基于TMT的定量蛋白質組學技術解析鹽脅迫提高庫德畢赤酵母耐熱性機制
2022-08-02 03:10:50劉秋影李春生楊賢慶王悅齊吳燕燕馬海霞
食品科學 2022年14期
關鍵詞:酵母菌
劉秋影,李春生*,楊賢慶*,王悅齊,吳燕燕,馬海霞
(1.中國海洋大學食品科學與工程學院,山東 青島 266003;2.中國水產科學研究院南海水產研究所,農業農村部水產品加工重點實驗室,國家水產品加工技術研發中心,廣東 廣州 510300)
發酵食品是一類利用微生物作用賦予食品營養與獨特風味并可延長食品保存期的加工食品,在中國有著悠久歷史和豐富內涵,是我國食品行業的特色產業之一。酵母菌是發酵食品中最常用的微生物之一,其乙醇發酵作用在白酒、啤酒、黃酒、葡萄酒、食醋等發酵食品的釀造過程中發揮重要功能。傳統酵母菌對乙醇發酵的溫度要求較高,其最適發酵溫度為28~33 ℃,一般不超過36 ℃,然而發酵過程中不斷釋放的熱量會造成發酵體系溫度升高,使酵母菌活性降低,最終導致乙醇產量下降。庫德畢赤酵母(),原稱東方伊薩酵母(),是食品中常見的酵母菌,已從多種發酵食品體系中分離鑒定,在中國白酒、紅酒、龍舌蘭酒、ogi飲料、togwa等發酵食品的風味形成上發揮著重要作用。庫德畢赤酵母是多耐性微生物,能夠耐受多種脅迫條件,如熱、高鹽、高濃度乙醇、高滲透壓、強酸、高濃度重金屬等,可以適應復雜的食品加工環境,在食品工業領域具有廣闊的應用前景。庫德畢赤酵母在高溫條件下(34~42 ℃)具有良好的乙醇生產能力。然而,當發酵溫度超過45 ℃,庫德畢赤酵母的生長及乙醇產量受到極大抑制。
在對耐性微生物應激防御機制的研究中發現,多種脅迫共同作用會對微生物產生交互保護作用,即當微生物對某一種逆境因子啟動防御保護時,會激發多種與壓力脅迫相關基因的表達,促使微生物對其他逆境因子的抵抗能力也得到增強。據報道,鹽酸脅迫能夠顯著提高干酪乳桿菌的交互脅迫抗性,其引發的生理應答效應使細胞在應對熱致死和氧致死脅迫下的存活率顯著提高。氧化應激脅迫增加了熒光假單胞菌SN15-2細胞內氧化應激反應相關蛋白和其他對細胞有保護作用蛋白的表達,從而提高了菌株的耐熱性。前期研究發現,無機鹽脅迫能夠顯著提高庫德畢赤酵母的熱耐性以及在高溫條件下的乙醇發酵能力,然而鹽脅迫下庫德畢赤酵母耐熱性提高機制還未明確。
近年來,蛋白質組學研究方法發展迅速,基于串聯質譜標簽(tandem mass tags,TMT)的定量蛋白質組學技術是目前差異表達蛋白(differentially expressed proteins,DEPs)定量分析方法中通量最高、系統誤差最小、功能最強大的分析方法之一,具有高靈敏度、高分離能力、高通量等特點。本研究以多耐性庫德畢赤酵母A16為研究對象,利用TMT定量蛋白質組學技術分析單獨熱脅迫組和熱-鹽共脅迫組之間的蛋白質組差異,篩選與耐熱性提高相關的關鍵蛋白,在蛋白質組水平闡明鹽脅迫提高庫德畢赤酵母耐熱性的分子機制,以期為耐熱酵母菌的基因工程改造提供重要技術基礎,對提高酵母菌的高溫乙醇發酵能力具有重要意義。
1 材料與方法
1.1 材料與試劑
庫德畢赤酵母A16分離自中國高溫白酒酒曲。
酵母浸出粉胨葡萄糖(yeast extract peptone dextrose,YEPD)斜面培養基:1%酵母浸粉、2%蛋白胨、2%葡萄糖、2%瓊脂,pH 5.0;YEPD液體培養基:1%酵母浸粉、2%蛋白胨、2%葡萄糖,pH 5.0。
蛋白質羰基含量測試盒、微量丙二醛(malondialdehyde,MDA)測試盒 中國生物工程南京建成生物工程研究所;TMT試劑、Pierce BCA試劑盒美國Thermo Fisher Scientific公司。
1.2 儀器與設備
C色譜柱、高效液相色譜儀、串聯質譜儀 美國Thermo Fisher Scientific公司;超聲波細胞破碎儀 江蘇波場智能科技股份有限公司;數顯式穩壓穩流電泳儀上海天能科技有限公司。
1.3 方法
1.3.1 酵母菌活化
用接種環將YEPD斜面保存的酵母菌接種到新鮮的YEPD斜面上。30 ℃培養24 h后,用接種環輕輕刮取菌體接種到YEPD液體培養基中,30 ℃、180 r/min搖床培養24 h。
1.3.2 鹽脅迫對酵母菌耐熱性和氧化損傷的影響
將活化培養的酵母菌接種到含有不同NaCl濃度(0~500 mmol/L)的YEPD液體培養基中,控制初始生物量為0.1 g/L(干質量)。45 ℃、180 r/min水浴振蕩培養24 h后,發酵液5 000 r/min離心10 min,去上清液,菌體用超純水洗滌2 次,然后在105 ℃烘干至質量恒定,計算酵母菌的生物量(g/L)。
將在不同脅迫條件下培養的酵母細胞離心,用磷酸鹽緩沖液洗滌2 次,菌體用500 W超聲破碎儀破壁30 min(10 s/10 s),4 ℃、12 000 r/min離心15 min取上清液,分別采用微量丙二醛測試盒和蛋白質羰基含量測試盒測定細胞內MDA含量(nmol/mg)和蛋白質羰基含量(nmol/mg)。
1.3.3 基于TMT的定量蛋白質組學分析
1.3.3.1 蛋白質提取
將活化培養的酵母菌分別接種到含有0 mmol/L NaCl(T45組)、100 mmol/L NaCl(T45S100組)和300 mmol/L NaCl(T45S300組)的YEPD液體培養基中,45 ℃、180 r/min水浴振蕩培養24 h后,發酵液5 000 r/min離心10 min,菌體用磷酸鹽緩沖液洗滌2 次后,加入適量的蛋白質裂解液(8 mol/L尿素、1%十二烷基硫酸鈉,含蛋白酶抑制劑),冰上超聲2 min后裂解30 min,4 ℃、12 000h離心30 min,取蛋白質上清液。使用Thermo Scientific Pierce BCA試劑盒進行蛋白定量,并用十二烷基硫酸鈉-聚丙烯酰氨凝膠電泳檢驗蛋白完整性。
1.3.3.2 酶解烷基化以及TMT標記
取蛋白樣品100 μg,用裂解液補充體積到90 μL。加入終濃度10 mmol/L三(2-羧乙基)膦還原劑,37 ℃反應60 min。加入終濃度40 mmol/L碘乙酰胺,室溫下避光反應40 min。每管各加入預冷的丙酮(丙酮-樣品體積比6∶1),-20 ℃沉淀4 h,10 000h離心20 min,取沉淀。用50 mmol/L三乙基碳酸氫銨緩沖液充分溶解樣品,按照酶-蛋白質量比1∶50加入胰蛋白酶在37 ℃酶解過夜。TMT試劑加入乙腈解凍重組,并加入多肽溶液中進行標記,室溫孵育2 h;加入羥胺,室溫反應15 min,將等量標記產物混合于一管中,真空濃縮儀抽干。
1.3.3.3 液相色譜-串聯質譜(liquid chromatographytandem mass spectrometry,LC-MS/MS)檢測
采用納升級LC-MS/MS技術(Easy-nLC 1200結合Q Exactive質譜儀)進行分析。肽段用質譜上樣緩沖液溶解,上樣后經C色譜柱(75 μmh25 cm)分離120 min,體積流量為300 μL/min。EASY-nLC液相梯度洗脫,流動相A為0.1%甲酸-2%乙腈,B為0.1%甲酸-80%乙腈。按照以下程序進行梯度洗脫:0~1 min,100%~95% A、0%~5% B;1~63 min,95%~77% A、5%~23% B;63~88 min,77%~52% A、23%~48% B;88~89 min,52%~0% A、48%~100% B;89~95 min,0% A、100% B。MS和MS/MS采集之間自動切換,質譜分辨率分別是70 000和35 000。MS掃描范圍為/350~1 300,選擇top20的母離子進行二級碎裂,動態排除時間為18 s。
1.3.3.4 蛋白質比對及DEPs篩選
以庫德畢赤酵母129的蛋白質組(GCF_001983325.1)作為參考蛋白質組,對質譜下機的原始文件經Proteome Discoverer進行分析。肽段鑒定的錯誤發現率(false discovery rate,FDR)設置為FDR≤0.01。使用R語言中的test函數計算樣本間差異顯著性,同時計算組間蛋白質表達差異倍數(fold change,FC)。以1.5 倍差異表達,即|logFC|>0.58且<0.05作為DEPs的篩選標準。
1.3.3.5 生物信息學分析
將鑒定到的蛋白質序列與基因本體論(Gene Ontology,GO)、直系同源群集(Cluster of Orthologous Group,COG)、京都基因與基因組百科全書(Kyoto Encyclopedia of Genes and Genomes,KEGG)、Pfam、亞細胞定位等數據庫進行比對,獲得蛋白在各數據庫的注釋信息。
1.4 統計學分析
生物量結果采用SPSS 18.0軟件對數據進行單因素方差分析(ANOVA),應用多重比較Tukey檢測對數據之間的差異性進行檢驗,<0.05,差異顯著。采用主成分分析(principal component analysis,PCA)評價不同處理下庫德畢赤酵母蛋白質組的相似性。利用TBtools構建不同比較組DEPs熱圖。
2 結果與分析
2.1 鹽脅迫對酵母菌耐熱性和熱脅迫誘導的氧化損傷的影響
如圖1A所示,不同NaCl濃度(100~500 mmol/L)作用下,庫德畢赤酵母在45 ℃熱脅迫下的生物量顯著提高,表明鹽脅迫對庫德畢赤酵母的熱脅迫具有明顯的交互保護作用。特別是當NaCl濃度為300 mmol/L時,鹽脅迫的交互保護作用最為顯著,此時庫德畢赤酵母的生物量較無鹽脅迫時提高了2.6 倍。細胞中多不飽和脂肪酸極易受到活性氧(reactive oxygen species,ROS)攻擊,使其發生過氧化反應,產物MDA常作為脂質過氧化的指標,而蛋白質羰基化是蛋白質氧化損傷最主要表現之一,蛋白質羰基含量可以反映細胞蛋白質氧化損傷程度。由圖1B可見,與正常培養(30 ℃)相比,熱脅迫下(45 ℃)庫德畢赤酵母細胞內MDA含量和蛋白質羰基含量顯著增加。鹽脅迫后,庫德畢赤酵母在熱脅迫下

圖1 鹽脅迫對庫德畢赤酵母菌株耐熱性(A)和氧化損傷(B)的影響Fig.1 Effect of salt stress on thermotolerance (A) and oxidative damage (B) of P.kudriavzevii
2.2 不同脅迫處理下酵母菌全蛋白功能注釋
為了揭示鹽脅迫提高庫德畢赤酵母耐熱性的作用機制,利用TMT定量蛋白質組學技術研究庫德畢赤酵母在單獨熱脅迫組(T45組)和熱-鹽共脅迫組(T45S100組和T45S300組)之間的蛋白質組差異。3 個處理組共鑒定到3 875 個蛋白質,基于每個處理組中蛋白質種類及含量,利用Pearson相關性分析和PCA對3 個處理組全蛋白的相似度進行研究。Pearson相關性越接近于1表明樣本間的蛋白組成相似度越高,圖2A結果顯示,平行樣品間的相似度最高,T45組與T45S100組全蛋白的差異性較小,而與T45S300組的差異性較大。圖2B PCA結果顯示,PC1和PC2可以反映全部信息的73.80%,提取較為完全,這說明這2 個PC能夠替代原始成分反映樣品信息。其中PC1對整體結果影響最大,代表了全部信息的56.40%,PC1結果顯示,平行樣品間距離最近,T45組與T45S100組距離較近,而與T45S300組較遠,說明T45組與T45S100組全蛋白的相似度較高,而與T45S300組的相似度較小。結果表明,高鹽脅迫能顯著改變庫德畢赤酵母在熱脅迫下的蛋白質表達。

將鑒定到的蛋白序列與GO、KEGG、COG、Pfam、亞細胞定位等數據庫進行比對,獲得蛋白質在各數據庫的注釋信息(圖2C)。共有2 073 個蛋白獲得COG功能注釋(圖2D),其中注釋到最多個蛋白質的功能,包括功能未知(687 個),翻譯、核糖體結構和生物發生(207 個),翻譯后修飾、蛋白質轉換、伴侶蛋白(192 個),細胞內運輸、分泌和囊泡運輸(168 個),翻譯(164 個),氨基酸的轉運和代謝(88 個),復制、重組和修復(78 個)等。共有2 738 個蛋白獲得KEGG通路注釋,其中重要通路有核糖體(108 個),RNA轉運(80 個),酵母細胞周期(78 個),氧化磷酸化(76 個),內質網中的蛋白質加工(74 個)等(圖2E)。的MDA含量和蛋白質羰基含量均顯著下降,與正常培養下的含量無顯著差異,特別是當NaCl濃度為300 mmol/L時,MDA和蛋白質羰基含量最低。

圖2 庫德畢赤酵母在不同脅迫下全蛋白功能注釋Fig.2 Functional annotations of whole proteins in P.kudriavzevii under various stresses
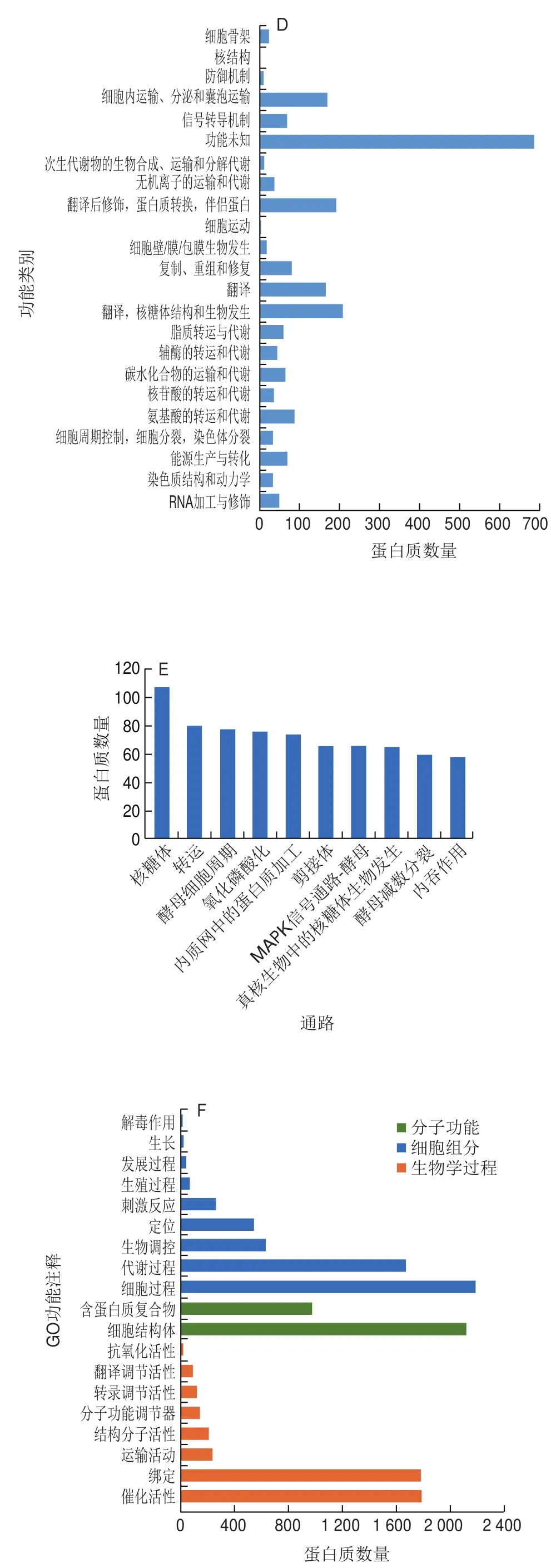
共有3 382 個蛋白獲得GO功能注釋,主要分為生物學過程、細胞組分、分子功能3 大類,其中生物學過程類別主要涉及細胞過程、代謝過程、生物調控、定位等功能;細胞組分類別主要涉及細胞結構體和含蛋白質復合物;分子功能類別主要涉及催化活性、綁定等功能(圖2F)。共有3 320 個蛋白獲得功能域注釋,其中注釋到最多個蛋白質的功能域為WD40(178 個)和Pkinase(83 個)(圖2G)。所有蛋白質(3 875 個)都得到了亞細胞定位預測,這些蛋白主要分布在細胞質(2 206 個)、細胞核(761 個)、線粒體(327 個)和高爾基體(228 個)(圖2H)。
2.3 鹽脅迫對酵母菌熱脅迫蛋白表達的影響
以|logFC|>0.58且<0.05作為DEPs的篩選閾值,結果如圖3A所示。T45S100 vs T45的DEPs較少,只有14 個,與T45組相比,T45S100組上調DEPs有8 個,下調DEPs有6 個。T45S300 vs T45的DEPs較多,共130 個,T45S300組上調DEPs有68 個,下調DEPs有62 個。T45S100 vs T45和T45 vs T45S300兩個比較組共有的DEPs有12 個,T45 vs T45S100和T45S300 vs T45獨有的DEPs分別有2 個和118 個(圖3B)。對DEPs進行GO、COG 、KEGG功能注釋。全蛋白功能注釋結果一致,T45S100 vs T45和T45S300 vs T45兩個比較組DEPs的GO功能主要涉及細胞過程(3和34)、代謝過程(3和32)、細胞結構體(4和32)、含蛋白質復合物(0和10)、催化活性(3和37)和綁定(2和30)(圖3C)。將兩個比較組DEPs與COG數據庫進行比對,結果發現,除了功能未知(2和22)外,COG功能主要集中在碳水化合物運輸和代謝(0和11),翻譯后修飾、蛋白質轉換、伴侶蛋白(1和6),氨基酸運輸與代謝(0和3)等(圖3D)。兩個比較組DEPs的KEGG代謝通路主要集中在物質的運輸與代謝中,其中T45S300 vs T45的DEPs較多,主要包括碳水化合物代謝(1和5)、氨基酸代謝(3和11)、脂質代謝(0和10)、核苷酸代謝(0和6)、能量代謝(0和8)和膜轉運(2和25)(圖3E)。


圖3 T45S100 vs T45和T45S300 vs T45比較組DEPs篩選及功能注釋Fig.3 Screening and functional annotations of DEPs in T45S100 vs T45 and T45S300 vs T45
基于KEGG代謝通路注釋,對上調和下調的DEPs進行分析,結果圖4所示。上調和下調的DEPs在T45S100組和T45S300組中基本表現出一致性,而且各個DEPs表達量的變化與鹽脅迫濃度密切相關:大部分上調的DEPs在T45S300組的上調倍數大于T45S100組,而下調的DEPs在T45S300組的下調程度大于T45S100組。在碳水化合物代謝中,高鹽脅迫(T45S300組)顯著提高了糖酵解/糖異生代謝通路中蛋白的表達,其中T45S300組上調DEPs有12 個,下調DEPs只有2 個;淀粉和蔗糖代謝(葡萄糖-6-磷酸異構酶、己糖激酶(hexokinase,HK)、HK2)和磷酸戊糖途徑(乙醇脫氫酶(alcohol dehydrogenase,ADHP)4、果糖二磷酸醛縮酶(fructose-bisphosphate aldolase,FBA))代謝通路中的DEPs全部上調。在氨基酸代謝中,鹽脅迫顯著降低了與氨基酸降解相關的酶的表達,如色氨酸、組氨酸、精氨酸等。在能量代謝中,涉及到氧化磷酸化(V型質子ATP酶亞基c’(V-type proton ATPase subunit c’,ATP6L)、細胞色素c氧化酶亞基6A(cytochrome c oxidase subunit 6A,COX6A))和硫代謝(蛋白質APA1)代謝通路的DEPs在T45S300組全部上調。在脂質代謝中,DEPs主要參與甘油磷脂代謝、甘油脂代謝、類固醇生物合成,其中涉及到甘油磷脂合成(二酰基甘油焦磷酸磷酸酶1)和類固醇合成(甲基固醇單加氧酶)的DEPs在T45S300組中全部上調。在膜運輸中,DEPs主要為糖、鈉離子和多肽的轉運蛋白,其中涉及到糖(糖轉運蛋白STL1)和離子轉運(鈉轉運ATP酶5(sodium transport ATPase 5,ENA))的DEPs在鹽脅迫后表達量顯著提高。

圖4 基于KEGG通路注釋的T45S100 vs T45和T45S300 vs T45比較組中DEPs熱圖Fig.4 Heat map of DEPs in T45S100 vs T45 and T45S300 vs T45 based on KEGG pathway annotation
2.4 鹽脅迫提高庫德畢赤酵母耐熱性機制
酵母菌在高溫條件下的乙醇發酵能力與其耐熱性密切相關,耐熱性的提高有助于提升其在高溫條件下的乙醇產量。本研究發現,鹽脅迫對庫德畢赤酵母熱脅迫產生交互保護作用,顯著提高其耐熱性。進一步采用TMT定量蛋白質組學技術研究了庫德畢赤酵母在單獨熱脅迫組和熱-鹽共脅迫組之間的蛋白質組差異,篩選了關鍵的DEPs,結果如表1所示。根據不同脅迫條件下蛋白質組的比較結果,在蛋白質組水平初步揭示了鹽脅迫對庫德畢赤酵母熱脅迫交互保護機制(圖5)。

圖5 鹽脅迫提高庫德畢赤酵母耐熱性機制示意圖Fig.5 Schematic representation of the proposed mechanisms of improved thermotolerance by salt stress in P.kudriavzevii
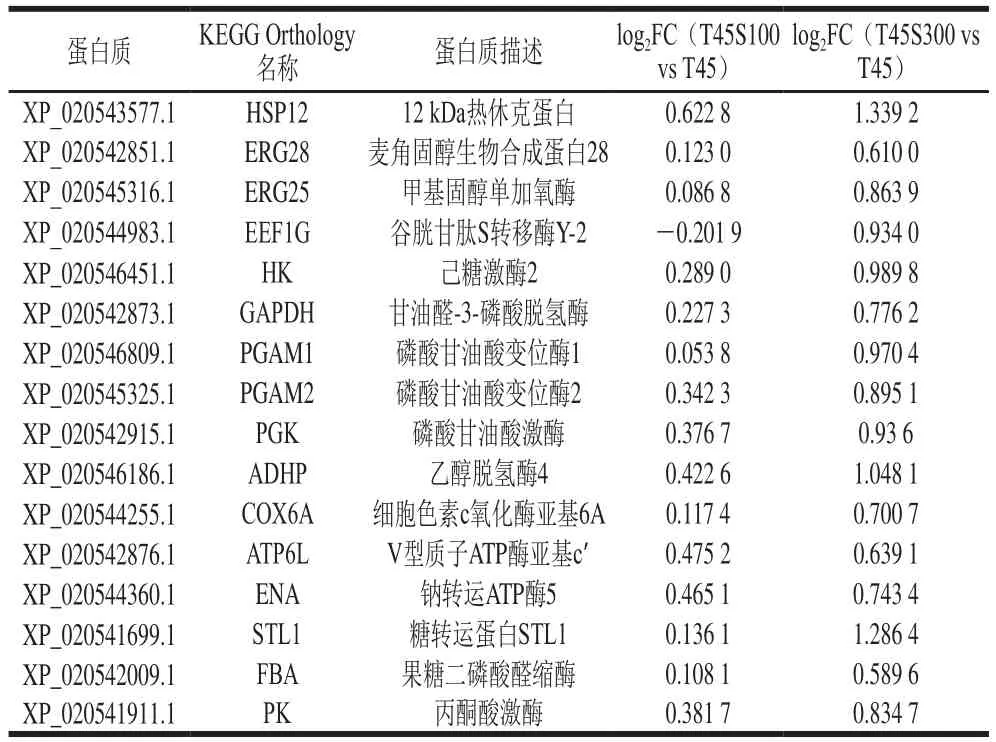
表1 鹽脅迫下與庫德畢赤酵母熱耐性提高相關的關鍵DEPsTable 1 Key DEPs related to the improved thermotolerance of P.kudriavzevii by salt stress
熱休克蛋白(heat shock protein,HSP)是一類受到外源刺激產生的具有高度保守性的非特異性保護蛋白。在熱脅迫下,HSP主要作為分子伴侶,預防和修復變性或錯誤折疊的蛋白質。小熱休克蛋白(small HSP,sHSP)的種類繁多,是HSP家族中最多樣化的群體。作為sHSP家族的一員,HSP12在鹽脅迫后其基因表達量顯著增加,對庫德畢赤酵母鎘脅迫產生了明顯的交互保護作用。研究比較蛋白質組結果顯示,鹽脅迫顯著增強了熱脅迫下庫德畢赤酵母HSP12的表達水平,特別是T45S300組,其細胞內HSP12含量較單獨熱脅迫下(T45組)提高了1.5 倍,有利于酵母菌維持熱脅迫下細胞內蛋白質的穩定,增強其耐熱性。麥角甾醇作為一種重要的酵母甾醇,參與維持細胞膜的流動性、完整性和通透性。有研究表明,麥角甾醇產量的增加明顯提高了的耐熱性。本研究發現,與麥角固醇合成相關的酶,包括麥角固醇生物合成蛋白28(ergosterol biosynthetic protein 28,ERG28)和甲基固醇單加氧酶(ERG25)在鹽脅迫后顯著增強,對于穩定熱脅迫下庫德畢赤酵母細胞膜的正常生理功能,提高其耐熱性具有重要作用。
熱脅迫可誘導細胞內超氧陰離子自由基、HO等ROS的產生,而過量ROS積累將引起細胞內脂質、蛋白質等生物大分子的氧化損傷。本研究也觀察到了相似的結果,熱脅迫下庫德畢赤酵母細胞內MDA含量和蛋白質羰基含量明顯增加,表明細胞脂質和蛋白質在熱脅迫下產生了明顯的氧化損傷,導致生物量顯著下降(圖1)。鹽脅迫后,庫德畢赤酵母在熱脅迫下的脂質和蛋白質氧化損傷得到了顯著改善。谷胱甘肽S-轉移酶(glutathione S-transferase,GST)是催化親電子基團與還原型谷胱甘肽的巰基偶聯重要的酶,在細胞內起到異源物質代謝和解毒作用。GST具有類似過氧化物酶的作用,能夠清除細胞內ROS。前期研究發現,鹽脅迫能顯著提高庫德畢赤酵母在鎘脅迫下GST基因的表達。本研究通過定量蛋白質組學技術也發現了類似的結果,鹽脅迫(300 mmol/L NaCl)顯著增強了庫德畢赤酵母熱脅迫下GST的表達水平,這對于降低熱脅迫誘導的氧化損傷、提高酵母的耐熱性發揮著重要作用。
碳水化合物代謝和能量代謝為酵母的正常生長和繁殖提供必要的能量。本研究中,鹽脅迫顯著提高多種碳水化合物代謝通路相關酶的表達,其中糖酵解相關的酶有HK、甘油醛-3-磷酸脫氫酶(glyceraldehyde-3-phosphate dehydrogenase,GAPDH)、磷酸甘油酸變位酶(phosphoglycerate mutase,PGAM)、磷酸甘油酸激酶(phosphoglycerate kinase,PGK)和丙酮酸激酶(pyruvate kinase,PK);參與磷酸戊糖途徑的酶有FBA和ADHP。這些酶表達量的提高促進了庫德畢赤酵母在熱脅迫下產生更多的NADH,這些NADH隨后進入線粒體的氧化磷酸化過程中用以進行ATP合成。與此同時,能量代謝途徑中涉及氧化磷酸化的多種關鍵酶在鹽脅迫后表達量顯著增加,主要包括COX6A和ATP6L,有利于庫德畢赤酵母在熱脅迫下的ATP合成。有研究表明,酵母細胞在熱脅迫下ATP水平明顯降低,ATP合成的改善有利于酵母耐熱性的提高。因此,本研究中鹽脅迫后糖酵解途徑、磷酸戊糖途徑、氧化磷酸化途徑等多種酶表達量的提高,有利于細胞產生更多的ATP,為熱脅迫下細胞的正常新陳代謝提供必要的能量,在酵母耐熱性提高過程中發揮著重要作用。
此外,與膜轉運相關的蛋白,ENA、糖轉運蛋白STL1表達水平在鹽脅迫后顯著提高。ENA具有結合ATP和ATP偶聯陽離子跨膜轉運體活性的功能,在維持細胞內Na的生理濃度方面起著關鍵作用,在正常生長條件下ENA在細胞中處于相對較低的水平。本研究中,隨著鹽脅迫濃度增加,ENA表達量逐漸上升,這可以防止細胞內Na含量過高,降低鹽脅迫對細胞造成的不利影響。STL1主要酵母細胞碳水化合物和甘油的運輸。前期研究發現,鹽脅迫能夠顯著提高庫德畢赤酵母發酵液中的甘油含量。STL1表達水平的提高對于維持酵母細胞內外甘油平衡、保障熱脅迫下細胞正常生理功能有重要意義。此外,前期的研究表明,鹽脅迫主要通過提高庫德畢赤酵母的耐熱性提高其在高溫條件下的乙醇產量。本研究發現,鹽脅迫后庫德畢赤酵母細胞內ADHP的表達量顯著提高,有利于將糖酵解產物乙醛進一步催化生成乙醇。本研究表明,鹽脅迫后ADHP表達量的提升是庫德畢赤酵母在高溫條件下乙醇產量提升的另一個重要原因。
3 結 論
鹽脅迫(100~500 mmol/L NaCl)能夠對庫德畢赤酵母熱脅迫產生交互保護作用,顯著提高其耐熱性。基于TMT技術的定量蛋白質組學研究發現,鹽脅迫特別是高鹽脅迫顯著影響庫德畢赤酵母在熱脅迫下的蛋白質表達。鹽脅迫顯著增強了庫德畢赤酵母細胞內HSP12,以及ERG28、ERG25等麥角固醇合成相關酶的表達,有利于維持熱脅迫下細胞內生物大分子結構和功能的穩定。鹽脅迫顯著抑制熱誘導的MDA和蛋白質羰基的產生,這可能與GST大量表達對ROS的清除作用有關。同時,熱脅迫下庫德畢赤酵母細胞內與碳水化合物代謝和能量代謝的多種酶(HK、GAPDH、PGAM1、PGAM2、PGK、ADHP、COX6A、ATP6L)在鹽脅迫后表達量顯著提高,有利于細胞內ATP合成,維持熱脅迫下細胞的正常新陳代謝。本研究不僅為耐熱酵母菌的基因工程改造提供重要技術基礎,同時還為其他耐性微生物交互保護的研究提供重要參考。
猜你喜歡
發明與創新·初中生(2024年6期)2024-06-16 00:00:00
昆明醫科大學學報(2022年1期)2022-02-28 07:43:38
中國民間療法(2021年8期)2021-07-22 05:53:28
軍事文摘·科學少年(2021年1期)2021-02-04 08:03:45
當代水產(2019年3期)2019-05-14 05:43:24
天然產物研究與開發(2018年9期)2018-10-08 03:25:32
煙草科技(2015年8期)2015-12-20 08:27:04
藥學與臨床研究(2015年4期)2015-06-05 11:35:51
閱讀與作文(小學低年級版)(2015年8期)2015-05-30 10:48:04
食品工業科技(2014年9期)2014-03-11 18:15:31
