干酪乳桿菌Zhang在不同溫度下的膜脂肪酸變化分析
2022-07-26 10:45:16包秋華代利霞梅邇藍
中國乳品工業 2022年7期
包秋華,代利霞,梅邇藍
(內蒙古農業大學乳品生物技術與工程教育部重點實驗室,農業農村部奶制品加工重點實驗室,呼和浩特 010018)
0 引言
益生菌菌株在食品工業中的成功應用取決于它們的功能特性,而功能特性的發揮需要保證益生菌菌株以存活狀態進入人體腸道[1]。乳酸菌在生產和應用過程中不可避免會受到不利于生存的環境條件變化的影響,如溫度、酸堿度、滲透壓、射線、寡營養、電解質和氧化劑等[2]。這些因素可能會引起乳酸菌基因型和表型發生一系列的生理變化[3]。其中溫度是對乳酸菌生長最具影響的環境因素。當在最適生長溫度下,繁殖速率最快,細胞結構完整,生長狀態最好,當高于或低于最適生長溫度時,乳酸菌會激活多種應激反應去應對不利生存的條件來抵抗脅迫[4-7]。Hermien V B[8]將植物乳桿菌置于28℃和40℃進行溫度脅迫,發現兩種轉錄調節因子有助于調節菌株對溫度脅迫的抗性,進而對其應激調節機制進行闡述。Correa D M A[9]對鼠李糖乳桿菌CRL 1505進行了耐熱性研究,55℃下處理20 min,細胞存活率沒有顯著變化,顯示出了高的內在耐熱性可見菌體對此逆境條件產生了耐熱機制。
細胞膜是菌體與外界環境隔離的主要屏障,當溫度發生劇烈變化時,細胞膜是首先感知到外界環境壓力的部位,也是首先被傷害的目標基質,這種傷害體現在破壞細胞膜的完整性。研究發現微生物的自我調節機制與細胞膜不飽和脂肪酸也有密切關系,細胞膜中的不飽和脂肪酸含量的增加可以增強微生物在溫度變化過程中細胞膜的流動性,起到維持細胞膜的功能,從而提高微生物的抗逆性[10-12],張愛靜[13]通過熱脅迫處理E.coil ATCC43889菌株,發現隨著溫度的升高,個體形態變長,生物被膜生成能力增強,飽和脂肪酸含量升高,不飽和脂肪酸含量下降,菌株耐熱性增強。乳酸菌在溫度變化過程中也可以通過改變細胞膜脂肪酸的含量與組分,調節細胞膜的流動性去維持菌株正常生長,減免損傷[14-15]。李寶坤[16]通過對羅伊氏乳酸桿菌冷熱脅迫處理后,采用Bligh&Dyer法對脂肪酸進行提取,發現羅伊氏乳酸桿菌CICC 6226菌體中的不飽和脂肪酸比例上升,而在此條件下菌體的存活率也最高,推斷在冷熱脅迫條件下,羅伊氏乳酸桿菌對此逆境產生了保護機制,這種機制產生后會對細胞膜脂肪酸的含量及組分產生很大的影響,對微生物環境適應性和應激脅迫響應機制的探究具有重要意義,對保障食品安全及企業面臨的生產問題有一定的改善。
干酪乳桿菌Zhang(Lactobacillus casei Zhang,Lb.casei Zhang)是一株從傳統酸馬奶中分離的益生菌,經過嚴格的科學實驗,對該菌的生理特性、益生功效、分子機理、生產性能等一系列關鍵技術進行了攻關研究,取得了重大突破,并證明了其安全性,且已市場化應用[17-18]。本文通過研究逆境條件下干酪乳桿菌Zhang膜脂肪酸的變化,從而探究其是否具有抵御不良生存環境下的應激機制。
1 材料與方法
1.1 材料
菌株:干酪乳桿菌Zhang(Lb.casei Zhang),內蒙古農業大學乳品生物技術與工程教育部重點實驗室乳酸菌菌種資源庫保存并提供。
試劑:液體MRS、固體MRS、生理鹽水、氯仿-甲醇溶液、氯仿、甲醇鈉-甲醇、正己烷。
1.2 儀器
DHP-9272恒溫培養箱,上海恒科有限公司;Eppendorf 5810 R離心機,德國Eppendorf公司;MLS-3750高壓蒸汽滅菌鍋,日本三洋公司;氣相色譜儀6850型,美國Agilent公司;EYELA MG-2200氮吹儀,日本。
1.3 實驗方法
1.3.1 菌株活化
將Lb.casei Zhang以2%的接種量接入MRS液體培養基中,置于37℃恒溫培養箱培養18~24 h,活化到第三代后,將對數生長末期的菌體通過革蘭氏染色進行形態觀察和電鏡掃描觀察微觀結構。
1.3.2 不同溫度條件Lb.casei Zhang的處理
將活化的菌液分裝到50 mL離心管中,5 000 g離心5 min,棄上清,加入適量生理鹽水洗菌,離心后收集菌泥,把預熱好的45、60、70℃MRS液體培養基分別倒入50 mL離心管中,在對應溫度下對Lb.casei Zhang熱處理5、10、30 min,同時在37℃下做對照試驗。
1.3.3 不同溫度條件Lb.casei Zhang活菌數量變化
在不同溫度條件下,采用稀釋平板計數法檢測Lb.casei Zhang在0、6、12、18 h的活菌數量,每次試驗重復3次,每次試驗3個平行。
1.3.4 膜脂肪酸的提取及甲酯化
對4種不同溫度處理的Lb.casei Zhang分別在0、6、12、18 h進行取樣,并采用Bligh&Dyer法[16]提取細胞膜脂肪酸并進行甲酯化。
1.3.5 脂肪酸組分和含量的測定
采用Agilent 6850氣相色譜儀對樣品進行脂肪酸檢測。先用37種脂肪酸甲酯標準品(Supelco 47885-U)定標,得到了能夠檢測37種脂肪酸甲酯標準品的最佳氣相檢測條件,然后通過氣相色譜法檢測Lb.casei Zhang在不同溫度條件不同時間的細胞膜脂肪酸組分和含量,每個樣品重復測定3次。
檢測條件:色譜柱為定制Agilent公司毛細填充管,填充物為(88%-氰丙基)芳基-聚硅氧烷,規格60 m×0.25 mm i.d×0.25μm film;載氣:氮氣(純度為99.999%);進樣口溫度:260℃;柱溫升溫程序:初始溫度80℃,保持1 min,隨后以6.5℃/min速率增至170℃,以2.75℃/min速率增至215℃,并保持2 min,以40℃/min速率增至230℃,并保持2 min;氫氣流量35 mL/min、空氣流量350 mL/min、氮氣流量35 mL/min;檢測器為氫火焰離子檢測器,檢測器溫度260℃;進樣量1μL,分流比1∶5。
1.3.6 數據分析
將待測樣本的脂肪酸色譜圖與標準色譜圖進行分析比較,確定待測樣品中脂肪酸組分,并用面積歸一化法(以峰值面積的百分比表示)確定各種脂肪酸的百分含量[19]。上述所有數據均采用SPSS等軟件計算平均值和標準差,并用Origin 2017軟件作圖。
2 結果與分析
2.1 Lb.casei Zhang的細胞形態學觀察
將活化第三代的Lb.casei Zhang進行革蘭氏染色和掃描電鏡觀察,結果見圖1。
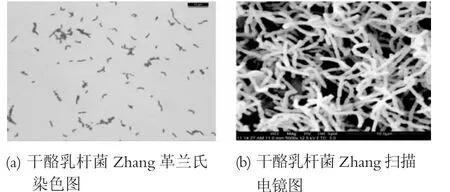
圖1 Lb.casei Zhang的細胞形態學觀察
通過圖1可以看出,Lb.casei Zhang為革蘭氏陽性短桿菌,大小形態均一,沒有其他雜菌的圖樣,可以用于后續實驗。
2.2 Lb.casei Zhang在不同溫度條件下活菌數變化結果
對Lb.casei Zhang進行不同溫度處理后,不同時間點的活菌數變化情況見圖2。結果顯示在不同溫度處理下,活菌數變化規律均不同。37℃和45℃處理時活菌數變化趨勢總體平穩,變化幅度較小,但37℃在0~18 h內先略微增加然后開始下降;60℃和70℃處理時活菌數在0~6 h出現大幅度下降,而在6~18 h出現緩慢降低,整體呈現下降趨勢。
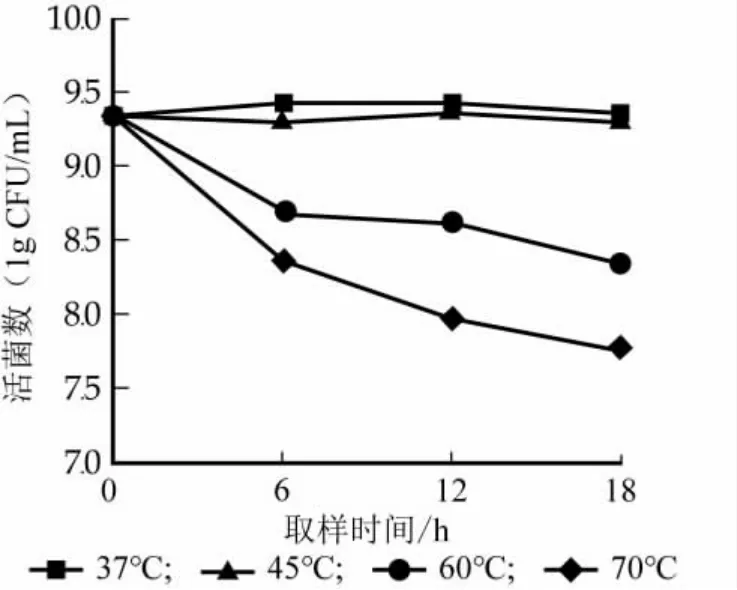
圖2 不同溫度脅迫下Lb.casei Zhang活菌數變化結果圖
2.3 Lb.casei Zhang在不同溫度條件下膜脂肪酸組分和含量的檢測
通過氣相色譜法檢測Lb.casei Zhang在不同溫度條件下不同時間點處理的脂肪酸,部分脂肪酸變化峰圖結果見圖3。從色譜圖的峰形、基線等方面看,數據很好,可信度高。
通過脂肪酸的氣相色譜圖可以看出脂肪酸含量發生了顯著變化。從圖3可以看出,Lb.casei Zhang在溫度為45℃處理0 h時油酸(C18∶1n9c)的峰高(出峰時間20.693 min)高于18 h(出峰時間20.673 min)的峰高。

圖3 Lb.casei Zhang在不同溫度條件下的脂肪酸氣相色譜圖
在不同溫度條件下,通過氣相色譜法測出主要發生變化的脂肪酸有5種∶肉豆蔻酸(C14∶0)、棕櫚酸(C16∶0)、棕櫚油酸(C16∶1)、油酸(C18∶1n9c)、亞油酸(C18∶2),這5種脂肪酸占總細胞膜脂肪酸的比例最大。不同溫度條件下飽和脂肪酸中以棕櫚酸為主,含量約占總細胞膜脂肪酸含量的28%,而不飽和脂肪酸中以亞油酸為主,占總膜脂肪酸含量的31.49%,脂肪酸種類見表1。

表1 Lb.casei Zhang的脂肪酸種類
2.4 不同溫度條件下Lb.casei Zhang細胞膜脂肪酸的變化分析
Lb.casei Zhang在不同溫度條件處理下不同時間點的膜脂肪酸相對百分含量變化,結果見表2。

表2 Lb.casei Zhang不同溫度下脂肪酸變化

(續表2)
由表2可見,Lb.casei Zhang在不同溫度下18 h內細胞膜中含量相對較高的5種脂肪酸變化均顯著,其中棕櫚酸(C16:0)是Lb.casei Zhang細胞膜的主要飽和脂肪酸,變化范圍在(25.28±0.12)%~(33.03±0.60)%,其次是肉豆蔻酸(C14∶0)。亞油酸(C18∶2)是主要的不飽和脂肪酸,變化范圍在(28.49±0.47)%~(37.24±0.62)%。
2.4.1 37℃條件下Lb.casei Zhang脂肪酸組分和含量變化
在37℃時,Lb.casei Zhang 5種脂肪酸相對百分含量以及總不飽和脂肪酸和總飽和脂肪酸相對百分含量的對比分析,結果見圖4。
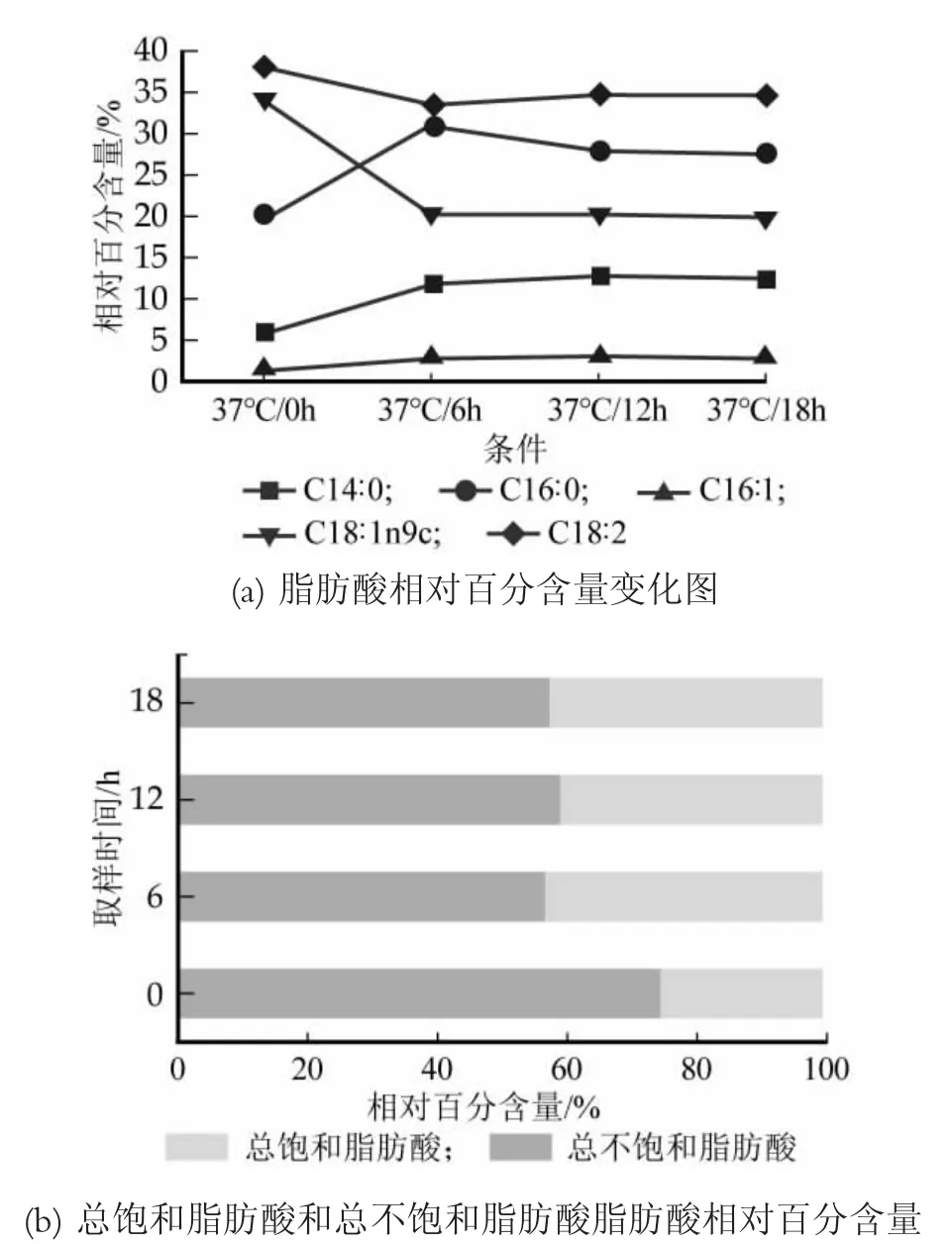
圖4 干酪乳桿菌Zhang在37℃條件下脂肪酸組分和含量變化
從圖4(a)可以看到,37℃條件下各脂肪酸相對百分含量隨時間的變化比較明顯,油酸(C18∶1n9c)和亞油酸(C18∶2)隨著時間的增加相對百分含量出現減少,而且變化浮動較大,在6 h之后基本平穩,僅出現略微的變化。肉豆蔻酸(C14∶0)、棕櫚酸(C16∶0)、棕櫚油酸(C16∶1)在0~6 h均出現了增幅,棕櫚酸(C16∶0)的增加幅度較大,且在6 h之后開始下降。此外,從圖4(b)中可以直觀的看出37℃條件下總飽和脂肪酸與總不飽和脂肪酸的相對百分含量變化,總不飽和脂肪酸在0~18 h內先減少后出現了增加,但增加幅度較小,總飽和脂肪酸相反,呈現先增加后減少的趨勢。
2.4.2 45℃條件下Lb.casei Zhang脂肪酸組分和含量變化
對Lb.casei Zhang在溫度為45℃時各脂肪酸相對百分含量以及總不飽和脂肪酸和總飽和脂肪酸相對百分含量進行了對比分析,結果見圖5。

圖5 干酪乳桿菌Zhang在45℃條件下脂肪酸組分和含量變化
從圖5(a)可以看到,45℃條件下各脂肪酸相對百分含量隨時間的變化,肉豆蔻酸(C14∶0)、棕櫚酸(C16∶0)和亞油酸(C18∶2)在0~6 h脂肪酸相對百分含量均出現了增加,油酸(C18∶1n9c)和棕櫚油酸(C16∶1)出現了下降,油酸的變化浮動較大,在6 h之后基本平穩,僅出現略微的變化。并且從圖5(b)中可以直觀的看到45℃下總飽和脂肪酸與總不飽和脂肪酸的變化,總不飽和脂肪酸在時間0~18 h內先減少后增加,總體趨勢為增加,總飽和脂肪酸相反,先出現增加然后減少。
2.4.3 60℃條件下Lb.casei Zhang脂肪酸組分和含量變化
對Lb.casei Zhang在溫度為60℃時各脂肪酸相對百分含量以及總不飽和脂肪酸和總飽和脂肪酸相對百分含量進行了對比分析,結果見圖6。
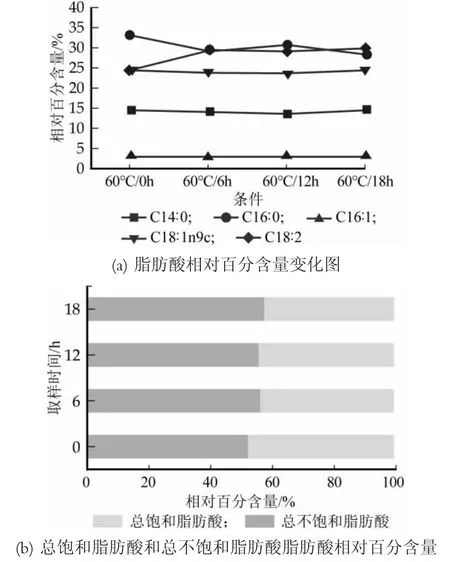
圖6 干酪乳桿菌Zhang在60℃條件下脂肪酸組分和含量變化圖
從圖6(a)可以看到,60℃條件下各脂肪酸相對百分含量隨時間的變化,肉豆蔻酸(C14∶0)、棕櫚酸(C16∶0)、棕櫚油酸(C16∶1)、油酸(C18∶1n9c)在0~6 h隨著時間的變化相對百分含量出現減少,亞油酸(C18∶2)相對百分含量出現增加,之后的變化也不顯著,并且從圖6(b)可以直觀的看出60℃下飽和脂肪酸與不飽和脂肪酸的變化,總不飽和脂肪酸在時間0~18 h內逐漸增加,但增加幅度較小,總飽和脂肪酸相反,這一結論與Ma YL[20]的研究結果相似。
2.4.4 70℃條件下Lb.casei Zhang脂肪酸組分和含量變化
對Lb.casei Zhang在溫度為70℃時各脂肪酸相對百分含量以及總不飽和脂肪酸和總飽和脂肪酸相對百分含量進行了對比分析,結果見圖7。
從圖7(a)可以看到,70℃條件下各脂肪酸相對百分含量隨時間的變化,棕櫚酸(C16:0)和油酸(C18:1n9c)在0~6 h先增加,從第6 h開始出現降低的趨勢,然后在12 h呈現略微的增加,肉豆蔻酸(C14:0)和亞油酸(C18:2)隨著時間的增加相對百分含量出現減少—增加—減少的趨勢,而棕櫚油酸(C16:1)在0~18h內相對百分含量的變化很微弱,且含量較低。此外,從圖7(b)中可以直觀的看出70℃下飽和脂肪酸與不飽和脂肪酸的變化,總不飽和脂肪酸在時間0~18 h內先減少后出現了增加,但增加幅度較小,總飽和脂肪酸相反,先增加后減少。
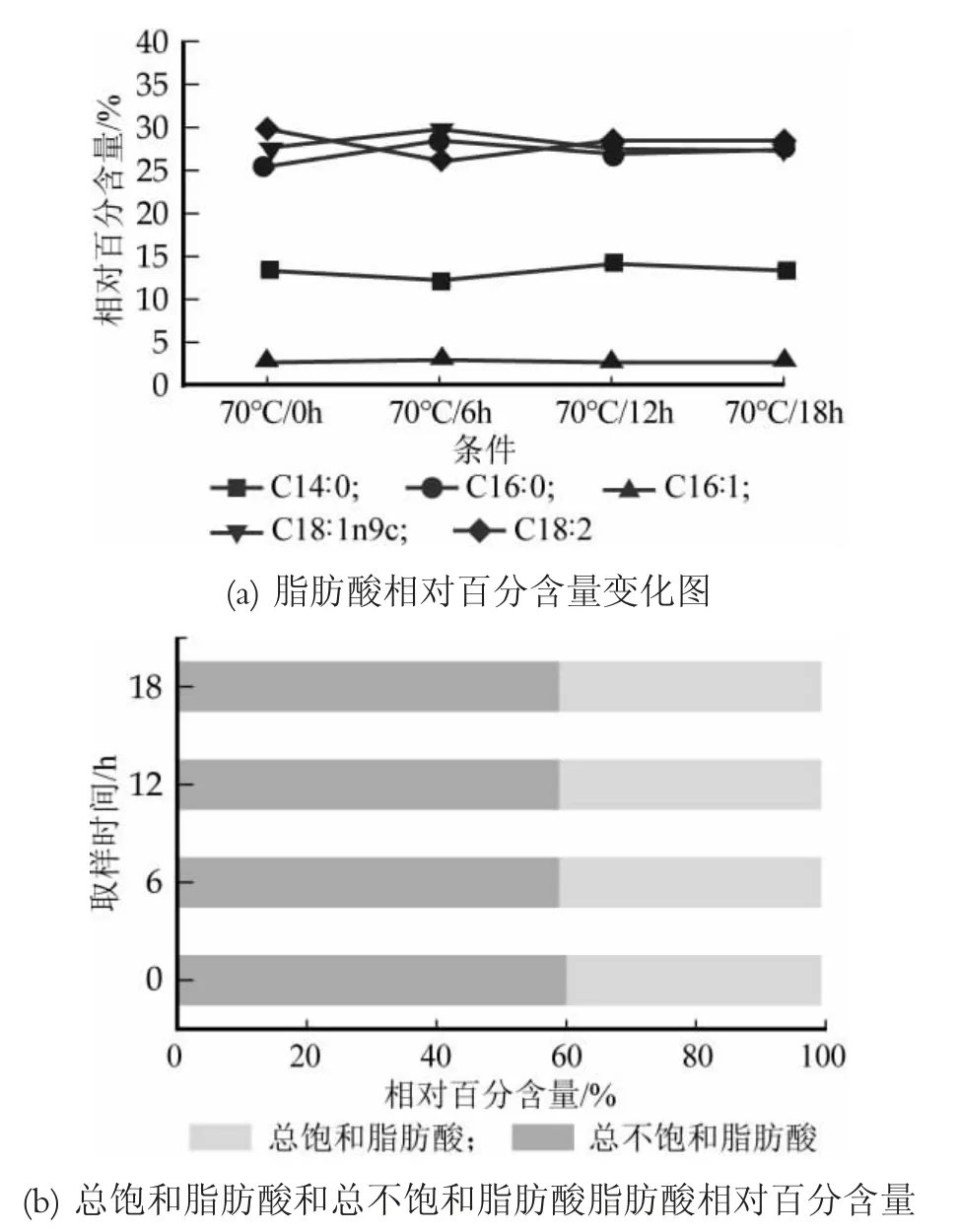
圖7 干酪乳桿菌Zhang在70℃條件下脂肪酸組分和含量變化
3 討論
當乳酸菌處于不同溫度脅迫下,它會激活多種應激反應去抵抗這些不適生長溫度的脅迫,其中細胞膜脂肪酸組成與含量的改變是其中一種,是細菌在不利條件下保持生存能力的重要保證。本研究中通過對Lb.casei Zhang在45、60、70℃條件下進行熱激處理,隨著處理溫度的升高,Lb.casei Zhang的存活率明顯降低;細胞膜脂中不飽和脂肪酸含量增加,而飽和脂肪酸含量減少,推斷Lb.casei Zhang在熱激處理條件下產生了抗逆性,為抵御高溫環境下的傷害,Lb.casei Zhang通過改變細胞膜脂肪酸組分和含量,增加細胞膜流動性來適應環境變化。這一結果與Vaessen E M J[21],Haddaji N[22]等研究結果一致,乳酸菌在逆境條件下,發生了應激反應,SFA/USFA(飽和脂肪酸/不飽和脂肪酸)比率下降,以及細胞膜流動性隨著處理溫度的升高而增加。阿根廷一位研究人員Gomez Z A[23]對8株嗜熱乳酸桿菌的脂肪酸組成和抗凍融性進行了研究,在-20℃與37℃下反復凍融循環,測定細胞膜中脂肪酸的比例以及飽和脂肪酸與不飽和脂肪酸的變化,發現C19∶0的增加可以使細胞免受凍融損傷。我們的研究中發現,當Lb.casei Zhang在45、60、70℃處理下,主要通過C18∶2(亞油酸)的增加來使細胞免受熱激損傷。Kentaro T[24]研究發現一些耐冷細菌響應生長環境變化也是通過調節它們的脂肪酸組成,因此脂肪酸組成的調節被認為是維持膜流動性所必需的。這也從側面解釋了我國具有自主知識產權的益生菌Lb.casei Zhang對熱有一定耐受性的機理。
實際生活中我們會盡可能避免含乳酸菌的食品受到不利環境的影響,而在工業生產中,會有意將乳酸菌置于脅迫條件下,將乳酸菌接種到腌制食品中,也就是高滲透壓環境,可以提高乳酸菌不飽和脂肪酸的含量,降低飽和脂肪酸的含量,改善腌制食品的品質以及風味質量[25],而且經過熱激處理的菌株,在制備發酵乳的過程中,會使發酵乳的后酸化顯著下降,對于提高發酵乳產品貨架期具有十分重要的現實意義[26]。利用這些脅迫響應可以生產出高質量的發酵食品,包括干酪、葡萄酒、酸奶、焙烤食品等。
4 結論
本研究通過在不同溫度下對Lb.casei Zhang進行脅迫處理,采用Agilen 6850氣相色譜儀和60米的色譜柱成功檢測了其脂肪酸種類和含量的變化,并對飽和脂肪酸與不飽和脂肪酸相對百分含量進行了研究。得到以下結論:在不同溫度脅迫下,主要發生變化的5種脂肪酸是肉豆蔻酸(C14∶0)、棕櫚酸(C16∶0)、棕櫚油酸(C16∶1)、油酸(C18∶1n9c)、亞油酸(C18∶2)。從相對百分含量上看,不飽和脂肪酸含量增加,飽和脂肪酸含量減少,推測Lb.casei Zhang為應對不良環境變化,通過增加不飽和脂肪酸含量來改變細胞膜流動性,增加細胞活性,起到了維持細胞膜的功能,從而提高了微生物的抗逆性,初步解析了Lb.casei Zhang具有抵御不良生存環境下的應激機制。
