布氏輪藻肌動蛋白基因家族的全基因組鑒定及表達分析
2022-07-22 06:53:16胡莎莎張冬群張苡芃韓鵬舉蘭利瓊卿人韋
四川大學學報(自然科學版) 2022年4期
關鍵詞:分析
胡莎莎,張冬群,張苡芃,韓鵬舉,蘭利瓊,卿人韋
(四川大學生命科學學院生物資源與生態環境教育部重點實驗室,成都 610065)
1 引 言
近年來關于基因家族的研究主要集中于轉錄因子家族、酶家族(激酶、代謝通路相關酶類),如WRKY基因家族、MYB轉錄因子家族等,關于Actin基因家族的分類、功能分析研究較少.Actin基因家族是一類超基因家族,除了包含典型的Actin蛋白外,還含有ALP同源蛋白結構域,它們具有稱為“actin fold”的共同結構特征,這些蛋白由一個多基因家族編碼共同組成了Actin 基因家族[1].該基因家族在動植物中廣泛存在,與各種微絲結合蛋白共同作用,形成獨特的微絲骨架結構,參與成膜體結構的組成、囊泡運輸、細胞運動、物質轉運、脅迫響應等各項生命活動[2].與動物相比,植物肌動蛋白之間的序列差異更大,表明肌動蛋白基因家族可能在植物中具有更加復雜的生物學功能[3,4].
輪藻植物具有成膜體結構,且微絲、微管等骨架蛋白參與了該結構的組成[5].另外,在輪藻植物中還廣泛存在無絲分裂現象,且微絲結構的破壞顯著抑制了細胞的無絲分裂過程[6].但目前為止,還未有關于輪藻植物Actin基因家族的鑒定、結構分析等相關研究,因此本研究基于布氏輪藻全基因組對Actin基因家族進行了鑒定分析,從基因序列特征、蛋白理化性質、結構特征、系統發育、啟動子順式作用元件及組織表達等方面對布氏輪藻的Actin基因家族進行系統的研究,以揭示Actin基因家族在輪藻中的基因結構、蛋白特征及可能參與的功能,這一結果也將為進一步探究布氏輪藻的Actin基因家族的具體的生理生化功能及其特殊結構形成的具體分子機制奠定基礎.同時,可以為探究輪藻植物無絲分裂現象的基因調控機制提供一定的理論依據.
2 材料與方法
2.1 材 料
布氏輪藻的基因組文件和注釋文件下載于Phyzome基因組數據庫(https://phycocosm.jgi.doe.gov/Chabra1/Chabra1.home.html),擬南芥Actin基因家族下載于擬南芥基因組數據庫TAIR(https://www.arabidopsis.org/browse/genefamily/index.jsp).
2.2 方 法
2.2.1 成員的鑒定和理化性質分析 利用TBtools根據布氏輪藻基因組文件和注釋文件提取基因的CDS序列,并將CDS序列文件轉化為蛋白序列文件,去除重復后獲得非冗余的蛋白序列文件.基因家族的鑒定通常包括兩種方法,第一種方法利用Blast軟件通過序列相似性進行序列比對的方法篩選,第二種利用Pfam數據庫提供的蛋白結構域模型文件摸索未知序列[7],如果具有相同的結構域則認為該基因屬于同一家族成員.為了獲得更完整的布氏輪藻Actin基因家族成員,本研究結合了兩種方法對布氏輪藻的Actin基因家族進行了鑒定.首先,以擬南芥Actin基因家族蛋白序列作為參考與布氏輪藻的蛋白序列進行本地Blast比對(E-value<10-10), 獲得布氏輪藻中Actin基因的潛在序列.在Pfam數據庫中下載Actin結構域的隱馬爾可夫模型(PF00022),使用HMMER軟件對布氏輪藻蛋白序列進行檢索(E-value<10-10),篩選包含Actin結構域的潛在基因家族.對blast結果與HMMER結果取并集,將獲得的所有蛋白序列提交至NCBI的CD-search網站最終獲得包含完整Actin結構域的基因.根據基因在scaffold上的順序,將其分別重命名為CbACT1~CbACT16.使用ExPASy在線網站獲得Actin基因的等電點、分子質量和序列長度等信息,并預測其不穩定系數和平均親水系數.通過SOPMA網站預測分析蛋白序列的二級結構,并使用Plant-mPLoc進行蛋白亞細胞定位分析.
2.2.2 系統發育及基因結構分析 使用MAFFT軟件對獲得的蛋白序列進行比對,比對策略選擇auto其他參數選擇默認[8].將比對結果導入Phylosuite軟件[9],使用Trim AI對序列進行修剪,并將其轉換為PHY格式,最后通過IQ-TREE進行建樹分析, Bootstrap設置為1000[10],IQTREE通過自動檢測氨基酸替換模型建立最大似然樹.基因結構的展示根據布氏輪藻的GFF注釋文件,利用TBtools軟件的Gene Structure View程序進行分析.
2.2.3 蛋白保守基序分析和結構域分析 通過MEME在線預測蛋白的保守基序,motif設置為10,其他保持默認選項,利用TBtools軟件繪制蛋白保守基序圖.利用NCBI的CD-search工具對16條Actin基因進行保守結構域分析,提取結果文件中的結構域信息,將其導入TBtools對蛋白序列的結構域可視化.
2.2.4 染色體定位及共線性分析 使用BLAST對輪藻所有蛋白序列進行雙向比對并將雙向Blast結果進行合并(E<10-5).利用TBtools結合Excel對GFF文件進行簡化處理,將簡化的GFF文件及Blast結果文件作為輸入文件,通過MCScanX軟件進行共線性分析,最后利用TBtools將結果進行可視化[11].
2.2.5 啟動子區域順勢作用元件分析 使用TBtools從布氏輪藻的基因組序列文件中提取Actin基因家族成員起始密碼子上游2000 bp的DNA序列作為Actin基因的啟動子區域,將其提交至PlantCARE網站預測Actin所包含的順式作用元件.將結果導入到Excel中整理,最后用TBtools軟件可視化啟動子元件.
2.2.6 轉錄表達模式分析 根據相關文獻下載布氏輪藻基因組數據庫中的表達矩陣,檢索16個Actin基因在4個不同組織包括營養組織、藏精器、藏卵器及合子中的表達量,將表達量值經過log2(FPKM+1)轉換后利用TBtools軟件的HeatMap繪制基因表達熱圖.
3 結果與分析
3.1 Actin基因家族的鑒定和蛋白理化性質分析
通過序列比對和結構域篩選,共獲得布氏輪藻16個Actin基因,根據基因所在Scaffold的排列順序將所有基因重命名為CbACT1~CbACT16.理化性質分析結果表明Actin基因氨基酸數量在361(CbACT9)~1182(CbACT7)AA之間,分子量在39 886.71(CbACT14)~117 256.72(CbACT13)Da之間,理論等電點在4.68(CbACT12)~8.93(CbACT11)之間,只有三個為堿性蛋白質(理論等電點>7),其余均為酸性蛋白質(理論等電點>7).氨基酸的平均親水系數表明,除了 2個基因(CbACT1、CbACT15)為疏水性蛋白外(平均親水系數>0),其余均為親水性蛋白(平均親水系數<0).不穩定指數表明有7個蛋白(CbACT2、CbACT5、CbACT7、CbACT11、CbACT13、CbACT15、CbACT116)為不穩定蛋白(不穩定指數>40),其余Actin基因均為穩定蛋白(不穩定指數<40)(表1).

表1 布氏輪藻的肌動蛋白基因家族基本特征
3.2 二級結構預測及亞細胞定位分析
亞細胞定位預測顯示除了CbACT13定位于葉綠體之外,其余均定位于細胞質(表2).為了進一步了解Actin基因家族的空間結構特征,對其進行了二級結構分析,結果表明,16條Actin基因均含有α螺旋、無規則卷曲、延伸鏈和β折疊.其中,α螺旋和無規則卷曲占二級結構總量的26.48%(CbACT2)~46.51%(CbACT2),是該基因家族的主要二級結構元件;延伸鏈占比12.30%(CbACT15)~22.22%(CbACT1),β轉角所占比例最少,在6.27%(CbACT3)~11.25%(CbACT16)之間.
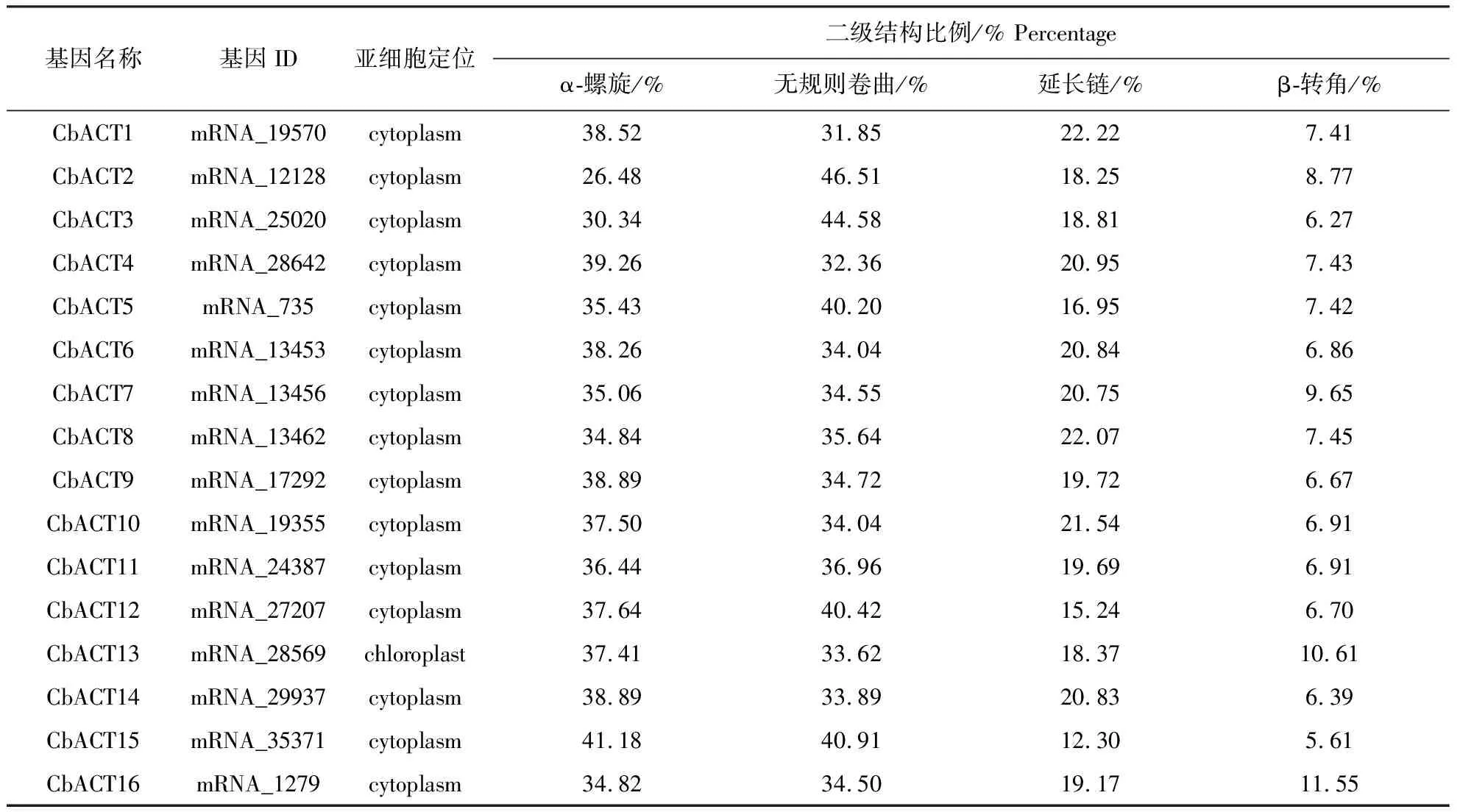
表2 布氏輪藻的肌動蛋白亞細胞定位及二級結構
3.3 系統發育和基因結構分析
為了研究布氏輪藻的Actin基因家族的系統發育關系和基因結構特征,本研究對布氏輪藻的16條Actin基因的蛋白序列進行了序列比對建樹分析和基因結構分析(圖1).系統發育分析結果表明布氏輪藻的Actin基因可以分為兩個亞家族,結合Actin基因的結構分析,發現同一亞家族的基因
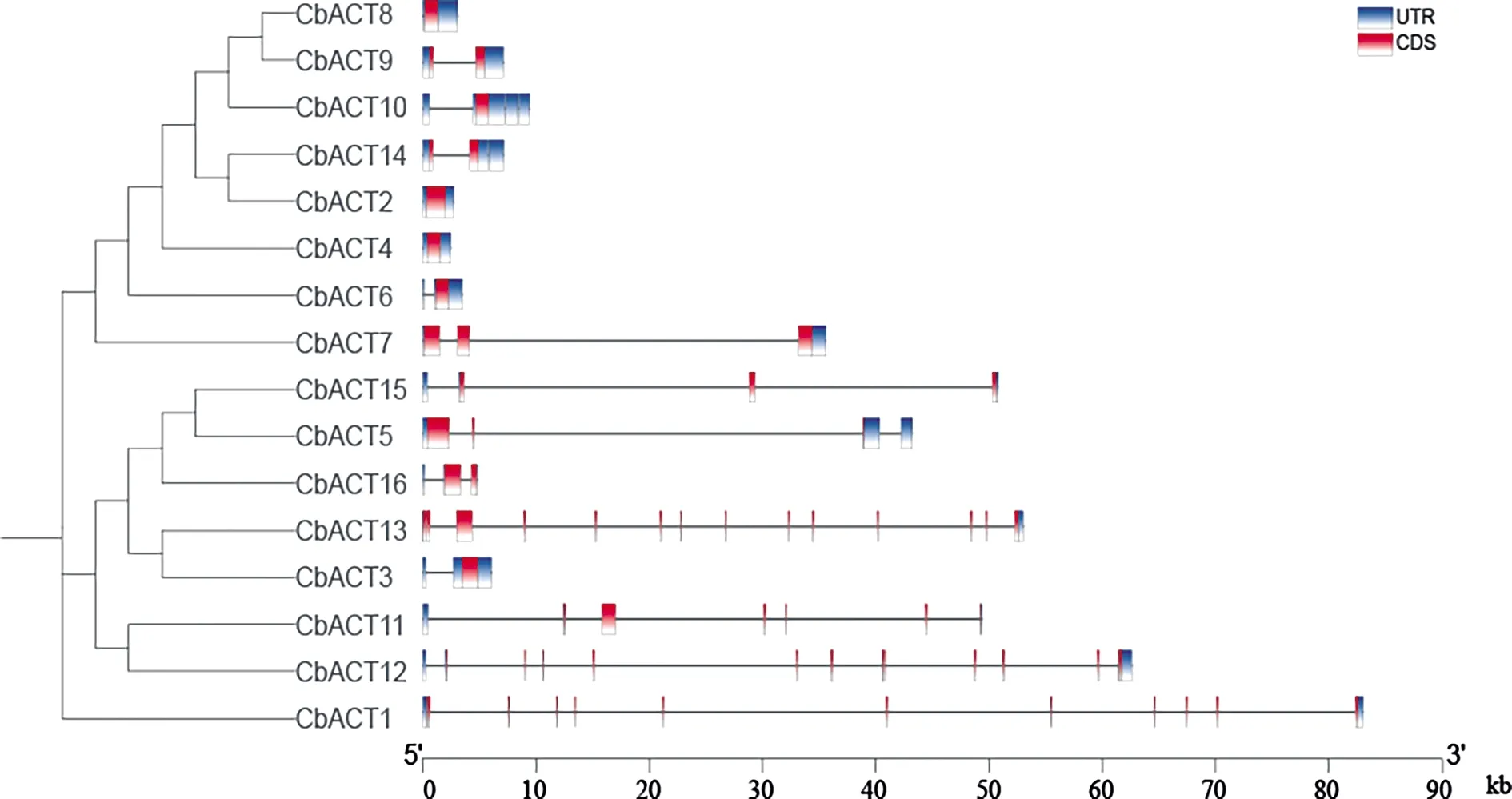
圖1 CbACT蛋白進化樹及基因結構
結構較為相似,類型I中基因結構較為保守,外顯子數目差異較小,基因序列外顯子數目均小于4.而類型Ⅱ發生了不同程度的內含子插入導致基因結構變化較大,外顯子數目變化范圍在1~14之間,這些結構差異的基因進而通過不同的轉錄加工,參與不同的轉錄調控過程.
3.4 結構域及保守基序分析
利用NCBI-CDsearch對布氏輪藻的Actin基因進行結構域分析(圖2),所有的Actin基因家族均含有Actin的典型結構域,除此之外類型Ⅰ的CbACT7還包含有RT-like結構域,類型Ⅱ有2條序列還包含有F-box-like、PRK-13357的結構域,因此在輪藻中Actin功能還參與了更加復雜的生命活動.為了進一步了解序列的保守特征,利用MEME在線工具對該基因家族的保守基序進行了預測分析,結果顯示:motif7,motif3,motif6在所有序列中都存在,屬于高度保守序列,可能作為Actin基因家族保守結構域的關鍵序列而存在.基序在不同家族成員中的分布狀態均存在一定的差異,進化關系越相近的成員共有的保守基序越相似,如類型Ⅰ蛋白序列均含有基序1、2、3、4、6、9.因此這些相同的保守基序表明同一亞家族的成員可能具有相似的功能,不同的保守基序可能是不同Actin執行不同功能所必須.
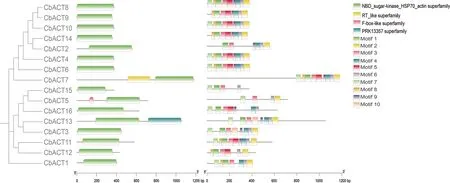
圖2 CbACT結構域及保守基序
3.5 染色體定位及共線性分析
Actin基因家族的染色體定位結果表明除了CbACT6、CbACT7、CbACT8分布于同一條scaffold上,其余單獨分布于13條scaffold上(圖3).為了研究Actin基因家族的擴增機制,利用TBtools對布氏輪藻Actin基因家族進行物種內共線性分析,結果表明只有一對Actin基因為片段復制所產生,沒有發現串聯重復的基因,推測該植物的Actin基因家族成員少部分是由基因片段復制
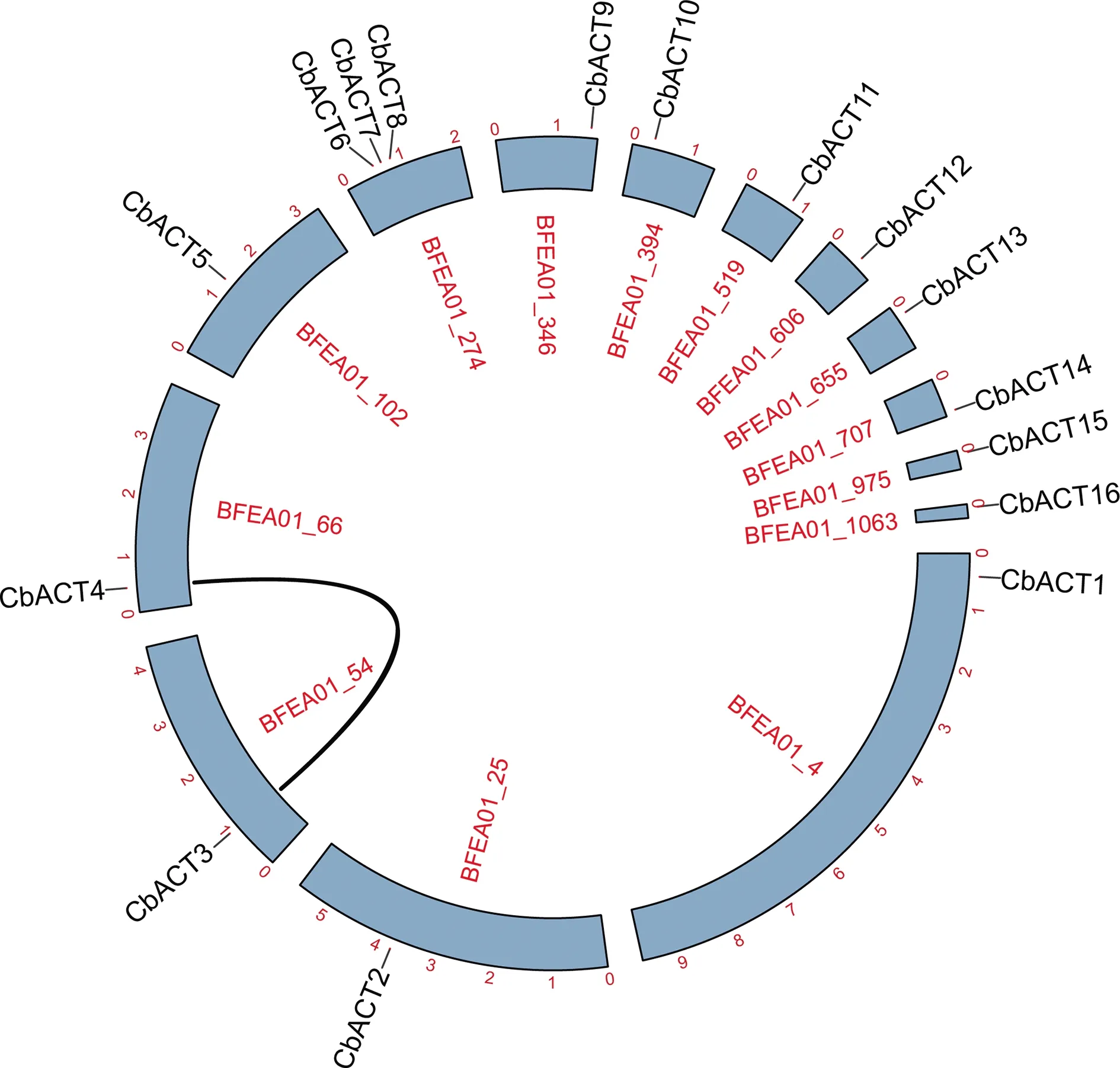
圖3 CbACT基因染色體定位及共線性分析
產生,大部分成員可能是跟隨物種進化遺傳而來.另外為了解Actin在不同物種間的進化遺傳關系,利用萊茵衣藻、克里藻的全基因組及其Actin基因家族與布氏輪藻的基因組、Actin基因家族進行了物種間的共線性分析,結果并未發現兩兩之間存在共線性關系.由于目前已解析輪藻綱植物及親緣關系較近的雙星藻綱、鞘毛藻綱基因組較少,因此Actin基因家族在藻類物種間的進化分析缺乏相關的數據支持,未來隨著藻類基因組數據庫的更加完善將會為不同基因家族在藻類中的進化分析提供一定的幫助.
為了解Actin基因家族可能參與的轉錄調控過程,本研究分析了16個Actin基因上游2000 bp包含的順勢作用元件.在16個Actin基因家族中共鑒定出1094個順勢作用元件,根據功能將其分為6種類型.所有的Actin基因都包含了大量的基本啟動子元件(AT-TATA-box、CAAT-box、TATA-box)和光響應元件(G-box、Sp1、TCCC-motif).其他順式作用元件主要包括脅迫響應元件包括低溫脅迫(LTR)、干旱脅迫(MBS)、厭氧響應(ARE、GC-motif)等;激素響應元件包括生長素(AuxRR-core)、赤霉素(GARE-motif)、脫落酸(ABRE、)、水楊酸(TCA)、乙烯(ERE)等響應元件.除此之外還有包含了組織特異性元件(CAT-box、RY-element)及與玉米黃素代謝有關的特殊元件(O2-site)(圖4和圖5).結果表明布氏輪藻的Actin基因廣泛參與了輪藻的生長發育、生物和非生物脅迫、激素調控等過程,不同成員之間具有不同數量、種類的啟動子元件,從而導致不同成員之間功能的差異性及多樣性.

圖4 CbACT啟動子順勢作用元件
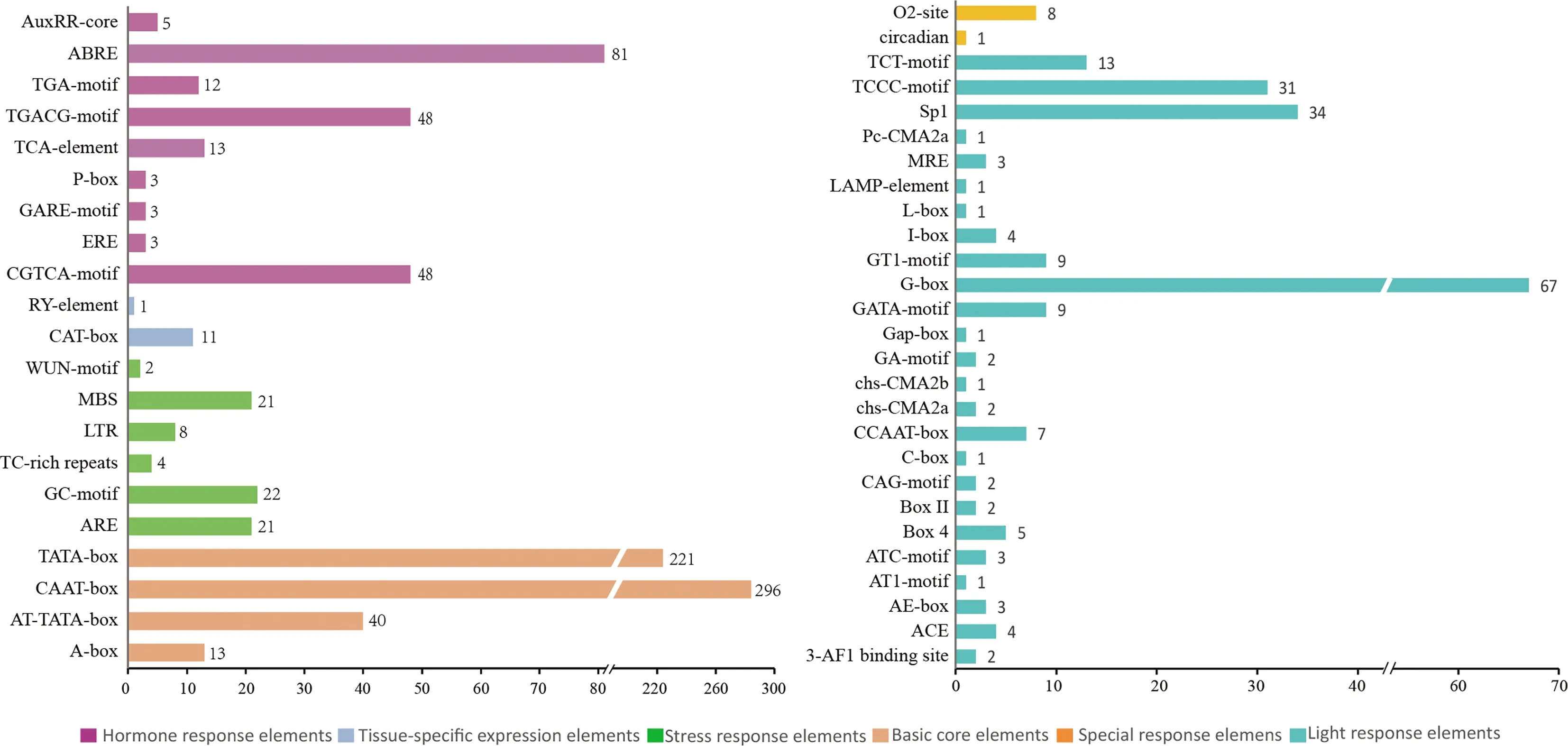
圖5 CbACT順勢作用元件統計分析
3.7 不同組織部位表達譜分析
為了解不同Actin基因在輪藻不同組織中的表達情況,本研究基于布氏輪藻RNA-Seq轉錄組數據,對布氏輪藻不同組織的Actin基因進行了表達量分析(圖6).大部分基因的表達量在各組織中表達量較小,CbACT4、CbACT10、CbACT14三個基因的表達量在四個組織中表達量較高,表明這些基因對于調控輪藻植物的生長發育發揮了較大的作用.CbACT5在合子、藏精器、藏卵器三個有性生殖器官中表達量較高,說明該基因可能主要影響了生殖器官的發育過程.另外由片段復制得到的CbACT4和CbACT3具有相似的表達模式,在各組織中表達量相似,表明這兩個基因還未發生功能的分化共同調節不同組織的發育過程.
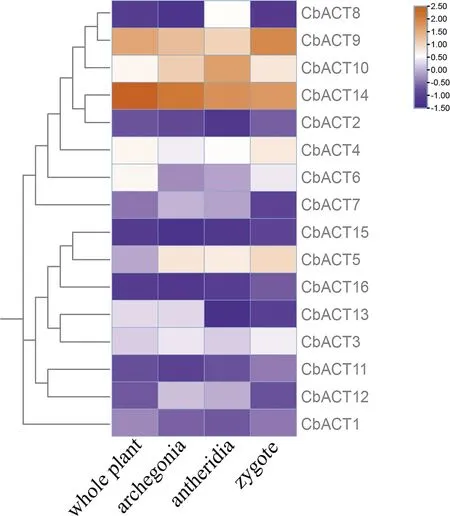
圖6 CbACT不同組織表達譜
4 討 論
Actin基因在調節植物形態建成、細胞運動等方面具有重要作用,本研究根據同源性分析和結構域篩選,在布氏輪藻中共獲得16個Actin基因,對該基因家族的二級結構分析表明,布氏輪藻的Actin基因家族主要以無規則卷曲為主,與葡萄的Actin二級結構分析結果相似,說明Actin基因家族的結構存在一定的保守性[12];Actin的亞細胞定位分析結果顯示,布氏輪藻的Actin基因除了CbACT13定位于葉綠體,其他15個Actin基因均定位于細胞質,這也與該基因家族主要參與細胞骨架的構成、細胞中物質運輸的功能分析結果相一致[13].擬南芥Actin7參與了細胞分裂時葉綠體的移動,因此CbACT13可能也參與了葉綠體相關的生命活動[14].系統發育分析結果顯示,輪藻的Actin基因家族可以分為較為結構保守的類型Ⅰ和結構變化較大的類型Ⅱ兩個分支.這也與Wu等[15]的研究結果類似,在藻類中Actin基因家族至少可以分為兩個分支,其中一個分支序列較為保守,而另外一個分支序列則存在較大的差異.除了典型的Actin結構域之外,布氏輪藻Actin基因還有F-box-like、PRK-13357、RT-like結構域.F-box like蛋白參與調控蛋白降解、信號轉導、逆境脅迫等生物學過程[16].RT_like廣泛存在于植物基因組中,在一定條件下可以發生轉座,從而調控靶向基因的失活、突變等,使得不同基因的表達調控更具有多樣性[17].更加復雜的結構域的存在使得輪藻植物的Actin基因家族的功能更加多樣化,從而更好地調節植物的環境適應性;保守基序分析表明,基序3、6、7存在于所有基因家族中,類型I相比與類型Ⅱ共有保守基序更多,這些差異的基序可作為兩個亞家族之間分類的依據;基因復制是基因家族發生擴增的主要方式,布氏輪藻基因組研究結果表明布氏輪藻未發生過全基因組加倍事件,因此基因家族的擴增方式可能主要來自于片段復制[18].但在布氏輪藻中Actin基因家族只有一對基因來自于片段復制,推測可能部分基因在植物逐漸進化過程中由于基因重排、基因轉座、基因丟失等原因而導致共線性基因對減少;基因的表達需要各種轉錄因子與啟動子不同區域結合,從而調控基因適應不同環境的變化,因此啟動子順勢作用元件的分析對于了解不同基因的結構、功能分析具有重要意義.對布氏輪藻的啟動子順勢作用元件分析結果表明布氏輪藻Actin基因家族啟動子區域除了基本啟動子元件(CAAT-box、TATA-box、A-box等)外廣泛存在光響應(G-box、TCCC-motif、Sp1等)、激素調控(AuxRR-core、ABRE、TGACG-motif等)、生物和非生物脅迫等元件(ARE、LTR、GC-motif等),說明Actin作為植物中重要的基因家族對于布氏輪藻環境適應、生長發育的調控、植物晝夜節律等發揮了重要作用;輪藻與其他綠藻門植物相比,出現了組織的分化,具有雌雄生殖器官、假根、小枝等,不同基因通過組織特異型表達調控不同組織的分裂分化.通過分析不同組織的表達譜表明,在布氏輪藻中Actin基因家族的功能發生了分化,不同家族成員在不同組織中表達量存在偏好性,有三個基因在不同組織中明顯高于其他組織,推測這幾個基因相對于其他家族成員對于輪藻的生長發育更為重要.另外,對于來自片段復制的基因對表達譜趨于一致,說明該基因對可能還未發生功能分化,共同參與了植物的生長發育過程.與擬南芥的Actin表達譜相比,在輪藻中Actin并未明顯的分為生殖型和營養型兩個類型,但有少數幾個Actin基因在生殖器官和營養器官的表達出現了較為明顯的差異,因此可能在低等的藻類植物中也存在兩種類型的基因,但是兩種類型的基因在序列相似性上尚未存在明顯的差異,所以推測被子植物出現的兩種類型的Actin基因是在跟隨植物進化過程中由古老的Actin基因逐漸發生變異從而最終分化形成了生殖型和營養型的Actin亞家族.
目前在植物Actin的研究主要集中擬南芥、豌豆、棉花等,但是在其他物種特別是在藻類植物中研究較少.因此Actin基因對輪藻及其他藻類的生長發育、形態建成、脅迫響應等功能的發揮及其內在的分子機制還需要進一步的深入研究.本研究從布氏輪藻全基因組中鑒定16個Actin基因家族,全面分析了Actin基因家族的蛋白理化特性、系統發育關系、基因結構、組織表達模式等,結果表明Actin基因家族廣泛參與了輪藻植物的生長發育、脅迫響應等生物學過程,研究結果為Actin基因基因家族在輪藻科中的系統發育研究及對輪藻生長發育的具體功能研究提供了一定的參考價值.
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
