡樹的分布格局影響較大。
4.2 討論
通過比較不同環境的海蓮種群樹高、胸徑和冠幅直徑結構特征,可以反映不同環境和立地條件對海蓮種群的作用強度和種群適應特征,不同樣地的海蓮種群結構的差異很可能與局部氣候、地形、土壤條件等區域環境特點有相關關系,反映出生境差異對種群結構的影響[16]。
海蓮種群的樹高與胸徑的關系及冠幅直徑與胸徑的關系用不同的數學模型方程來擬合,均呈極顯著相關關系,這與其他一些報道中的結果一致[13]。該文通過對海蓮種群的樹高、冠幅直徑與胸徑生長因子之間的相互作用關系分析,為進一步研究海南省文昌市清瀾港紅樹林保護區海蓮林種群結構特征、生長發育規律和自然演替規律等提供可靠的參考依據。
通過對胸徑和冠幅直徑的研究,不僅可以利用胸徑和冠幅直徑的相關方程來進行預測,在給定胸徑的某一林分的人工造林密度和郁閉度時間,而且也可以在適當的重疊情況下,充分的利用林分空間,使整個林分獲得最高的生產量,從而達到速生、優質、豐產的目的[12]。對紅樹主要樹種的樹高、胸徑(或地徑)的相關關系進行研究,也同樣可以為進一步研究人工引種,或自然的紅樹林生長發育規律、群落特征規律等提供可靠參考依據。同時,冠幅直徑的大小與胸徑的大小均能體現出植物個體在種群或群落中的競爭力,二者存在顯著的相關關系[17]。該研究中海蓮種群的胸徑和冠幅直徑呈現較高的正相關,而個體數量隨胸徑和冠幅直徑級的增加而逐漸減少,呈現較高的負相關,說明海蓮種群胸徑和冠幅直徑越大,種間競爭越強烈,進而使植物個體生長速率下降,甚至出現死亡,這個過程體現了海蓮種群的自疏過程。
在紅樹林群落中,由于其具有胎生繁殖的特性,成熟后的胚軸從母體上掉落,散落在母株周圍的泥土中,然后由胚軸發芽長成幼苗。海蓮的胚軸較長且林下有很多氣生根,盡管潮水漲落,也很難將掉落的胚軸帶到其他地方,因而容易形成其種群的集群分布格局。一旦幼苗長成成年個體,其樹冠的高郁閉度很難讓其他種類入侵成活,因此這種既定的分布格局比較難以改變。通過分析表明,清瀾港的紅樹林優勢海蓮種群均趨于集群分布(表7),說明海南清瀾港的整體環境很適宜海蓮種群的繁衍生長。
許多研究表明,從植物種群在不同發育階段的空間格局變化規律,可以推測成年個體的死亡、新生個體的產生以及種群遭受人為干擾的概況[18-19]。一般情況下,植物以母株為中心的繁殖方式是使低齡樹呈現聚集分布的原因;隨著樹齡的增長,植物對營養條件、光因子的需求會加大,以及在自疏和它疏等生物作用下,種群會趨于均勻分布;隨著樹齡的繼續增加,種群的分布格局類型也將趨于隨機分布[20-21]。該研究中由于海蓮種群的胎生現象使海蓮幼苗成片生長在母樹的周圍,而且因為海蓮種內幼苗之間的競爭比較小,容易形成集聚格局。在海蓮種群的繼續發育的過程中,個體對環境條件的要求會加劇,這便導致種內、種間的競爭加強,在競爭過程中,海蓮種群會因為自疏和它疏的作用,使種群密度劇烈下降,其結果勢必會使同一集群內的個體間出現分化,從而導致海蓮的種群密度顯著下降,集群將會解體,擴散趨勢將會越來越明顯;當這種變化持續到中樹階段,集群強度明顯比小樹的集群強度低,甚至在部分樣地出現了隨機分布現象。處于幼齡階段的海蓮種群呈現高程度的集群現象,有利于其存活和發揮群體效應;而成年后由于個體增大,集群的強度降低有利于個體獲得更多的環境資源。由此可知,海蓮種群的集群強度從幼齡到成年的變化規律,是種群的一種生存策略或者適應機制。該研究結果(表8)進一步表明,在自然條件下,海蓮種群的幼苗,幼樹呈現較高的集群分布,中齡樹也是集群分布,但集群分布明顯下降;而老齡樹為隨機分布;但在人為活動的干擾下,海蓮種群幼齡樹的分布格局較容易受到影響,因此未來要特別關注人為活動的類型和強度對清瀾港海蓮幼齡種群分布格局的影響。
猜你喜歡
百科探秘·海底世界(2025年4期)2025-05-07 00:00:00
歌海(2024年6期)2024-03-18 00:00:00
兒童故事畫報·自然探秘(2022年3期)2022-04-27 00:17:10
湖南包裝(2022年6期)2022-02-11 09:01:18
幼兒園(2021年4期)2021-07-28 07:38:04
發明與創新·小學生(2020年7期)2020-07-18 16:13:39
瘋狂英語·新讀寫(2020年4期)2020-06-03 07:01:02
瘋狂英語·初中天地(2018年6期)2018-11-24 02:39:24
歌海(2018年4期)2018-05-14 12:46:15
少兒科學周刊·兒童版(2015年11期)2015-12-17 03:38:09
