不同沙障材料對固沙林地面節肢動物群落結構分布的影響
2022-08-04 09:10:06馮永宏茍琪劉任濤羅雅曦常海濤張靜蔣嘉瑜
生態科學 2022年5期
馮永宏, 茍琪, 劉任濤,*, 羅雅曦, 常海濤, 張靜, 蔣嘉瑜
不同沙障材料對固沙林地面節肢動物群落結構分布的影響
馮永宏1, 茍琪2,3, 劉任濤2,3,*, 羅雅曦2, 常海濤2, 張靜1, 蔣嘉瑜2,3
1. 寧夏大學農學院, 銀川 750021 2. 寧夏大學西北土地退化與生態恢復國家重點實驗室培育基地, 銀川 750021 3. 寧夏大學生態環境學院, 銀川 750021
為了明確沙障材料差異性對固沙林地面節肢動物群落結構的影響規律, 以騰格里沙漠東南緣2013年布設的麥草、尼龍網兩種沙障模式下的固沙林地為研究樣地, 于2018年春、夏、秋季, 研究了沙丘底部和頂部不同沙障模式下的固沙林地地面節肢動物群落組成及多樣性分布特征。結果發現: (1)沙丘底部, 2種沙障林地地面節肢動物優勢類群除了均包括蠼螋科、步甲科、葉甲科外, 麥草方格林地還包括擬步甲科和鞘翅目幼蟲2個類群, 而尼龍網方格林地還包括鱗翅目幼蟲1個類群; 但在沙丘頂部, 2種沙障林地地面節肢動物優勢類群均為蠼螋科、步甲科、葉甲科和擬步甲科。(2)沙丘底部和頂部, 優勢類群步甲科、葉甲科個體數在2種沙障林地間均無顯著差異; 但優勢類群擬步甲科個體數表現為麥草方格林地顯著高于尼龍網方格。在沙丘底部, 蠼螋科個體數表現為2種沙障林地間無顯著差異; 而在頂部, 蠼螋科個體數表現為麥草方格林地顯著高于尼龍網方格。(3)沙丘底部和頂部, 地面節肢動物個體數、類群數均表現為麥草方格林地顯著高于尼龍網方格, 但Shannon指數、優勢度指數2種沙障林地間均無顯著差異。(4)RDA和偏RDA分析結果表明, 花棒冠幅與高度、油蒿冠幅是不同微生境中地面節肢動物個體數分布產生差異的主要因素。(5)研究表明, 沙障材料對地面節肢動物群落組成、個體數和類群數分布均產生顯著影響, 而且地面節肢動物群落組成分布亦受到沙丘微地形的影響, 但沙障材料對地面節肢動物多樣性分布的影響較小。
騰格里沙漠; 沙障材料; 沙丘微地形; 地面節肢動物; 群落多樣性
0 前言
沙漠化是全球性嚴重的環境問題之一, 全球現有約4560萬km2的沙漠化土地, 并且以每年5—7萬km2的速度增長[1]。沙漠化不僅造成嚴重的生態系統失衡, 而且給我國農業生產和人民生活帶來嚴重影響[2]。因此, 需要采取合理的人工沙漠化防治措施來改善生態環境。在騰格里沙漠東南緣, 進行固沙林營造并扎設草方格, 可以有效改善土壤理化性質, 改良土壤結構, 促進退化土壤修復[3–5], 已被認為是成功的生態恢復措施而得到了大面積推廣。然而, 不同沙障材料由于其本身特性及價格成本的差異性, 往往會影響沙漠固沙造林工程的實施效果和固沙造林后植被恢復過程[6]。并且, 地面節肢動物及其多樣性是固沙林地生態系統中生物多樣性的重要組成部分, 在物質轉換、凋落物分解、土壤發育和成熟等生態功能過程中扮演著重要的角色[7], 能夠直接反映固沙林地生態恢復過程和進程。因此, 研究麥草、尼龍網等不同沙障模式下的固沙林地地面節肢動物群落組成及多樣性分布特征, 對于干旱風沙區生物多樣性保育、人工植被建設以及沙漠化防治均具有重要的理論與實踐意義。
目前, 學者們已從不同角度對不同材料沙障的固沙效應做了較多研究[8–11]。孫濤等[11]在民勤發現尼龍網沙障隨時間的延長仍具有較好的固沙功能, 而麥草沙障的固沙功能隨時間的增加逐漸降低。王雨浩[12]在騰格里沙漠的東南緣發現在沙障規格相同的情況下, 麥草沙障的防風固沙效益要優于聚乳酸纖維沙障。研究表明, 不同材料草方格固沙效應的差異性與沙障本身特性及應用成本密切相關。例如, 尼龍網沙障作為一種新型的防護材料, 具有造價相對低廉, 抗老化, 抗風沙, 特別是在一些風沙災害比較嚴重的地區, 尼龍網防護能快速達到立竿見影的效果[3]; 但尼龍網方格更是一種物理性防沙技術, 且不易分解而造成塑料污染。而草方格沙障采用的麥草為生物材料, 既可以作為枯落物, 其分解可以為沙漠微生物提供養分來源, 又具有保水性能[5]; 但缺點是分解較快, 且分解之后的固沙效果降低。因此, 不同材料沙障在固沙效應方面均存在自己的優勢和劣勢, 將會對固沙林地的土壤-植被系統恢復進程產生不同的影響。但是, 不同沙障材料對固沙林地地面節肢動物群落結構分布的影響規律, 尚不清楚。并且, 以往研究多針對地形條件相對平坦的沙地中某一種固沙材料的研究[13]。例如, 屈建軍等[8]對半隱蔽格狀沙障的綜合防護效益研究中, 發現同一規格的格狀沙障往往由于設置的地形部位不同,其防護作用可能產生很大的差異。已有研究表明, 微地形變化可以對沙質荒漠擬步甲科昆蟲分布產生深刻影響[13]。但是, 關于草方格固沙林地中地面節肢動物多樣性對較大起伏地形的適應性是否存在差異研究, 報道較少。
鑒于此, 在騰格里沙漠東南緣選取麥草、尼龍網沙障模式下的人工固沙林地為研究對象, 在沙丘底部和頂部生境中布設調查樣點, 采用陷阱誘捕法對地面節肢動物群落組成、多樣性分布特征進行調查, 結合環境分析, 旨在闡明不同沙障材料對固沙林地地面節肢動物群落結構分布的影響規律, 豐富沙漠生態系統土壤動物生態學的研究內容, 為人工林建設、采取有效的防沙治沙措施及退化生態系統恢復提供依據。
1 材料與方法
1.1 研究區概況
研究區位于寧夏回族自治區中衛市騰格里沙漠東南緣(37°25′N, 104°45′E), 海拔1350—1450 m, 屬于典型的溫帶荒漠化草原向草原化荒漠的過渡地區。年平均氣溫10.0 ℃, 最低溫-25.1 ℃, 最高溫38.1 ℃。年平均降水量186.2 mm, 且分布不均勻, 降水主要集中于7—9月, 全年日照時間3264 h。全年主要西北風居多, 年均風速2.9 m·s–1。土壤以細沙粒為主, 以風沙土及灰棕荒漠土分布最廣。該區地形起伏變化較大, 景觀主要是高大且密集的網格狀沙丘, 沙丘從西北向東南傾斜, 且呈階梯狀分布。
在世界銀行貸款項目“寧夏黃河東岸防沙治沙林業生態工程”中衛人工固沙植被項目的資助下, 于2013年開始新一輪草方格人工植被固沙工程建設。麥草沙障是用長條狀麥草捆設置成1 m×1 m的網格狀, 地面以上15 cm, 地面埋入沙內深度15 cm; 尼龍網沙障是用尼龍網材料設置成1 m×1 m的網格狀, 地面以上15cm, 地面埋入沙內深度15cm。種植規格為: 1 m×3 m和1 m×1 m(株距×行距)分別種植花棒和檸條, 并在沙障內撒播油蒿種子。經過近5年的固沙恢復, 沙丘底部麥草方格內花棒、檸條和油蒿密度分別為0.14株·m–2、0.14株·m–2、11.6株·m–2, 尼龍網方格內花棒、檸條和油蒿密度分別為0.19株·m–2、0.19株·m–2、7.4株·m–2; 沙丘頂部麥草方格內花棒、檸條和油蒿密度分別為0.11株·m–2、0.08株·m–2、12.4株·m–2, 尼龍網方格內花棒、檸條和油蒿密度分別為0.61株·m–2、0.06株·m–2、2.8株·m–2。灌叢植物種包括: 檸條()、油蒿()、花棒()、沙拐棗()和沙木蓼()等旱生灌木。
1.2 試驗設計與取樣調查
1.2.1 試驗設計
于2018年春季, 在騰格里沙漠東南緣2013年開始建設的人工灌叢固沙林地中, 分別選擇麥草、尼龍網兩種沙障模式下的林地為研究樣地, 每種沙障模式下的林地設置5個重復樣區, 每個樣地面積均為900 m2, 間距為50 m左右。在每種樣地, 選擇丘頂和丘底分別布設調查樣點4個, 共布設2沙丘微生境×2沙障×5重復×4個樣點=80樣點。
1.2.2 地面節肢動物取樣調查
分別于2018年春季(4月)、夏季(7月)、秋季(9月), 在每個調查樣點, 采用國際通用的陷阱誘捕法采集地面節肢動物[14]。具體方法為: 在每個調查樣點的中心位置將塑料杯(上下直徑分別為14 cm和7 cm, 高度為10 cm)埋入土中, 杯口略低于地面, 同時在其內加入少量甘油和體積分數為3%的福爾馬林溶液, 以增加誘捕的有效性。每次持續時間均為12 d(每隔3 d檢查收集1次, 避免采集標本的損失)。試驗結束后將收集到的地面節肢動物標本帶回實驗室進行分類鑒定。
地面節肢動物標本分類鑒定主要依據《寧夏賀蘭山昆蟲》[15]、《中國土壤動物檢索圖鑒》[16]和《昆蟲分類》[17]等。根據不同地面節肢動物類群在群落總個體數中所占比例來劃分等級, 其劃分標準為優勢類群(占群落總個體數的10%以上)、常見類群(1%—10%)和稀有類群(<1%)[18]。
1.2.3 植被調查、土壤樣品采集與分析
在每個調查樣點布設5 m×5 m的樣方, 測定檸條和花棒的高度(cm)、冠幅(m2)指標。同時, 在5 m× 5 m的樣方內, 按照“M”型設置5個1 m×1 m的草方格, 調査沙障內油蒿的高度(cm)、冠幅(m2)指標。由于灌叢植被同一年中不同季節變化不大, 故只調查1次。
在每個調查樣點內, 利用5點取樣法采集混合土樣裝入自封袋(取樣深度為0—10 cm), 放置在保溫箱中, 土壤樣品帶回實驗室, 取混合土樣中的1/4用于土壤含水量的測定, 剩余3/4土樣經自然風干后, 過2 mm土壤篩, 以去除樹根、葉片等雜物, 以供土壤理化性質測定使用。具體測定指標包括: 土壤溫度、容重、含水量、pH、電導率、粒徑組成、有機碳和全氮共8個指標。
土壤溫度(℃)采用地溫儀測定。土壤容重(采用環刀法在每個調查樣點取原狀土樣1個; g·cm–3)和土壤含水量(%)均采用烘干稱重法測定。土壤pH和土壤電導率(μS·m–1)均采用1:5土水比浸提后, 分別使用P4多功能儀(Muiti-line P4 Universal Meter, WTW公司, Germany)和便攜式電導率儀進行測定[19]。土壤粒徑組成采用Mastersizer3000激光衍射粒度分析儀進行測定。根據美國農業部(USDA)制土壤質地分級標準將本試驗土壤樣品質地劃分為: 細沙粒(100—250 μm)、極細沙粒(50—100 μm)和粉粒(2—50 μm)3個等級[1]。土壤有機碳(%)和全氮(%)分別采用重鉻酸鉀氧化外加熱法和凱氏定氮法(意大利DK6, UDK140分析儀)測定[20]。
1.3 數據處理
分別將春、夏、秋季每個樣地采樣點數據進行整理, 用于統計地面節肢動物個體數(只·陷阱–1)、類群數以及計算群落多樣性, 包括Shannon指數和優勢度指數。然后, 利用平均值法將3個季的數據進行平均。
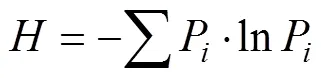
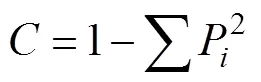
式中,P表示類群的個體數在總個體中的比例, 即P=/。
所有數據采用SPSS 25.0軟件進行統計分析。通過獨立樣本檢驗分析同一位置不同沙障材料調查樣點間地面節肢動物個體數、類群數、多樣性指數的差異性。統計在<0.05水平上存在顯著差異。應用Origin 2018進行作圖。
利用已建立的4種不同生境地面節肢動物個體數據矩陣, 采用DCA分析(Detrended correspondence analysis, DCA)確定排序軸的梯度長度(Lengths of gradient, LGA)。依據LGA值大小選擇適宜的排序分析方法, 其基本準則是: 當LGA<4時, 宜采用線性模型排序; 當LGA>4時, 宜采用非線性模型排序[21]。由于本文地面節肢動物群落DCA計算出的LGA=1.779<4, 故采用線性模型的冗余分析(Redundancy analysis, RDA)來確定地面節肢動物個體數分布與環境因子間的關系。同時, 采用偏RDA分析(Partial RDA)和蒙特卡洛置換檢驗(Monte Carlo permutation test), 定量評價每個環境因子對地面節肢動物群落分布變化的貢獻率(即獨立解釋量)。在偏RDA分析的基礎上, 選擇對對地面節肢動物群落分布有較高影響的解釋變量, 繪制地面節肢動物類群分布與解釋變量關系的二維排序圖(Biplot)。應用CANOCO4.5進行分析運算。為了保證試驗數據滿足正態分布及減小異常值對分析結果的影響, 對動物個體數量及環境因子數據進行平方根轉換, 對樣方進行中心化和標準化(Center and standardize)并對物種數據進行誤差方差標準化(Standardize by error variance)處理。
2 結果與分析
2.1 環境特征
由表1可以看出, 無論是在沙丘底部還是頂部, 土壤有機碳、pH、含水量、電導率、容重、土壤粉粒和溫度均表現為麥草方格和尼龍網方格間無顯著差異(>0.05)。但是, 在沙丘底部, 僅土壤極細沙粒含量表現為麥草方格顯著高于尼龍網方格(<0.05); 在沙丘頂部, 僅土壤全氮和細沙粒含量表現為麥草方格顯著低于尼龍網方格(<0.05)。
由表2可知, 無論沙丘底部還是頂部, 檸條高度表現為麥草方格顯著低于尼龍網方格(<0.05); 而油蒿高度表現為麥草方格和尼龍網方格間無顯著差異(>0.05)。在沙丘底部, 花棒冠幅表現為麥草方格顯著低于尼龍網方格(<0.05), 而檸條冠幅、花棒高度、油蒿冠幅均表現為麥草方格和尼龍網方格間無顯著差異(>0.05); 在沙丘頂部, 檸條冠幅、油蒿冠幅均表現為麥草方格顯著低于尼龍網方格(<0.05), 花棒高度表現為麥草方格顯著高于尼龍網方格(<0.05), 而花棒冠幅表現為麥草方格和尼龍網方格間無顯著差異(>0.05)。
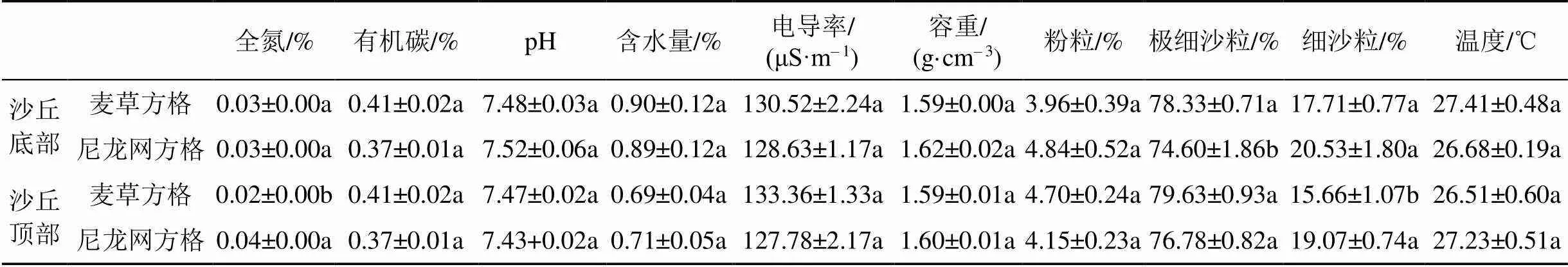
表1 不同材料沙障沙丘底部和頂部土壤理化性質(均值±標準誤)
注: 同列不同字母表示同一沙丘位置不同沙障材料處理間差異顯著(<0.05)。
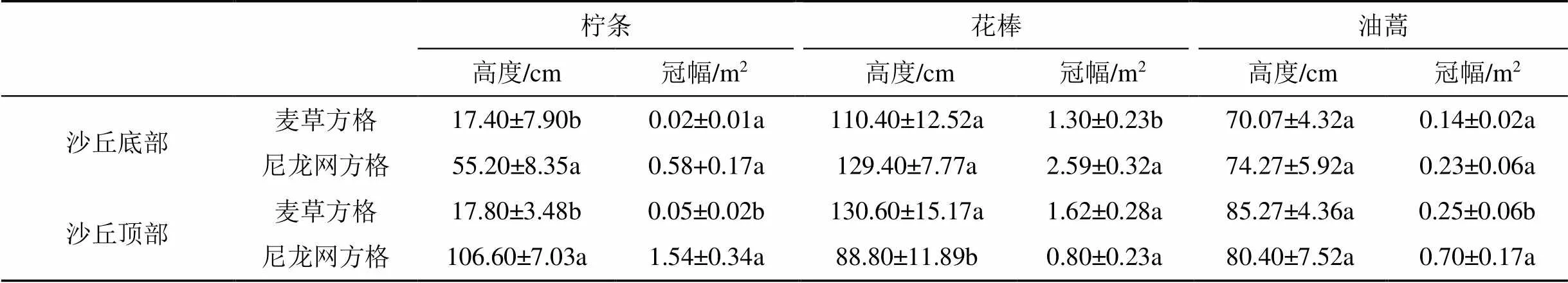
表2 不同材料沙障沙丘底部和頂部地表植被指標(均值±標準誤)
注: 同列不同字母表示同一沙丘位置不同沙障材料處理間差異顯著(<0.05)。
2.2 地面節肢動物群落組成與數量特征
本次調查共計獲得地面節肢動物659只。鑒定隸屬于10目25科27個類群(表3)。其中, 優勢類群為步甲科、擬步甲科、蠼螋科和葉甲科, 其個體數分別占總個體數的30.20%、18.51%、16.24%和13.20%。常見類群為鞘翅目幼蟲、鱗翅目幼蟲、蚱蜢科、金龜甲科和平腹蛛科, 其個體數分別占總個體數的6.53%、3.19%、2.28%、2.12%和1.67%。其余18個類群為稀有類群, 其個體數僅占總個體數的6.07%。
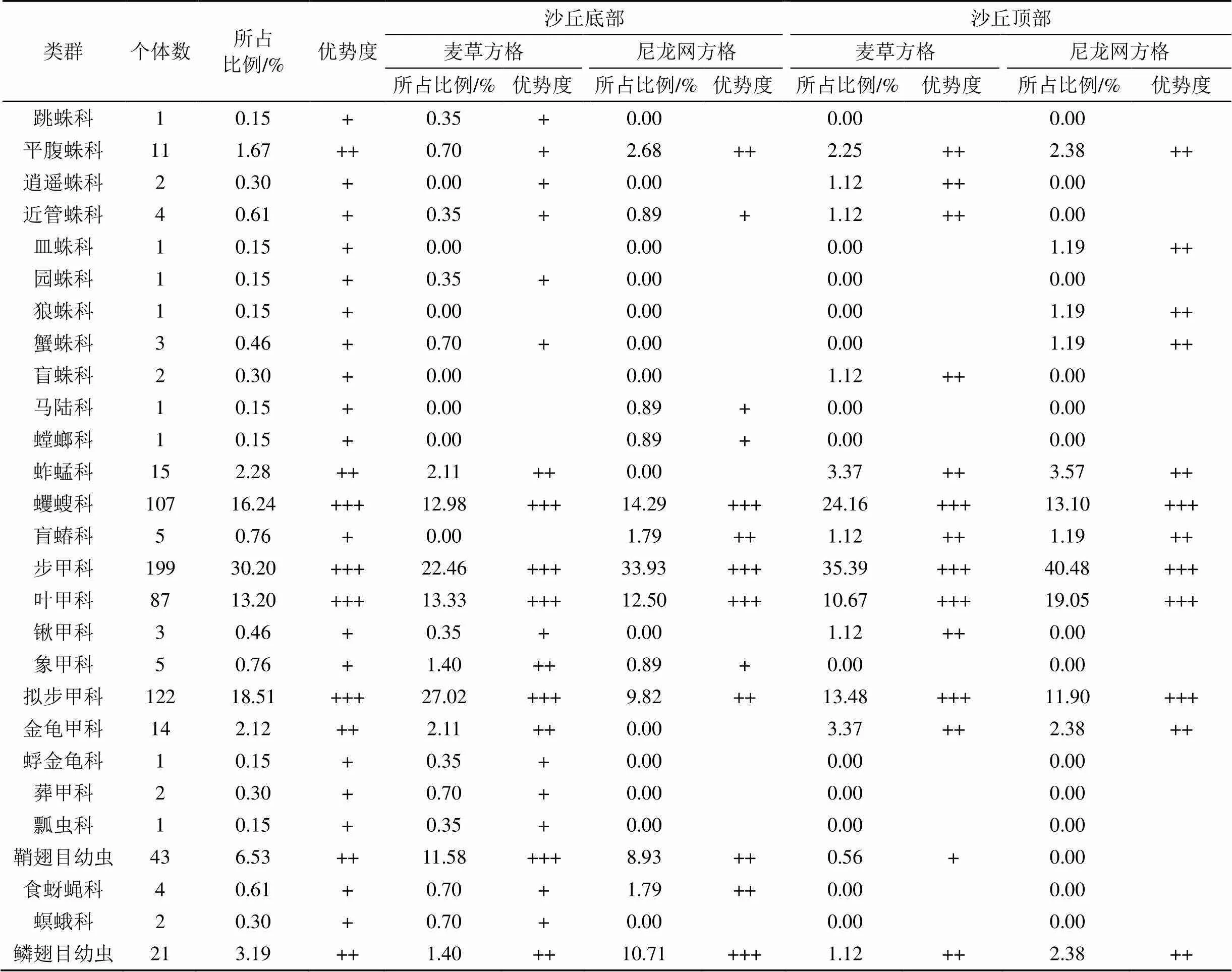
表3 地面節肢動物群落組成及多度分布
注: 個體數占全部捕獲量的10%以上為優勢類群, 用+++表示; 介于1%—10%為常見類群, 用++表示; 介于0.1%—1%為稀有類群, 用+表示。
在沙丘底部, 麥草方格固沙林地內共捕獲地面節肢動物285只、18科、20類群。其中, 優勢類群包括擬步甲科、步甲科、葉甲科、蠼螋科和鞘翅目幼蟲5個類群, 個體數分別占總個體數的27.02%、22、46%、13.33%、12.98%和11.58%; 常見類群包括蚱蜢科、金龜甲科、象甲科和鱗翅目幼蟲4個類群, 個體數分別占總個體數的2.11%、2.11%、1.4%和1.4%; 其余11個類群為稀有類群, 其個體數僅占總個體數的5.61%。尼龍網方格固沙林地內共捕獲地面節肢動物112只、10科、12類群。其中, 優勢類群包括步甲科、蠼螋科、葉甲科和鱗翅目幼蟲4個類群, 個體數分別占總個體數的33.93%、14.29%、12.5%和10.71%; 常見類群包括擬步甲科、鞘翅目幼蟲、平腹蛛科、盲蝽科和食蚜蠅科5個類群, 個體數分別占總個體數的9.82%、8.93%、2.68%、1.79%和1.79%; 其余3個類群為稀有類群, 其個體數僅占總個體數的3.57%。
在沙丘頂部, 麥草方格固沙林地內共捕獲地面節肢動物178只、12科、14類群, 其中, 優勢類群包括步甲科、蠼螋科、擬步甲科和葉甲科4個類群, 個體數分別占總個體數的35.39%、24.16%、13.48%和10.67%; 常見類群包括蚱蜢科、金龜甲科、平腹蛛科、鍬甲科、逍遙蛛科、近管蛛科、盲蛛科、鱗翅目幼蟲和盲蝽科9個類群, 個體數分別占總個體數的3.37%、3.37%、2.25%、1.12%、1.12%、1.12%、1.12%、1.12%和1.12%; 其余1科為稀有類群, 其個體數僅占總個體數的0.56%。尼龍網方格固沙林地內共捕獲地面節肢動物84只、11科、12類群, 其中, 優勢類群包括步甲科、葉甲科、蠼螋科和擬步甲科4個類群, 個體數分別占總個體數的40.48%、19.05%、13.1%和11.9%; 常見類群包括蚱蜢科、金龜甲科、平腹蛛科、鱗翅目幼蟲、皿蛛科、狼蛛科、蟹蛛科和盲蝽科8個類群, 個體數分別占總個體數的3.57%、2.38%、2.38%、2.38%、1.19%、1.19%、1.19%和1.19%; 無稀有類群。
從圖1可以看出, 無論在沙丘底部還是頂部, 步甲科、葉甲科個體數均表現為麥草方格和尼龍網方格間無顯著差異(>0.05), 而擬步甲科個體數均表現為麥草方格顯著高于尼龍網方格(<0.05)。在沙丘底部, 蠼螋科個體數表現為麥草方格和尼龍網方格間無顯著差異(>0.05); 但在沙丘頂部, 蠼螋科個體數表現為麥草方格顯著高于尼龍網方格(<0.05)。

注: 不同字母表示同一沙丘位置不同沙障材料處理間差異顯著(P<0.05)。
Figure 1 The abundance of dominant arthropod groups at dune bottom and top within checkerboard of different materials
2.3 地面節肢動物群落多樣性
從圖2可以看出, 無論在沙丘底部還是頂部, 地面節肢動物個體數、類群數均表現為麥草方格顯著高于尼龍網方格(<0.05); 但地面節肢動物Shannon指數、優勢度指數均表現為麥草方格和尼龍網方格間無顯著差異(>0.05)。
2.4 地面節肢動物個體數分布與環境因子間的相關分析
由表4可知, 步甲科、擬步甲科個體數均與檸條高度和冠幅呈負相關關系(<0.05), 而與土壤總氮、極細沙粒、細沙粒、花棒高度、冠幅、油蒿冠幅之間均無相關性(0.05)。蠼螋科個體數與檸條高度和油蒿冠幅呈負相關關系(<0.05), 而與土壤總氮、極細沙粒、細沙粒、花棒高度、冠幅、檸條冠幅之間均無相關性(0.05)。葉甲科個體數與土壤總氮、極細沙粒、細沙粒、花棒高度、冠幅、檸條高度、冠幅、油蒿冠幅之間均無相關性(0.05)。
對地面節肢動物個體數與環境因子間關系的RDA排序分析表明, 第一典型軸(=2.442,=0.014)和所有典型軸(=1.647,=0.002)在統計學上達到顯著水平, 說明排序分析能夠較好地反映地面節肢動物個體數與環境因子的關系。并且, 前兩個排序軸累積解釋了29.5%(第一排序軸解釋了18.3%和第二排序軸解釋了11.2%)的地面節肢動物群落變異。從圖3中可以看出, 沙丘底部麥草方格和尼龍網方格主要在第二、三象限, 而頂部麥草方格與尼龍網方格主要在第一、四象限。
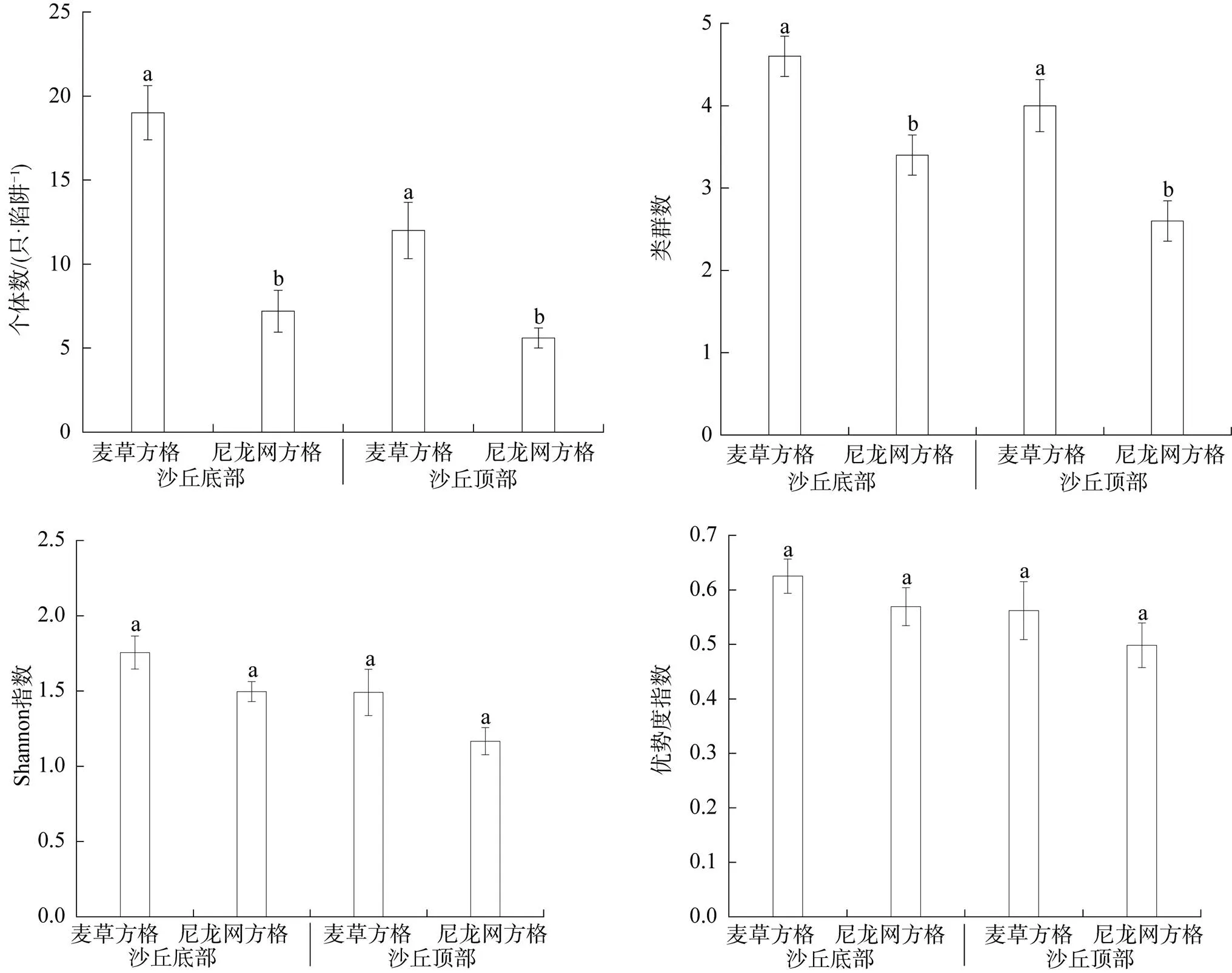
注: 不同字母表示同一沙丘位置不同沙障材料處理間差異顯著(P<0.05)。
Figure 2 The arthropod community index at dune bottom and top within checkerboard of different materials
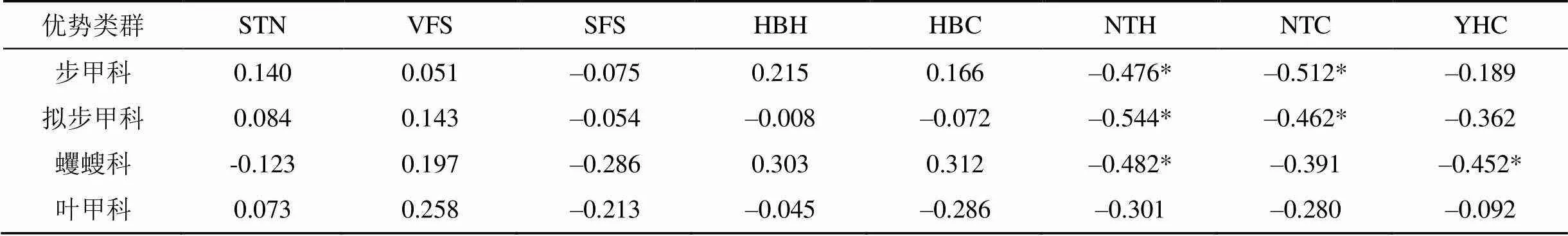
表4 優勢類群個體數與環境因子間的Pearson相關系數
注: STN: 土壤總氮; VFS: 土壤極細沙粒; SFS: 土壤細沙粒; HBH: 花棒高度; HBC: 花棒冠幅; NTH: 檸條高度; NTC: 檸條冠幅; YHC: 油蒿冠幅。*表示在0.05水平(雙側)顯著相關。
偏RDA分析表明(表5), 花棒冠幅與高度、油蒿冠幅對地面節肢動物個體數分布產生顯著影響(<0.05), 貢獻率分別為9%、9%和8%, 而其余環境因子對地面節肢動物個體數影響不顯著(>0.05)。
3 討論
3.1 不同沙障材料對土壤與植被分布的影響
本研究中, 在沙丘頂部, 土壤全氮表現為麥草方格顯著低于尼龍網方格。其原因可能是沙丘頂部尼龍網方格中冠幅較大的檸條根系含有大量的根瘤菌, 可以固定空氣中的氮素, 進而有利于其土壤全氮的積累[22]。土壤細沙粒表現為麥草方格顯著低于尼龍網方格。其原因可能是較大的檸條冠幅和油蒿冠幅可以降低風速[19], 從而導致質量較重的土壤細沙粒沉積在沙丘頂部。土壤極細沙粒含量表現為麥草方格和尼龍網方格間無顯著性差異。原因可能是沙丘頂部風速大, 導致頂部麥草方格和尼龍網方格內極細沙粒均隨風的運移沉積在沙丘其他部位[6], 故2種沙障間極細沙粒無顯著差異。但是在沙丘底部, 土壤極細沙粒含量表現為麥草方格顯著高于尼龍網方格。分析原因可能是在麥草沙障林地中, 由于油蒿密度較大, 覆蓋度較高, 而且油蒿冠層結構致密低矮便于細粒物質的捕獲保存[23], 同時油蒿根系發達并且主要在淺層土壤連續活躍活動[24], 因而其根系分泌物對土壤顆粒也有一定改善作用; 而尼龍網沙障樣地則以花棒和檸條居多, 由于近地層分枝較少且稀疏, 使其易受風蝕影響, 沙層表面較細顆粒相對不易積累[25], 同時, 花棒和檸條為深根系植物[26], 因此對淺層土壤顆粒的影響相對較小。無論在沙丘底部還是頂部, 土壤有機碳、pH、含水量、電導率、容重、粉粒和溫度均表現為麥草方格與尼龍網方格間無顯著差異, 這與羅雅曦[19]在騰格里沙漠的研究結果相似。可能原因是麥草容易腐爛, 經過5年時間導致麥草所起的保持水分和提高養分的作用隨之減弱[27]。因此, 不同沙障材料對土壤有機碳、pH、含水量、電導率、容重、粉粒和溫度的影響較小。
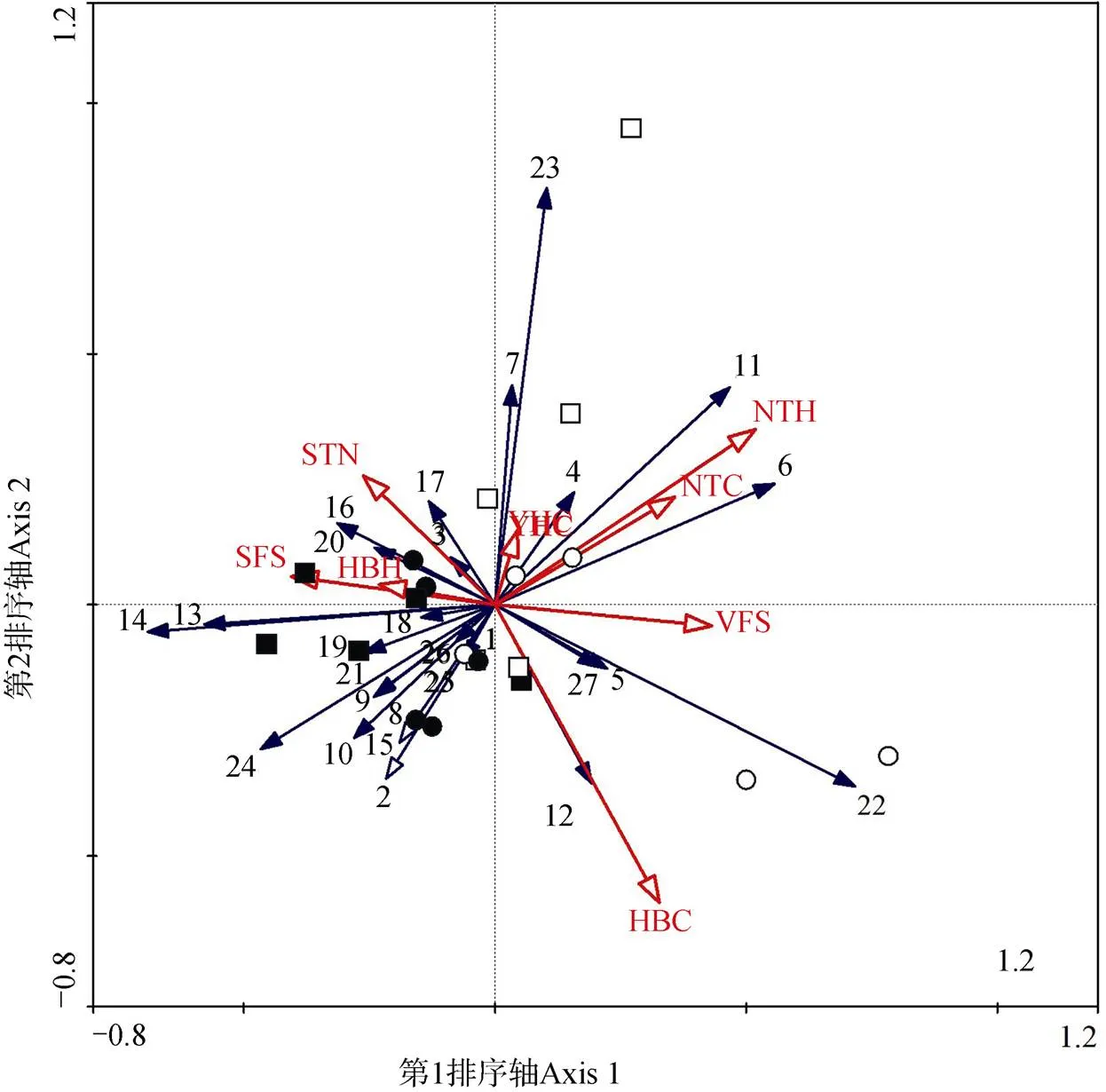
注: HBC: 花棒冠幅; HBH: 花棒高度; NTH: 檸條高度; SFS: 土壤細沙粒; VFS: 土壤極細沙粒; STN: 土壤總氮; NTC: 檸條冠幅; YHC: 油蒿冠幅; 1: 步甲科; 2: 擬步甲科; 3: 螟蛾科; 4: 葉甲科; 5: 金龜甲科; 6: 蚱蜢科; 7: 盲蝽科; 8: 瓢蟲科; 9: 跳蛛科; 10: 象甲科; 11: 平腹蛛科; 12: 鍬甲科; 13: 食蚜蠅科; 14: 鞘翅目幼蟲; 15: 蠼螋科; 16: 近管蛛科; 17: 皿蛛科; 18: 園蛛科; 19: 蟹蛛科; 20: 蜉金龜科; 21: 葬甲科; 22: 逍遙蛛科; 23: 狼蛛科; 24: 鱗翅目幼蟲; 25: 螳螂科; 26: 馬陸科; 27: 盲蛛科; ●=底部麥草方格; ■=底部尼龍網方格; ○=頂部麥草方格; □=頂部尼龍網方格。
Figure 3 RDA two-dimensional diagram of the relationship between ground-active arthropods community distribution and environmental factors
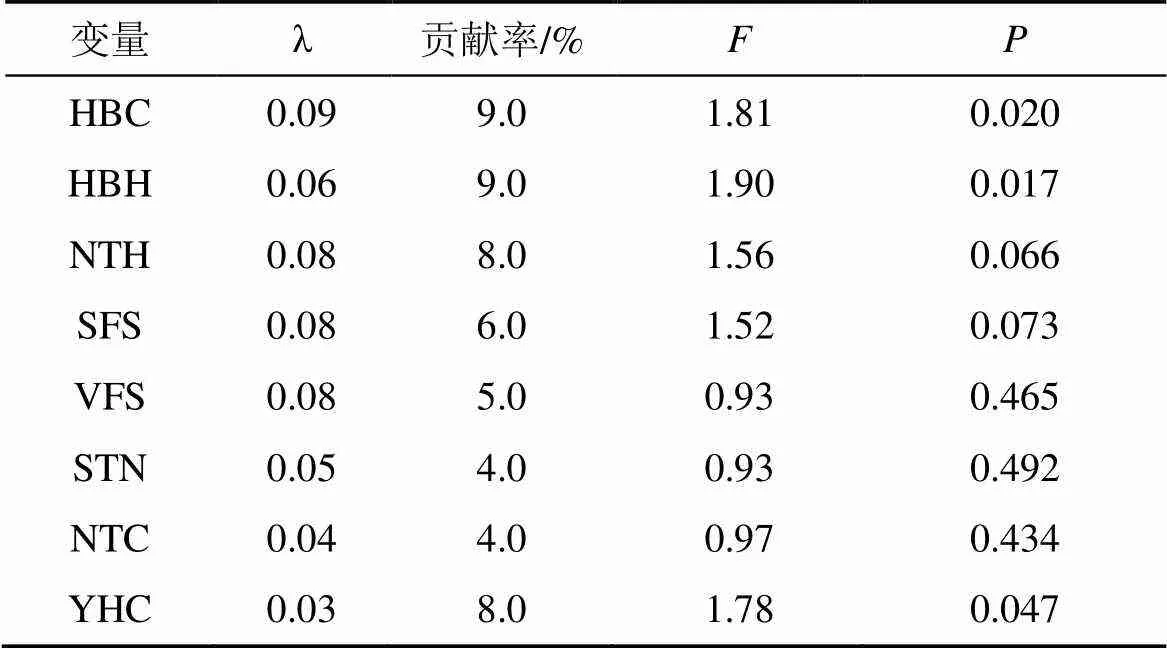
表5 環境因子對地面節肢動物個體數變化的相對貢獻偏RDA分析
注: λ: 邊際效應; HBC: 花棒冠幅; HBH: 花棒高度; NTH: 檸條高度; SFS: 土壤細沙粒; VFS: 土壤極細沙粒; STN: 土壤總氮; NTC: 檸條冠幅; YHC: 油蒿冠幅。
無論在沙丘底部還是頂部, 檸條高度均表現為麥草方格顯著低于尼龍網方格, 表明不同沙障材料對檸條高度存在顯著影響。分析原因可能是麥草壽命短, 容易腐爛, 扎設一定時間后, 某一部位麥草被風蝕裸露后可能會加速鄰近區域麥草的裸露, 麥草所起的保持水分和提高養分的作用隨之減弱[27]。因此, 對于需水量相對較大的檸條難以維持其自身長時間的生長發育及其繁衍, 從而導致檸條高度呈現出麥草方格顯著低于尼龍網方格。在沙丘底部, 檸條和油蒿冠幅均表現為麥草方格和尼龍網方格間無顯著差異。原因可能是沙丘底部土壤含水量均較高[27], 故2種沙障間檸條和油蒿冠幅無顯著差異。但在沙丘頂部, 檸條和油蒿冠幅表現為麥草方格顯著低于尼龍網方格。分析原因可能是麥草沙障抗風沙侵蝕能力減弱[6], 從而導致檸條和油蒿冠幅生長較慢。在沙丘底部, 花棒高度表現為麥草方格與尼龍網方格間無顯著差異, 而在沙丘頂部表現為麥草方格顯著高于尼龍網方格。其原因可能是沙丘底部麥草方格和尼龍網方格內土壤含水量均較高[28–29], 故花棒高度無顯著差異; 而在沙丘頂部, 麥草沙障可以作為枯落物, 其分解可以為沙漠微生物提供養分來源[5], 故麥草沙障對花棒高度影響較尼龍網沙障大。
3.2 不同沙障材料對地面節肢動物群落組成與多樣性分布的影響
在騰格里沙漠生態系統中, 隨沙障材料和沙丘微地形變化、土壤理化性質和植被改變的同時, 地面節肢動物群落組成與結構分布也隨之發生顯著變化。通過比較不同沙障材料林地間地面節肢動物優勢類群的組成, 發現在沙丘底部, 優勢類群除了共有步甲科、蠼螋科、葉甲科3個類群外, 麥草方格固沙林地還包括擬步甲科和鞘翅目幼蟲2個類群, 而尼龍網方格固沙林地內優勢類群則還包括鱗翅目幼蟲1個類群。說明麥草方格固沙林地更適合擬步甲科和鞘翅目幼蟲生存, 而尼龍網方格固沙林地更適合鱗翅目幼蟲生活。分析原因可能是沙丘底部麥草方格固沙林地內檸條高度顯著低于尼龍網方格, 而檸條、花棒密度較低, 導致地面阻力較小, 有利于擬步甲科個體的移動和活動[13], 進而導致擬步甲科個體數分布較多, 故在沙丘底部擬步甲科個體數表現為麥草方格顯著高于尼龍網方格, 這與婁巧哲等[30]在古爾班通沙漠的研究結果類似。同時, 麥草沙障可以作為枯落物, 其分解可以為沙漠鞘翅目幼蟲提供食物來源[5], 故麥草方格固沙林地有利于擬步甲科和鞘翅目幼蟲生存。但與此相反的是, 尼龍網方格林地檸條高度較大, 為鱗翅目幼蟲取食提供了有利的食物資源。鱗翅目幼蟲常取食葉片包括卷葉、綴葉、結鞘、吐絲結網或鉆入植物組織取食[31], 故尼龍網方格固沙林地更適合鱗翅目幼蟲生存。在沙丘頂部, 擬步甲科則是2種沙障材料林地的共同優勢類群, 說明擬步甲科個體數更適應沙丘頂部這種地面阻力較小的特殊環境[13], 進而導致擬步甲科成為沙丘頂部2種沙障材料林地的共有優勢類群。由于沙丘頂部麥草方格內油蒿冠幅顯著低于尼龍網方格, 且麥草方格內油蒿生長稀疏, 從而導致地面阻力較小, 有利于擬步甲科個體的移動和活動[13]。故擬步甲科個體數表現為麥草方格顯著高于尼龍網方格。
本研究發現, 無論在沙丘底部還是頂部, 步甲科、葉甲科個體數均表現為麥草方格和尼龍網方格間無顯著差異, 這與2種沙障材料林地間土壤含水量、電導率、容重、pH、粉粒、地溫無顯著差異有關。分析原因可能是地面節肢動物的個體數分布特征受到土壤含水量、電導率、容重、pH、粉粒、地溫等環境因子的影響較大, 而受到土壤養分含量的影響較小[32]。在沙丘底部, 蠼螋科個體數表現為麥草方格和尼龍網方格間無顯著差異, 但在沙丘頂部, 表現為麥草方格顯著高于尼龍網方格。前者原因可能是沙丘底部土壤相對濕潤[24], 蠼螋偏喜于在土壤水分條件較好的沙地環境中生存, 故蠼螋科個體數分布較多而成為優勢類群, 且在2種沙障材料林地間無顯著差異。后者原因可能是沙丘頂部麥草沙障可以作為枯落物, 為植食性動物提供食物來源的同時, 亦間接為捕食性蠼螋科個體提供了潛在的食物來源[5], 從而導致麥草方格林地蠼螋科個體數顯著高于尼龍網方格。
本研究還發現, 無論是在沙丘底部還是頂部, 麥草方格內地面節肢動物個體數、類群數均顯著高于尼龍網方格。分析原因主要是麥草沙障作為枯落物可以為地面節肢動物提供食物來源, 同時又具有保水性能, 微生境的改善可以為地面節肢動物活動提供生存條件[5], 故麥草沙障較尼龍網沙障更有利于地面節肢動物的生存。但地面節肢動物群落多樣性表現為無論是在沙丘底部還是頂部, 地面節肢動物Shannon指數、優勢度指數均表現為麥草方格和尼龍網方格間無顯著差異。分析原因可能是經過5年的草方格固沙, 某一部位麥草被風蝕裸露后可能會加速鄰近區域麥草的裸露, 麥草所起的保持水分和提高養分的作用隨之減弱[19,27], 從而導致不同沙障林地地面節肢動物多樣性分布無顯著差異性, 同時也說明不同材料沙障對地面節肢動物多樣性分布的影響有限。
3.3 地面節肢動物群落與環境因子的關系
本研究RDA和偏RDA分析結果表明, 花棒冠幅、花棒高度和檸條高度均對地面節肢動物個體數分布產生顯著影響, 這與董煒華等[29]的研究結果相似。但是地面節肢動物個體數分布受到的土壤環境因子較小, 這與已有的研究結果不一致[30,32]。許多研究結果已經證實, 荒漠水分和全氮含量是影響荒漠節肢動物生長和分布的重要環境因子[33–34]。在景觀及更大尺度上, 氣候條件如溫度和降水是影響地面節肢動物分布的環境因子[35–36]。劉任濤等[37]在干旱區不同林齡灌叢下研究表明, 土壤容重、pH和電導率對地面節肢動物群落結構影響顯著。綜合分析表明, 本研究所調查的土壤因子并不能反映環境條件對地面節肢動物群落分布的影響, 并且相關環境因子(包括植被和土壤因子)對地面節肢動物個體數分布的總貢獻率僅為29.5%, 說明仍有其他重要因素如土壤全P、全K、有機質等因子對地面節肢動物個體數分布產生影響[37–38], 今后可能需要對土壤全K、全P和有機質含量等其他因素進行調查。
4 結論
(1)地面節肢動物群落優勢類群包括步甲科、擬步甲科、蠼螋科和葉甲科, 反應了地面節肢動物對沙漠特殊生境的選擇性和適應性。
(2)沙障材料對地面節肢動物群落組成、個體數和類群數分布均產生顯著影響, 而且地面節肢動物群落組成分布亦受到沙丘微地形的影響。
(3)麥草方格較尼龍網方格可以更有效豐富地面節肢動物個體數, 但沙障材料對地面節肢動物多樣性分布的影響較小。
(4)灌叢營造+草方格措施可以對固沙林地地面節肢動物個體數分布產生直接影響, 特別是花棒冠幅、花棒高度和檸條高度成為主要限制因素。
[1] 常海濤, 劉任濤, 劉佳楠, 等. 草方格造林固沙過程中土壤性質變化及分形特征—以騰格里沙漠東南緣為例[J]. 水土保持學報, 2018, 32(3): 58–65.
[2] LI Jinchang, LIU Haixia, SU Zhizhu, et al. Changes in wind activity from 1957 to 2011 and their possible influence on aeolian desertification in northern China[J]. Journal of Arid Land, 2015, 7(6): 755–764.
[3] 屈建軍, 劉賢萬, 雷加強, 等. 尼龍網柵欄防沙效應的風洞模擬實驗[J]. 中國沙漠, 2001(3): 62–66.
[4] LI Xinrong, CHENG Yingwu, SU Yangui, et al. Effects of biological soil crust on desert insect diversity: evidence from the Tengger Desert of northern China[J]. Arid Land Research and Management, 2006, 20: 1–18.
[5] 胡英娣, 周潔. 用于草方格沙障的麥草方格保存研究[R]. 沙坡頭沙漠研究試驗站編, 1991.
[6] 馬學喜, 王海峰, 李生宇, 等. 兩種固沙方格沙障的防護效益及地形適應性對比[J]. 水土保持通報. 2015, 35(3): 344–349.
[7] 張立欣, 段玉璽, 王博, 等. 庫布齊沙漠不同人工固沙灌木林土壤微生物量與土壤養分特征[J]. 應用生態學報, 2017, 28(12): 3871–3880.
[8] 屈建軍, 凌裕泉, 俎瑞平, 等. 半隱蔽格狀沙障的綜合防護效益觀測研究[J]. 中國沙漠, 2005, 25(3): 329–335.
[9] 董治寶, FRYREAR DONALD W, 高尚玉. 直立植物防沙措施粗糙特征的模擬實驗[J]. 中國沙漠, 2000, 20(3): 260–263.
[10] 李瑞軍. 棉稈沙障防風固沙效益比較[D]. 蘭州: 甘肅農業大學, 2009: 47.
[11] 孫濤, 劉虎俊, 朱國慶, 等. 3種機械沙障防風固沙功能的時效性[J]. 水土保持學報, 2012, 26(4): 12–16.
[12] 王雨浩. 麥草、PLA及其混合沙障防風固沙效益研究[D]. 呼和浩特: 內蒙古師范大學, 2019: 78.
[13] 劉繼亮, 趙文智, 李鋒瑞, 等. 微地形變化對沙質荒漠擬步甲科昆蟲分布的影響[J]. 干旱區研究, 2017, 34(6): 1388–1394.
[14] 劉任濤, 趙哈林, 趙學勇. 科爾沁沙地不同造林類型對土壤動物多樣性的影響[J]. 應用生態學報, 2012, 23(4): 1104-1110.
[15] 王新譜, 楊貴軍. 寧夏賀蘭山昆蟲[M]. 北京: 科學出版社, 2010.
[16] 尹文英. 中國土壤動物檢索圖鑒[M]. 北京: 科學出版社, 1998.
[17] 鄭樂怡, 歸鴻. 昆蟲分類[M]. 南京: 南京師范大學出版社, 1999.
[18] 劉任濤, 郗偉華, 劉佳楠, 等. 沙地檸條(Caragana)灌叢微生境節肢動物群落特征[J]. 中國沙漠, 2018, 38(1): 117–125.
[19] 羅雅曦. 不同材料沙障固沙林地土壤—植被系統變化及其生態效應評價[D]. 銀川: 寧夏大學, 2019: 40.
[20] 鮑士旦. 土壤農化分析3版[M]. 北京: 中國農業出版社, 2000.
[21] 趙娟,劉任濤,劉佳楠, 等.北方農牧交錯帶退耕還林與還草對地面節肢動物群落結構的影響[J].生態學報, 2019, 39(5): 1653–1663.
[22] 蘇永中, 趙哈林, 張銅會. 幾種灌木、半灌木對沙地土壤肥力影響機制的研究[J]. 應用生態學報, 2002, 13(7): 802–806.
[23] 鮑乾, 楊瑞, 李萬紅, 等. 喀斯特高原峽谷區不同恢復模式的土壤生態效應[J]. 水土保持學報, 2017, 31(3): 154–161+168.
[24] 高國雄. 毛烏素沙地東南緣人工植被結構與生態功能研究[D]. 北京: 北京林業大學, 2007: 181.
[25] 尹輝. 我國西南典型喀斯特峰叢洼地土壤理化特征研究[D]. 北京: 中國地質科學院, 2012: 143.
[26] 莫保儒, 蔡國軍, 楊磊, 等. 半干旱黃土區成熟檸條林地土壤水分利用及平衡特征[J]. 生態學報, 2013, 33(13): 4011–4020.
[27] 王逸敏. 固沙沙障對植被、土壤和生物土壤結皮的影響[D]. 西安: 西北大學, 2018: 68.
[28] 張樂, 劉志民. 丘間低地生態過程研究進展[J]. 生態學雜志, 2007(7): 1101–1106.
[29] 董煒華, 李金霞, 劉繼亮. 沙丘和丘間低地地面節肢動物群落比較研究[J]. 干旱區資源與環境, 2016, 30(1): 157–161.
[30] 婁巧哲, 徐養誠, 馬吉宏, 等. 古爾班通古特沙漠南緣地表甲蟲物種多樣性及其與環境的關系[J]. 生物多樣性.2011, 19(4): 441–452.
[31] 劉淑蓉. 天津八仙山鱗翅目幼蟲多樣性初步研究[D]. 天津: 南開大學, 2011: 90.
[32] 常海濤,劉佳楠,趙娟,等. 烏拉特荒漠草原紅砂灌叢林地地面節肢動物群落季節分布特征[J]. 生態學報, 2020 (9): 1–10.
[33] 趙文智, 劉鵠. 干旱、半干旱環境降水脈動對生態系統的影響[J]. 應用生態學報, 2011, 22(1): 243–249.
[34] 趙哈林, 劉任濤, 趙學勇, 等. 旱作農田改為水澆地對沙質土壤節肢動物群落的影響[J]. 干旱區資源與環境, 2014, 28(1): 9–14.
[35] LI Fengrui, LIU Jiliang, LIU Changan, et al. Shrubs and species identity effects on the distribution and diversity of ground - dwelling arthropods in a Gobidesert[J]. Journal of Insect Conservation, 2013: 1–13.
[36] ZHAO Halin, LIU Rentao. The " bug island" effect of shrubs and its formation mechanism in Horqin Sand Land, Inner Mongolia[J]. Catena, 2013, 105: 69–74.
[37] 劉任濤, 朱凡, 柴永青. 干旱區不同年齡灌叢斑塊地面節肢動物的聚集效應[J]. 應用生態學報, 2014, 25(1): 228–236.
[38] 劉繼亮,李鋒瑞,牛瑞雪,等.黑河中游干旱綠洲土壤鹽漬化對大型土壤動物群落的影響[J]. 應用生態學報. 2012, 23(6): 1551–1561.
Effects of shrub revegetation within different sand barrier materials on the community structure of ground-active arthropods in desert ecosystems
FENG Yonghong1, GOU Qi2,3, LIU Rentao2,3,*, LUO Yaxi2, CHANG Haitao2, ZHANG Jing1, JIANG Jiayu2,3
1. College of Agriculture, Ningxia University, Yinchuan 750021, China 2. Key Laboratory for Restoration and Reconstruction of Degraded Ecosystems in Northwestern China of Ministry of Education, Ningxia University, Yinchuan 750021, China 3. College of Ecology and Environment, Ningxia University, Yinchuan 750021, China
The aim of this study was to probe into the effects of different sand barrier materials on arthropod community structure in afforested shrub plantations in desert ecosystems. The composition and diversity distribution of ground-active arthropod communities were investigated at the bottom and top of sand dunes within checkerboards of wheat straw and nylonin spring, summer and autumn of 2018 in the southeast edge of Tengger Desert. The results showed that: (1) At dune bottom, there were three dominant groups including Labiduridae, Carabidae and Chrysomelidae observed in the microhabitats within checkboards of two materials. In addition, there were two groups including Tenebrionidae and Coleoptera larvae in the microhabitats within wheat-straw checkerboard, and one group including Lepidoptera larvae in the microhabitats within nylon checkerboard. However, at dune top, there were similar dominant groups in the microhabitats between the two materials including Labiduridae, Carabidae, Chrysomelidae, and Tenebrionidae. (2) There was no significant difference in the abundance of Carabidae and Chrysomelidae between the microhabitats at dune bottom and top. At dune bottom, the abundance of Tenebrionidae was significantly higher in the microhabitats within wheat-straw checkerboard than within nylon checkerboard, and the abundance of Labiduridae was found to indicate no significant difference between the dune microhabitats within two materials; at dune top, there was no significant difference in the abundance of Tenebrionidae between the microhabitats within two materials, and the abundance of Labiduridae was significantly higher in the microhabitats within wheat-straw checkerboard than within nylon checkerboard. (3) At dune bottom and top, the total abundance and group richness of ground-active arthropods was significantly higher in the microhabitats within wheat-straw checkerboard than within nylon checkerboard, but there was no significant difference in Shannon index and dominance index in the microhabitats between the two materials. (4) The results of RDA and partial RDA analysis showed that the height and crown width of, and crown width ofwere the main factors driving the differences of abundance distribution of ground-active arthropods between different microhabitats. It was concluded that sand barrier materials indicated significant effects on the composition, abundance and group richness distribution of ground-active arthropod community, and their composition and abundance distribution were also affected by the dune microhabitats, but there was little effect of sand barrier materials on the ground-active arthropod diversity distribution in Tengger Desert ecosystem.
Tengger Desert; sand barrier material; dune microhabitats; ground-active arthropod; community structure
馮永宏, 茍琪, 劉任濤, 等. 不同沙障材料對固沙林地面節肢動物群落結構分布的影響[J]. 生態科學, 2022, 41(5): 35–45.
FENG Yonghong, GOU Qi, LIU Rentao, et al. Effects of shrub revegetation within different sand barrier materials on the community structure of ground-active arthropods in desert ecosystems[J]. Ecological Science, 2022, 41(5): 35–45.
10.14108/j.cnki.1008-8873.2022.05.005
958.15+5
A
1008-8873(2022)05-035-11
2020-08-19;
2020-09-27
寧夏自然科學基金(2020AAC0214, 2021AAC03047); 寧夏重點研發計劃項目(2021BEG03007); 國家自然科學基金(41867005)
馮永宏(1996—), 男, 陜西榆林人, 碩士研究生; 主要從事水土保持與荒漠化防治研究, E-mail: 13772928515@163.com
劉任濤, 男, 博士, 研究員, 主要從事防沙治沙與生態恢復相關研究, E-mail: nxuliu2012@126.com
