腐食酪螨對阿維菌素的抗性相關基因鑒定與功能分析
2022-07-16 11:47:23曲紹軒駱昕徐平侯子強李輝平林金盛侯立娟蔣寧馬林
江蘇農業學報 2022年3期
曲紹軒 駱昕 徐平 侯子強 李輝平 林金盛 侯立娟 蔣寧 馬林



摘要: 為揭示腐食酪螨對阿維菌素的抗性形成機制,通過RNA-seq技術獲得亞致死劑量阿維菌素脅迫下腐食酪螨轉錄組序列,鑒定到138個差異表達的基因被注釋到解毒代謝中,包括4個谷胱甘肽 S 轉移酶基因( GST ),12個細胞色素P450基因(P450)和3個羧酸酯酶基因( CarE )。利用RNAi技術沉默差異表達的11個解毒代謝基因后,腐食酪螨對阿維菌素的敏感性發生顯著性變化,毒力指數在0.65~6.05。結果顯示, GST 家族中TPGST7、TPGST45和TPGST49以及P450基因TPCYP3A9、TPCYP3A13和TPCYP4C3均參與腐食酪螨對阿維菌素的抗性形成。此外,葡萄糖醛酸轉移酶( GLT )家族基因TPGLT28首次被證實參與腐食酪螨對阿維菌素的抗性形成。
關鍵字: 食用菌害螨; 抗性相關基因; RNAi; 抗藥性
中圖分類號: S436.46+2?? 文獻標識碼: A? ?文章編號: 1000-4440(2022)03-0617-08
Identification and functional analysis of resistance genes of Tyrophagus putrescentiae Schrank to abamectin
QU Shao-xuan, LUO Xin, XU Ping, HOU Zi-qiang, LI Hui-ping, LIN Jin-sheng, HOU Li-juan, JIANG Ning, MA Lin
(Institute of Vegetable Crops, Jiangsu Academy of Agricultural Sciences, Nanjing 210014, China)
Abstract: To reveal the farmation mechanism of the resistance of ?Tyrophagus putrescentiae ?to abamectin, the transcriptome sequence of ?Tyrophagus putrescentiae ?under a sublethal dose of abamectin was obtained by RNA-seq technology. A total of 138 differentially expressed genes were identified and annotated to detoxification metabolism, including four glutathione ?S ?transferase genes ( GST ), 12 cytochrome P450 genes (P450) and three carboxylesterase genes ( CarE ). After silencing the 11 differentially expressed genes by using RNAi technology, the susceptibility of the transcriptome sequence of ?Tyrophagus putrescentiae ?to abamectin changed significantly, and the virulence index ranged from 0.65 to 6.05. The results showed that TPGST7, TPGST45 and TPGST49 in ?GST ?family and P450 genes TPCYP3A9, TPCYP3A13 and TPCYP4C3 were involved in the formation of resistance of T. putrescentiae to abamectin. In addition, the glucuronosyltransferase (GLT) family gene TPGLT28 was first confirmed to be involved in the formation of abamectin resistance.
Key words: mushroom mite pest; resistance-related gene; RNAi; insecticide resistance
螨類害蟲個體微小,繁殖力和對環境的適應能力都極強,單一藥劑脅迫使螨類很快產生抗藥性,已無法達到理想的防治效果 [1-2] 。阿維菌素因其殺蟲殺螨活性強、高效低殘留等特點而被廣泛用于田間包括腐食酪螨在內的害螨防治,導致害螨對其抗藥性日益增加 [1,3-4] 。2020年全國農業有害生物抗藥性監測結果顯示,二斑葉螨(Tetranychus urticae Koch)在中國4省4縣市監測地區的種群對阿維菌素已產生了高水平抗性,抗性倍數在304~1 051 ,對蟲螨腈、腈吡螨酯也處于中等至高水平抗性 [5] 。由于存在高抗性問題,阿維菌素已不能再用于這些地區二斑葉螨的防治,這與害螨抗藥性形成快、增長幅度高緊密相關 [5-6] 。
基于現代科技的發展,轉錄組測序技術、RNAi技術越來越多地應用于昆蟲毒理學研究領域 [7] 。目前,大量研究結果證實害螨通過代謝解毒能力增強、靶標部位敏感性降低等途徑產生對阿維菌素的抗性,靶標抗性主要包括谷氨酸氯離子通道 (Glutamate-gated chloride channels , GluCl)、γ-氨基丁酸受體(Gamma-aminobutyric acid receptor,GABAR)和γ-氨基丁酸氯離子通路(GABACls)等,解毒酶家族包括多功能氧化酶( MFO )、谷胱甘肽 S 轉移酶( GST )和羧酸酯酶( CarE )等。此外利用基因芯片技術還發現內環裂解雙加氧酶( ID-ICD )、絲氨酸蛋白酶( SP )、組織蛋白酶L( L )和P-糖蛋白(P-GP)等與朱砂葉螨(T. cinnabarinus Boiduval)對阿維菌素的抗性形成相關 [6-9] 。
腐食酪螨(Tyrophagus putrescentiae Schrank)是世界性重要糧儲害螨之一,分布廣,食性雜。近年,隨著食用菌“設施化制種+生態出菇”模式的迅速發展,恒溫恒濕的發菌環境和穩定的寄主資源,使得腐食酪螨的發生頻次不斷上升,危害也越來越嚴重,成為食用菌栽培的最主要害螨之一 [10-11] 。該螨危害毛木耳、黑木耳、銀耳、真姬菇、金針菇等食用菌菌絲和子實體,還攜帶并傳播病原菌,短時間內可造成食用菌大幅減產甚至絕產 [12] 。腐食酪螨個體微小,體長在0.4~ 0.6 mm,可從栽培袋或出菇瓶的口端縫隙、出菇孔或接種處進入料中取食菌絲,噴灑的藥劑無法有效滅殺袋內或瓶內的螨蟲,且缺乏有效的防控措施,導致亂用和濫用殺蟲殺螨劑的現象頻發,嚴重制約著食用菌產業的綠色安全生產 [13] 。長期的化學防治,使其對阿維菌素、噠螨靈、有機磷類農藥產生了高抗藥性 ?[1-4] ,而有關腐食酪螨對阿維菌素的代謝、解毒及其抗性機制仍不清楚。
本研究利用高通量測序技術對腐食酪螨在阿維菌素脅迫下的轉錄組進行研究,挖掘和鑒定該螨對阿維菌素的抗性相關基因,并通過RNAi技術進一步明確與抗藥性密切相關的基因功能,為揭示腐食酪螨抗藥性的分子機制和制定治理策略奠定理論基礎。
1 材料與方法
1.1 試驗材料
1.1.1 供試螨蟲 供試腐食酪螨(T. putrescentiae)于2012年采自江蘇省豐縣師寨鎮毛木耳子實體上,轉至實驗室內,無接觸化學藥劑,人工氣候箱(25 ℃± 1 ℃,相對濕度80%± 5%)中以香菇(Lentinula edodes L.)麥粒種為飼料飼養至今。飼養方法同Qu等 [12] 。
1.1.2 主要試劑與儀器 90%阿維菌素原藥購自上海源葉生物科技有限公司,柱式法通用型總RNA提取試劑盒(9767)購自寶生物工程(大連)有限公司,一步法TB Green RT-PCR定量檢測試劑盒(RR096A)購自寶生物工程(大連)有限公司。主要儀器有SPX-2501人工氣候箱(上海博訊實業有限公司產品)、CFX95 Touch型實時熒光定量PCR儀(BIO-RAD公司產品)、PCR擴增儀(BIO-RAD公司產品)、核酸電泳儀和水平電泳槽(北京市六一儀器廠產品)、顯微凝膠成像儀(上海培清科技有限公司產品)、核酸定量儀(NanoDrop One, Thermo Fisher Scientific)。
1.2 試驗方法
1.2.1 亞致死劑量阿維菌素處理腐食酪螨 阿維菌素原藥(溶于丙酮)用ddH2 O稀釋成1 000 ?mg/L? 母液在4 ℃冰箱里避光儲存。使用ddH2 O將阿維菌素母液稀釋到0.5 ng/L ,采用胃毒觸殺聯合毒力測定法處理腐食酪螨,具體操作方法同駱昕等 [3] 方法。將0.025 g成螨(約1 500 頭)放入養蟲瓶內,蓋上瓶蓋,然后置于人工氣候箱(25 ℃、相對濕度75%± 5%、無光照)中飼喂24 h,然后用200目過濾網收集活螨,ddH2 O清洗3次,用濾紙吸干水分后放入1.5 ml的離心管中,液氮中預冷4 h以上,放入-80 ℃ 冰箱中保存,重復3次。對照組,直接取0.025 g飼養在香菇麥粒種中的成螨,ddH2 O清洗3次后,同處理組一樣的方式進行樣品預冷和保存。
1.2.2 轉錄組測序和生物學分析 螨蟲樣品委托華大基因有限公司(深圳)進行總 RNA 提取、質量分析、cDNA文庫的建立及測序工作。采用Illumina HiSeq 2500系統進行測序,將拼接的unigenes與NR(NCBI non- redundant protein sequences)、NT(NCBI nucleotide sequences)、Pfam(Protein family)、KOG/COG(Clusters of orthologous groups of proteins)、Swiss-prot(A manually annotated and reviewed protein sequence database)、KEGG (Kyoto encyclopedia ofgenes and genomes)、GO(Gene ontology)等數據庫進行基因功能注釋。使用DESeq2進行差異基因的篩選,采用FDR法對差異檢驗的P值作多重假設檢驗校正得到 Q 值,為了得到顯著差異表達的基因,篩選條件為差異倍數(Fold Change)≥2或≤ -2且 Q 值< 0.01,然后通過對差異基因進行GO功能注釋和KEGG通路富集分析 [4] 。
1.2.3 解毒代謝相關基因的鑒定 在轉錄組序列中搜索解毒代謝相關基因,以關鍵詞谷胱甘肽 S 轉移酶( GST )、細胞色素P450(P450)和羧酸酯酶( CarE )等進行搜索。篩選差異倍數大于4、FDR值< 0.01且被注釋在Drug metabolism-cytochrome P450(ko01100、ko00983或ko00480等)通路或Drug metabolism-other enzymes(ko01100、ko00983、ko1049、ko7383或k15743等)通路或metabolism of xenobiotics by cytochrome P450(ko00980或ko00982)通路。根據NR的注釋結果和KEGG通路的分析,過濾掉短片段和等位基因后,將擁有完整開放閱讀框的基因序列作為進一步研究基因功能的候選序列 [2,4,14-15] 。
1.2.4 候選的解毒代謝相關基因的功能分析 通過siRNA誘導的RNA干擾來驗證候選基因在腐食酪螨對阿維菌素抗性中的作用。siRNA序列利用在線工具(http://biodev.extra.cea.fr/DSIR/DSIR.html)設計,靶向序列為19個堿基,在RNA的3′端加dTdT垂懸(引物見表1) [14] 。siRNA合成由南京擎科生物科技有限公司完成,用Rnase-free水將siRNA引物稀釋成100 μmol/L (1.0 μg/μl ),保存于-80 ℃ 冰箱備用。
參照文獻[3]、[15]建立的飼喂法將siRNA導入腐食酪螨體內進行RNA沉默分析,這里我們將葉片換成香菇麥粒種,每個處理5粒麥粒種。60目過濾篩收集腐食酪螨成螨(3~ 5 d),然后用ddH2 O清洗4次后用250目紗網收集,吸干紗網上水分,轉置2.0 ml的離心管中饑餓24 h。將100頭成螨放入25 ml的養蟲瓶,分別用500 ng/μl 、200 ng/μl 、50 ng/μl 和25 ng/μl 的siRNA進行基因沉默,帶上蓋子飼喂24 h后,采集螨蟲樣品用于RNA提取,隨后RT-qPCR分析候選的解毒酶基因表達水平,分析最佳的siRNA飼喂劑量。重復 3 次,以ddH2 O處理的麥粒種飼喂的螨蟲為對照。以腐食酪螨 β-tublin 基因為內參(引物見表1),一步法RT- qPCR反應條件為:逆轉錄42 ℃溫育5 min,95 ℃ 10 s終止反應;PCR擴增:95 ℃預溫育5 s后,60 ℃ 20 s,循環40次,熔解曲線65~ 95 ℃,每15 s升溫0.3 ℃。采用 2 -△△ Ct? 方法計算基因的相對表達量 [15] 。
阿維菌素母液用ddH2 O稀釋成20.0 mg/L? 、10.0 mg/L? 、2.0 mg/L? 、1.0 mg/L? 和0.5 mg/L? ?5個質量濃度梯度備用,采用胃毒觸殺聯合毒力測定法處理腐食酪螨,具體操作方法同駱昕等 [3] 方法。處理組根據上述試驗確定的最佳劑量siRNA分別飼喂腐食酪螨成螨24 h后,將100頭轉到含不同質量濃度阿維菌素藥劑的養蟲瓶內,24 h后觀察腐食酪螨的死亡數量,每個siRNA處理重復3次,對照組為清水處理 [15] 。
1.3 數據分析
以上測得的數據采用Excel 2010 進行整理。單因素方差分析( one-way ANOVA) 和Student-Newman-Keuls (SNK) 檢驗后進行多重比較( P = 0.05),使用SPSS 21.0軟件完成。毒力測定所得數據用Excel軟件進行相關分析和回歸分析,得出相關系數、毒力回歸方程( Y =a x +b)、 LC 50 值及其95%置信區間。
2 結果與分析
2.1 腐食酪螨的轉錄組測序結果
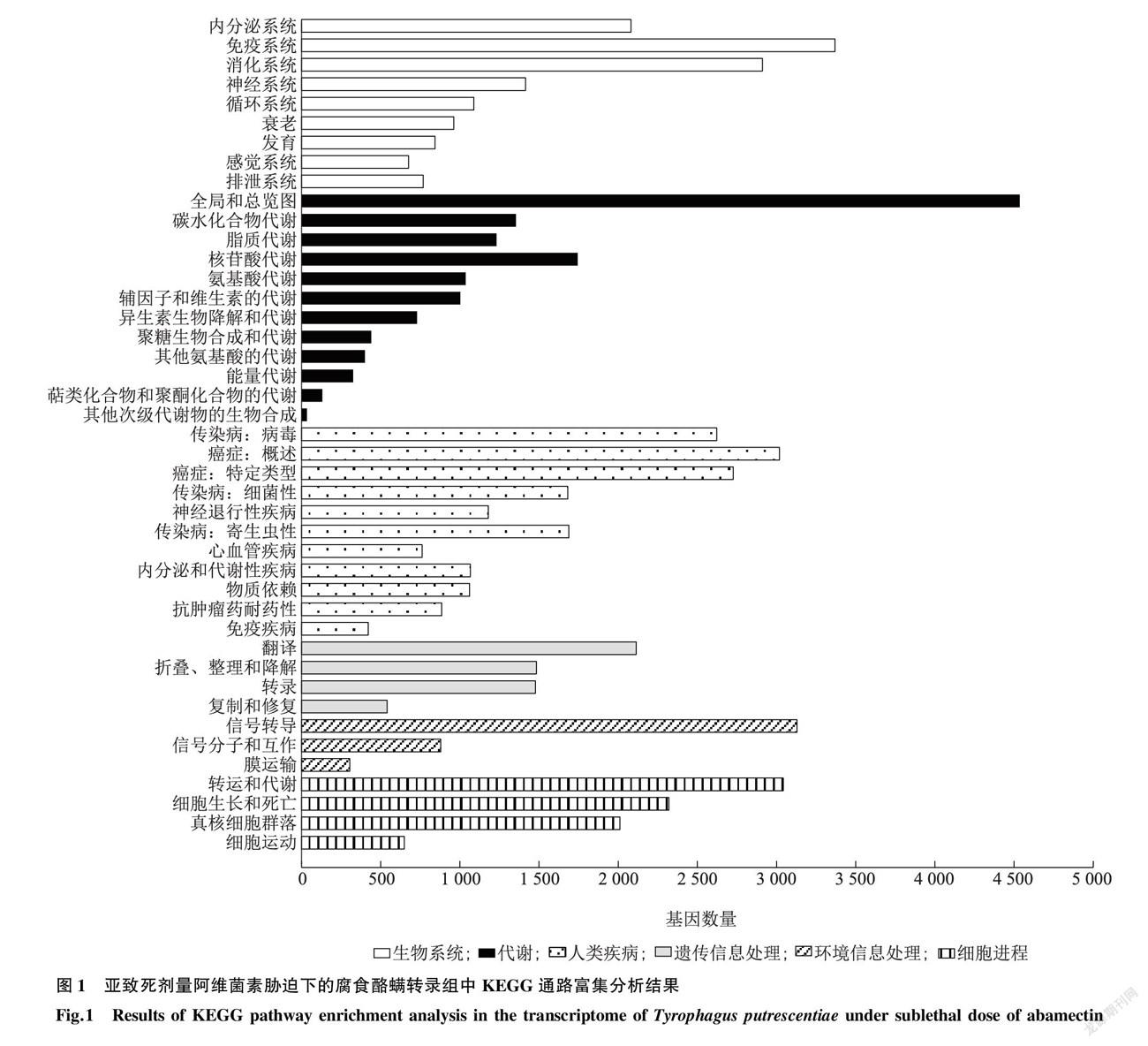
經測序質量控制,6個樣品共得到37.25 Gb數據,組裝并去掉冗余后得到62 268 個Unigene,總度、平均度、50% contig總長度(N50)以及G+C含量分別為82 154 640 ?bp、1 319 ?bp、2 360 bp和48.36%。然后將Unigene比對到七大功能數據庫進行注釋,最終分別有37 332 (NR:59.95%)、22 143 (NT:35.56%)、29 589 (SwissProt:47.52%)、28 621 (KOG:45.96%)、29 636 (KEGG:47.59%)、13 224 (GO:21.24%)以及33 530 (Pfam:53.85%)個Unigene獲得功能注釋。使用Transdecoder檢測出38 791 個蛋白質編碼序列(Coding sequence,CDS)。使用DESeq2進行對照組與處理組間的差異表達分析,將 ?Fold Change ≥2 或≤-2且 FDR < 0.01 作為篩選標準,統計分析所得的差異基因數量,共鑒定出差異基因14 435 個,其中上調表達1 188 個,下調表達13 247 個。進一步對差異表達基因進行聚類分析,KEGG Pathway富集分析結果顯示,亞致死劑量阿維菌素處理腐食酪螨后,差異基因在信號傳導、免疫系統、內分泌系統、藥物代謝和傳染性疾病相關通路富集(圖1)。GO 富集分析發現受阿維菌素脅迫后腐食酪螨在涉及結合、催化活性、細胞過程、新陳代謝過程、ATP結合、解旋酶活性等生物過程、細胞組分和分子功能組中大量富集(圖2),有138個差異表達的基因被注釋到解毒代謝中。
2.2 腐食酪螨轉錄組中解毒代謝相關基因分析
通過比對GO、KEGG和KOG數據庫注釋的序列,在亞致死劑量阿維菌素脅迫下腐食酪螨轉錄組中發現73個谷胱甘肽 S 轉移酶( GST )unigene、296個細胞色素P450(P450)unigene和97個羧酸酯酶( CarE )unigene等解毒代謝相關基因。過濾掉短片段、等位基因后,共鑒定到30個 GST ?unigene,全部與Drug metabolism-cytochrome P450通路、metabolism of xenobiotics by cytochrome P450通路有關且具有完整的ORF;鑒定到114個P450 unigene,其中22個P450 unigene與Drug metabolism-cytochrome P450通路相關,18個P450 unigene與Drug metabolism-other enzymes通路相關,24個P450 unigene與metabolism of xenobiotics by cytochrome P450通路相關。根據KOG注釋結果,對P450 unigene進行分類,其中46個unigene屬于Cytochrome P450 CYP3/CYP5/CYP6/CYP9 subfamilies(KOG0158),13個unigene屬于Cytochrome P450 CYP4/CYP19/CYP26 subfamilies(KOG0157);鑒定到68個 CarE ?unigene,其中6個 CarE ?unigene與Drug metabolism-other enzymes通路有關。
解毒代謝相關基因中鑒定到的P450基因數和羧酸酯酶基因數分別占3種解毒酶基因的 53.8%和32.1%,遠多于谷胱甘肽 S -轉移酶基因的數量,說明這2類解毒酶在腐食酪螨解毒代謝過程中有著極其重要的地位。
2.3 候選的解毒代謝相關基因的功能分析
從上述鑒定到的解毒代謝相關基因中篩選差異倍數大于4、FDR<0.01且被注釋在Drug metabolism-cytochrome P450通路或Drug metabolism-other enzymes通路或metabolism of xenobiotics by cytochrome P450的基因作為候選基因(表1),用于進一步研究其在腐食酪螨對阿維菌素抗性的作用。
腐食酪螨對阿維菌素抗性相關的11個基因包括4個 GST 家族基因(TPGST7、TPGST8、TPGST45和TPGST49)、4個P450家族基因(TPCYP3A9、TPCYP3A13、TPCYP3A18和TPCYP4C3)、1個羧酸酯酶基因(TPCarE6),2個葡萄糖醛酸轉移酶( GLT )基因TPGLT16和TPGLT28,分別用siRNA引物干擾24 h后,進行沉默效率測定。
RT-qPCR結果表明,不同siRNA劑量飼喂腐食酪螨成螨均使靶標基因的表達量降低(圖3),500 nmol/L ?siRNA可最高降低靶標基因表達量的74.6%,沉默效率在51.7%~ 74.6%,200 nmol/L 劑量對靶標基因的沉默效率在40.6%~ 58.2%,50 nmol/L 和25 nmol/L 劑量對靶標基因的沉默效率較低(19.2%~ 40.3%)。因此,選擇500 nmol/L 劑量對靶標基因進行沉默。
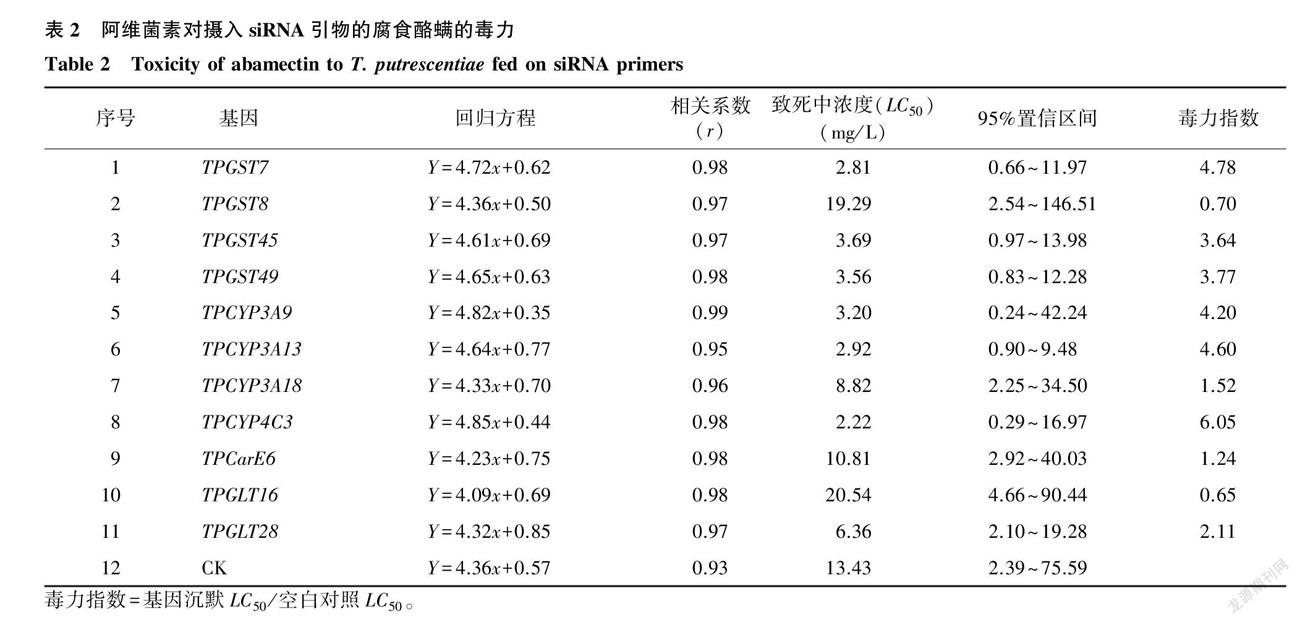
RNAi技術沉默差異表達的11個解毒代謝相關基因后,腐食酪螨對阿維菌素的敏感性也發生顯著性變化(表2)。 GST 家族基因TPGST7、TPGST8、TPGST45和TPGST49沉默后,阿維菌素對腐食酪螨的致死中濃度(LC50 )從未干擾時的13.43 mg/L 最低下降到2.81 mg/L ,毒力指數分別為4.78、0.70、3.64和3.77;P450家族基因TPCYP3A9、TPCYP3A13、TPCYP3A18和TPCYP4C3沉默后,阿維菌素對腐食酪螨的LC50 最低下降到2.22 mg/L ,毒力指數分別為4.20、4.60、1.52和6.05;羧酸酯酶基因TPCarE6沉默后的毒力指數為1.24;2個 GLT 家族基因TPGLT16和TPGLT28沉默后的毒力指數分別為0.56和2.11。結果表明,沉默 GST 家族基因TPGST7、TPGST45和TPGST49,P450家族基因TPCYP3A9、TPCYP3A13和TPCYP4C3,以及 GLT 家族基因TPGLT28后,腐食酪螨對阿維菌素的敏感性提高,說明這些基因很可能參與了腐食酪螨對阿維菌素抗性的形成。
3 討 論
阿維菌素是一種具有良好活性的殺蟲殺螨劑,被廣泛用于農業害蟲、害螨以及衛生害蟲的防治 [16-17] ,對其抗性機制研究是殺蟲劑毒理學的熱點之一 [7] 。目前,已證實P450家族中的CYP6、CYP9、CYP4基因是蚊蟲產生阿維菌素抗性的重要因素 [18] ; ?GST 、P450、 CarE 等參與農業害蟲對阿維菌素的代謝抗性 [6-8] ;果蠅和二斑葉螨的P-GP蛋白通過誘導幾丁質合成酶降低阿維菌素的穿透性而增強抗性 [9] ;在害螨中,位于二斑葉螨中腸上層細胞的CYP392A16是阿維菌素抗性形成的主效基因,該基因也同樣介導果蠅對阿維菌素的抗藥性 [19] 。
有關腐食酪螨的抗藥性研究較少,僅報道了腐食酪螨對磷化氫及多種常用的殺蟲劑產生抗性,但其抗藥性機理研究甚少 [1, 20] 。本研究對亞致死劑量阿維菌素脅迫下的腐食酪螨轉錄組進行了測序,獲得了解毒代謝相關基因表達模式,鑒定到30個 GST 、114個P450和68個 CarE 基因,發現被注釋到解讀代謝通路上的4個 GST 、12個P450和3個 CarE 基因差異表達。利用siRNA介導的RNA干擾技術分析了沉默差異表達基因對阿維菌素毒力的影響,結果證實了 GST 家族中的TPGST7、TPGST45和TPGST49和P450家族中的TPCYP3A9、TPCYP3A13和TPCYP4C3在腐食酪螨對阿維菌素的抗性形成中起重要作用。其中,P450家族中的3個基因屬于CYP3和CYP4基因家族。大量研究結果證明GST和P450家族基因參與殺蟲劑的解毒代謝,這些基因的過表達是害蟲和害螨產生對阿維菌素、吡蟲啉、噻蟲嗪和毒死蜱等不同殺蟲劑交互抗性的重要因素 [21-22] 。此外,沉默葡萄糖醛酸轉移酶( GLT )家族基因TPGLT28后,腐食酪螨對阿維菌素的敏感性提高,毒力指數為2.11,被證實也參與了腐食酪螨對阿維菌素抗性的形成,但這類家族基因的抗性作用機制目前尚未報道。本研究通過轉錄組和RNAi分析挖掘出參與腐食酪螨對阿維菌素抗性形成的重要基因,為今后解析腐食酪螨對阿維菌素抗藥性機理提供了依據。
參考文獻:
[1] 周 紅,丁 偉. 螨類化學控制存在的問題及其對策[J]. 植物醫生, 2020, 33(1):27-32.
[2] 周興隆,楊順義,李靜靜,等. 二斑葉螨多重抗性品系的選育及其解毒酶活力分析[J]. 西北農林科技大學學報(自然科學版), 2017, 45(6): 134-140.
[3] 駱 昕,曲紹軒,馬 林,等. 六種殺螨劑對羅賓根螨和腐食酪螨的室內毒力測定[J]. 食用菌學報, 2018, 25(2):132-136.
[4] XU Z F, ZHU W Y, LIU Y C, et al. Analysis of insecticide resistance-related genes of the Carmine spider mite ?Tetranychus cinnabarinus ?based on a de novo assembled transcriptome[J]. PLoS One, 2014,9(5):e94779.
[5] 張 帥. 2020年全國農業有害生物抗藥性監測結果及科學用藥建議[J]. 中國植保導刊, 2021, 41(2): 71-78.
[6] 沈一凡,沈慧敏,岳秀利,等. 二斑葉螨抗阿維菌素品系選育及其解毒酶系活力變化[J].植物保護, 2014, 40(5): 44-48,74.
[7] 伍一軍. 近二十年我國殺蟲劑毒理學研究進展——昆蟲對殺蟲劑的抗性研究[J]. 應用昆蟲學報, 2020, 57(5): 995-1008.
[8] KWON D H, YOON K S, CLARK J M, et al. A point mutation in aglutamate-gated chloride channel confers abamectin resistance in the two-spotted spider mite, ?Tetranychus urticae ?Koch[J]. Insect Molecular Biology, 2010, 19(4): 583-591.
[9] XU Z F, SHI L, PENG J F, et al. Analysis of the relationship between P-glycoprotein and abamectin resistance in ?Tetranychus cinnabarinus ?(Boisduval)[J]. Pesticide Biochemistry and Physiology, 2016, 129: 75-82.
[10] JEONG Y K, LEE H, LEE S J, et al. Molecular cloning and the allergenic characterization of tropomyosin from ?Tyrophagus putrescentiae [J]. Protein and Peptide Letters, 2007, 14: 431-436.
[11] KHERADMAND K, KAMALI K, FATHIPOUR Y, et al. Development, life table and thermal requirement of ?Tyrophagus putrescentiae ?(Astigmata: Acaridae) on mushrooms[J]. Journal of Stored Products Research, 2007, 43: 276-281.
[12] QU S X, LI H P, MA L, et al. Effects of different edible mushroom hosts on the development, reproduction and bacterial community of ?Tyrophagus putrescentiae ?(Schrank) [J]. Journal of Stored Products Research, 2015, 61: 70-75.
[13] QU S X, WANG X F, LI H P, et al. A Gustatory receptor used for rapid detection of ?Tyrophagus putrescentiae ?in fungi hosts[J]. Scientific Reports, 2018, 8(1): 11425.
[14] KENNERDELL J R, CARTHEW R W. Use of ds RNA-mediated genetic interference todemonstrate that frizzled and frizzled 2 act in the wingless pathway[J]. Cell, 1998, 95 (7): 1017-1026.
[15] 敖已倩云,申光茂,王夢瑤,等. 朱砂葉螨 β-COP和Sro 基因鑒定及其沉默致死效果[J].中國農業科學, 2017 ,50(18): 3529-3539.
[16] 林海蔚,韓慧霖,羅 茵,等. 阿維菌素B2a衍生物類皮雷菌素對草地貪夜蛾的室內毒力[J].南方農業學報,2020,51(6):1287-1292.
[17] 姚晶晶,陳麗君,王明銳,等. 西瓜中殺線蟲劑農藥殘留及風險評估[J].江蘇農業科學,2020,48(6):168-172.
[18] 彭 薈,劉宏美. 細胞色素P450介導蚊蟲代謝抗性及防治應用的相關研究進展[J].中國熱帶醫學, 2022, 22(1): 79-83.
[19] PAPAPOSTOLOU K M, RIGA M, SAMANTSIDIS G R, et al. Over-expression in cis of the midgut P450 CYP392A16 contributes to abamectin resistance in ?Tetranychus urticae [J]. Insect Biochemistry and Molecular Biology, 2022, 142: 103709.
[20] 剪福記. 儲藏物螨類熏蒸防治方法研究狀況及進展[J]. 糧食儲藏, 1996, 25(3): 3-10.
[21] 朱 江,邱星輝. 昆蟲抗藥性相關細胞色素P450基因的表達調控機制[J]. 昆蟲學報, 2021, 64(1): 109-120.
[22] SHEN J, LI Z, LI D, et al. Biochemical mechanisms, cross-resistance and stability of resistance to Metaflumizone in ?Plutella xylostella [J]. Insects, 2020, 11(5): 311.
(責任編輯:張震林)
收稿日期:2021-11-16
基金項目:財政部和農業農村部國家現代農業產業技術體系項目(CARS-20);國家自然科學青年基金項目(32001911);江蘇省農業科技自主創新基金項目[CX(21)2021]
作者簡介:曲紹軒(1982-),女,遼寧大連人,博士,副研究員,主要從事食用菌有害生物防治研究。(Tel)025-84390159;(E-mail)qusx@jaas.ac.cn
通訊作者:馬 林,(E-mail)malin1590@sina.com
