咬合改變和束縛應激中小鼠全腦神經激活的比較研究
2022-07-29 05:03:28劉偉才
口腔醫學 2022年6期
徐 明,李 娟,劉偉才,張 琪
臨床中有一類與焦慮癥有關的現象,以“無法被客觀檢查證實的咬合異常”為主訴癥狀[1-2],患者表現出對咬合和咬合治療的高意識性且通常認為異常咬合的產生與牙科操作相關[3]。研究人員認為這種癥狀是由中樞神經系統的變化造成的。Marbach等[4]推斷其病因為中樞神經系統中神經矩陣(neuromatrix)的變化;Clark等[5]、Tsukiyama等[6]認為是三叉神經系統的功能異常導致上述癥狀的產生;然而關于其病因學的假設均局限于個人判斷和有限的臨床觀察,在病例解釋中具有一定的局限性[3]。
既往研究中利用嚙齒類動物的咬合升高模型(磨牙粘接樹脂或粘接金屬冠)發現,在咬合升高后下丘腦-垂體-腎上腺(HPA)軸被激活、血清皮質酮水平增加且動物表現出焦慮樣行為[7-8]。通過對神經元活性標志物cfos蛋白進行免疫化學染色發現咬合升高手術后海馬[9]、杏仁核和臂旁核[10]中激活的神經元數目顯著增加。但咬合升高中全腦神經元激活情況目前尚不可知。本研究將觀察小鼠咬合升高后全腦神經元的激活情況,并與常用的致小鼠焦慮行為的束縛刺激進行對照,分析兩種刺激中共同激活的部位。
1 材料與方法
1.1 實驗模型構建
1.1.1 咬合升高小鼠模型構建 選擇12周齡C57BL/6J清潔級雄性小鼠(雷根生物科技有限公司,中國),體質量為(30±5)g(倫理編號:SL2021DW17)。小鼠在2%濃度的異氟烷中誘導麻醉,起效后轉至1%濃度異氟烷面罩持續麻醉。將小鼠固定于小鼠固定器中,暴露上頜磨牙,棉球干燥牙面,涂布樹脂水門汀(Kerr,美國),光固化燈固化20 s,在咬合面充填0.1~0.2 mm的光固化樹脂層,光固化40 s。
1.1.2 束縛刺激小鼠模型構建 將小鼠置于50 mL離心管中(離心管上烙制數十個通風孔),當動物進入后用墊料填塞剩余空間,便于根據不同小鼠的大小限制小鼠活動。2 h后將小鼠用于免疫熒光染色實驗。
1.2 行為學測試
將小鼠隨機分為2組:咬合升高組(OVR組)和假手術組(Sham組)。OVR組小鼠按1.1.1處理,Sham組小鼠采用同樣的操作流程,但是不涂布樹脂水門汀、不充填光固化樹脂。
1.2.1 曠場測試 測試于咬合升高后第7天進行。使用40 cm×40 cm×40 cm的方形不透明實驗箱,測試前1 h將小鼠從飼養間轉移至測試房間適應,降低動物對新環境的不安情緒。實驗前用75%的酒精清潔實驗箱,確保實驗箱清潔無味道,將動物從飼養籠中輕輕取出,置于箱子中央,攝像機記錄10 min。每只動物實驗結束后均需用75%乙醇再次清潔實驗箱。使用ANY-MAZE軟件(Stoelting,美國)分析視頻數據,在軟件中設置24 cm×24 cm的中心區域,中心區域外為邊緣區域,輸出小鼠在中心區域內的停留時間、運動距離、進入次數等。
1.2.2 高架十字迷宮測試 測試于咬合升高后第8天進行。使用包括35 cm×5 cm×16 cm的4個臂(兩個相對的閉合臂和兩個相對的開放臂,開放臂與中心區域相對的臂不封閉)和5 cm×5 cm×16 cm的正方形中央區域的實驗裝置,固定于離地面50 cm的金屬制十字架上。測試前1 h將小鼠從飼養間轉移至測試房間適應,實驗前用75%的乙醇清潔實驗箱,將動物從飼養籠中輕輕取出,頭朝開放臂方向置于箱子中央,攝像機記錄5 min,每只動物實驗結束后需用75%酒精再次清潔實驗箱。使用ANY-MAZE軟件分析視頻數據,輸出小鼠在開放臂的停留時間、運動距離和進入次數等。
1.3 免疫熒光染色
將小鼠隨機分為4組:OVR組、Sham組、RS組、Ctrl組。OVR組、Sham組模型構建同1.2;RS組模型構建同1.1.2;Ctrl組將小鼠放入新籠,單獨飼養,不做其他處理。2 h后將小鼠按以下步驟處理。1%戊巴比妥鈉(1 mL/kg)經小鼠腹腔注射麻醉,經左心室灌流20 mL的4 ℃生理鹽水后,用4%多聚甲醛灌流固定,剝離取出腦組織。將取出的腦組織放入4%多聚甲醛,并于4 ℃冰箱中固定6~8 h,30%的蔗糖溶液中脫水兩次,包埋后使用冰凍切片機制備35 μm的冠狀切片。配制含有10%正常驢血清、1%牛血清白蛋白、0.3% TritonX-100及1×磷酸鹽緩沖液(PBS)的抗體稀釋液,將切好的腦片置于含有兔抗cfos一抗(1∶2 000)(ABCAM,英國)的24孔板中,4 ℃搖床孵育過夜。孵育后PBS漂洗3次,每次10~15 min。加入驢抗兔二抗(1∶1 000)(Invitrogen,美國),室溫孵育1 h,孵育后PBS漂洗3次,每次5 min。加入Hochest(1∶10 000)(Sigma,美國),復染細胞核10 min。待貼片風干后,使用含防熒光淬滅劑的封片液封片。Olympus熒光顯微鏡VS200(Olympus,日本)在相同的曝光時間下拍攝腦片,根據第3版小鼠腦圖譜[11]利用Photoshop軟件繪制腦區邊界,Image J軟件將獲取熒光圖像按照色彩通道分離,在得到的黑白圖片中統計陽性細胞數。
1.4 統計學分析

2 結 果
2.1 行為學測試
2.1.1 曠場測試 OVR組小鼠在咬合升高后第7天進入曠場中央區域的次數、在中央區域的運動距離均低于Sham組(P<0.05),表明OVR組小鼠焦慮樣行為顯著增加,但兩組間中心區域時間和總運動距離差異無統計學意義(P>0.05)(圖1),表明在該時間段內咬合升高對小鼠運動能力及自發活動無明顯影響。

A:中心區域時間;B:中心區域進入次數;C:中心區域運動距離;D:總運動距離;E:OVR組軌跡圖;F:Sham組軌跡圖;*:P<0.05,ns:P>0.05
2.1.2 高架十字迷宮測試 OVR組小鼠在咬合升高后第8天在開放臂的時間、在開放臂的運動距離均低于Sham組(P<0.05),兩組間開放臂進入次數和總運動距離差異無統計學意義(P>0.05)(圖2),同樣表明OVR組小鼠焦慮樣行為顯著增加,咬合升高在該時間段對小鼠運動能力及自發活動無明顯影響。
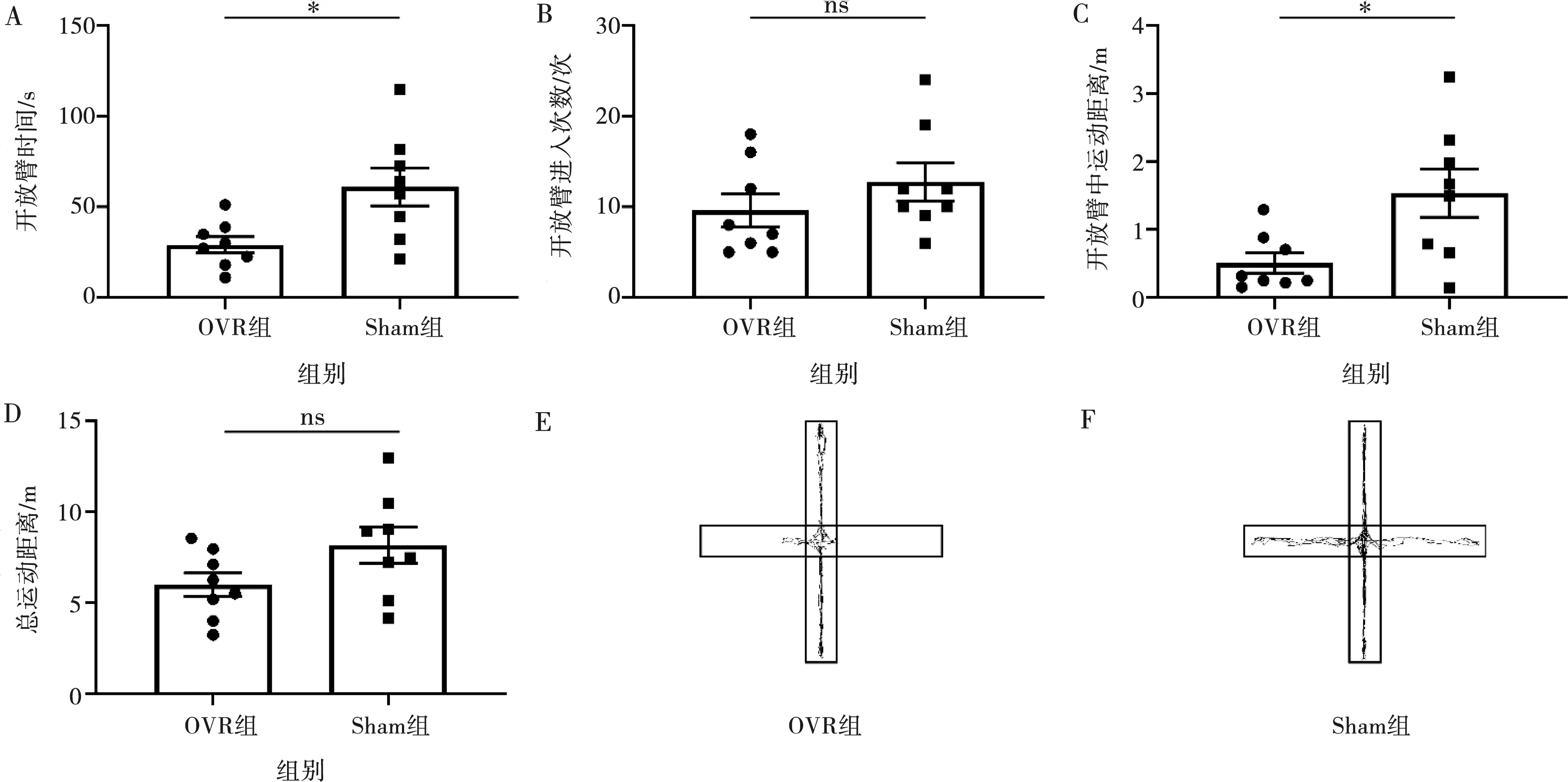
A:進入開放臂時間;B:開放臂進入次數;C:開放臂運動距離;D:總運動距離;E:OVR組軌跡圖;F:Sham組軌跡圖;*:P<0.05,ns:P>0.05
2.2 cfos表達變化
2.2.1 咬合升高后cfos表達變化 咬合升高2 h后,OVR組小鼠內側前額葉(medial prefrontal cortex,mPFC)、扣帶回皮層(cingulate cortex,Cg)、屏狀核(claustrum,Cl)、島葉皮層(insularcortex,INS)、伏隔核(nucleus accumbens,NAC)、外側隔核(lateralseptal,LS),包括外側隔核腹側部分(LSV)和背側部分(LSD)、下丘腦室旁核(paraventricular hypothalamic nucleus,PVN)、基底外側杏仁核(basolateralamygdaloid nucleus,BLA)、中央杏仁核(centralamygdaloid nucleus,CeA)、外側韁核(lateral habenula,LHb)、下丘腦背內側核(dorsomedialhypoth-alamic nucleus,DM)、底丘腦旁核(parasubthalamic nucleus,PSTN)、臂旁核(parabrachial nucleus,PB)、孤束核(nucleus of the solitary tract,NTS)中cfos陽性細胞數較Sham組顯著增加(P<0.05)(圖3),提示這些腦區中神經元激活程度增加。
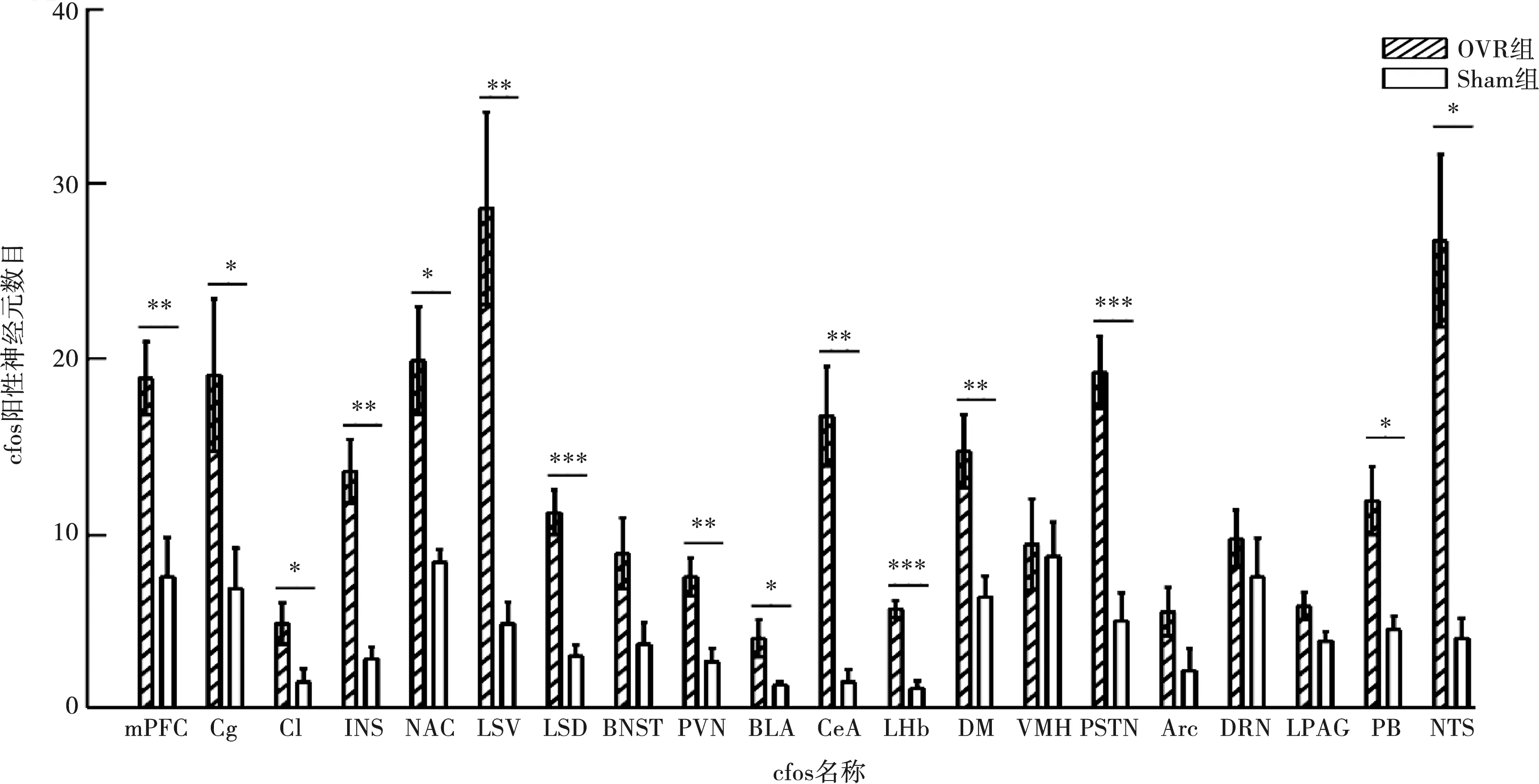
*:P<0.05,**:P<0.01,***:P<0.001
2.2.2 束縛刺激后cfos表達變化 RS組小鼠Cl、NAC、LS、PVN、弓狀核(arcuatehypothalamic nucleus,Arc)、中縫背核(dorsalraphe nucleus,DRN)、NTS中cfos陽性細胞數較Ctrl組顯著增加(P<0.05)(圖4)。Cl、NAC、LS、PVN、NTS中cfos陽性細胞數在OVR組及RS組均顯著增加,提示這些腦區中神經元在兩種不同的刺激中激活程度均增加(圖4、5)。

*:P<0.05,**:P<0.01,***:P<0.001

A:咬合升高后cfos表達情況;B:束縛刺激后cfos表達情況;圖中為免疫熒光染色(色彩通道分離后),虛線為腦區邊界
3 討 論
本研究發現咬合改變會導致小鼠焦慮樣行為增加,咬合升高后mPFC、Cg、Cl、INS、NAC、LS、PVN、BLA、CeA、LHb、DM、PSTN、PB、NTS等部位表達cfos蛋白的神經元數目增加。尚琳等[10]對小鼠進行咬合升高后發現,手術后第7、14天小鼠焦慮樣行為增加,PB及CeA中神經元被激活,與本實驗結果一致。但本研究觀察到在急性咬合改變操作之后,全腦中還有其他被激活的神經元,這些神經元分布的部位與目前認為調控焦慮的神經環路有很多重疊[12],重疊的部位可能是咬合改變與焦慮癥狀聯系的關鍵。
曠場實驗是利用小鼠對明亮開放區域厭惡的天性衡量小鼠的焦慮水平,焦慮增加的小鼠更傾向于靠近邊緣區域[13]。高架十字迷宮實驗利用動物對新異環境的探究特性和對高懸開放臂的恐懼形成矛盾沖突行為測試動物的焦慮狀態[14]。咬合改變后小鼠進入曠場中央區域的次數和在中央區域的運動距離,在高架十字迷宮開放臂的時間以及在開放臂的運動距離均顯著減少,說明咬合改變可以導致小鼠的焦慮樣行為增加,這與臨床中咬合操作和焦慮癥的關聯一致[1]。
束縛刺激是常用的誘導焦慮樣行為方法,研究顯示急性束縛后嚙齒類動物的焦慮樣行為增加[15],cfos蛋白是神經元活化的標志物,其在各種刺激后短暫且快速地表達,刺激后2 h表達水平最高[16]。本實驗利用束縛刺激中被激活的神經元做對照,觀察該刺激中激活的神經元與咬合改變刺激中被激活神經元的重疊部分,這些在兩種刺激中均顯著激活的部位可能是咬合改變導致焦慮癥的致病機制。急性束縛刺激2 h后Cl、NAC、LS、PVN、Arc、DRN、NTS等部位表達cfos蛋白的神經元數目增加。Cl、NAC、LS、PVN、NTS在兩種刺激中表達cfos蛋白的神經元數目均顯著增加。既往研究顯示這些部位與焦慮樣情緒的產生密切相關。Cl在機體暴露于急性應激源時激活,被認為在控制由壓力引起的焦慮行為中至關重要[17];NAC是大腦獎賞和動機系統的重要組成部分,是嚙齒動物焦慮行為神經環路的重要環節[18];LS雖然在某些研究中顯示其輸出的功能具有抗焦慮的作用,但是最近也有研究顯示LS中表達Ⅱ型促腎上腺皮質激素釋放因子受體(CRFR2)神經元的激活可產生持續性的焦慮,并可導致血液中的皮質醇水平增加[19];PVN是HPA軸的重要組成部分,PVN中促腎上腺皮質激素釋放激素(CRH)神經元可沿HPA軸啟動激素級聯反應應對壓力狀態[20];NTS中大量表達糖皮質激素受體,該受體的功能抑制可以降低大鼠高架十字迷宮中開放臂的探索行為[21]。
大腦對咬合改變與束縛刺激這兩種應激狀態的反應也存在差異,咬合改變后mPFC、Cg、INS、BLA、CeA、LHb、DM、PSTN、PB顯著被激活,而束縛刺激后上述腦區并未明顯激活,而Arc、DRN在束縛刺激后顯著激活。差異性表達的腦區提示兩種刺激引起的應激反應有所差異[10,22-23]。對這些腦區的關注可能可以從其他角度理解咬合改變對機體的影響。
綜上所述,本研究從神經元的活動水平初步發現咬合改變后與焦慮行為相關部位的神經元顯著激活,但這些神經元的激活是否直接導致小鼠的焦慮樣行為尚不明確,今后可通過神經元的功能操縱,如利用光遺傳學或化學遺傳學的方法進一步對其功能進行探索和驗證。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
學苑創造·A版(2020年9期)2020-10-13 09:41:02
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
中國科技博覽(2016年2期)2016-04-25 20:32:39
小學生導刊(2016年34期)2016-04-11 00:49:44
電測與儀表(2015年5期)2015-04-09 11:30:52
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
