珍稀瀕危植物獨花蘭研究進展
2022-08-10 02:00:32劉旭陽王仕寶朱志凱
陜西農業科學 2022年6期
關鍵詞:植物
張 慧,劉旭陽,王仕寶,朱志凱,吳 鵬
( 1.漢中職業技術學院 藥學院;陜西 漢中 723000;2.漢中職業技術學院 秦巴山區藥(食)用植物研究所,陜西 漢中 723000;3.漢中市南鄭區食品藥品檢驗檢測中心,陜西 漢中 723000 )
蘭科獨花蘭(ChangnieniaamoenaS. S. Chien)是我國特有的單種屬植物,其藥用價值較高,在《全國中草藥匯編》中記載:其藥材名為長年蘭,以假鱗莖或全草入藥,其功效為清熱、涼血、解毒,主治咳嗽,痰中帶血,熱癤疔瘡[1]。獨花蘭的外部形態特殊、色彩美麗,全株只有一葉一花,葉片寬卵狀橢圓形至寬橢圓形,一般長6.5~11.5 cm,寬5~8.2 cm,葉片上面綠色,背面紫紅色。花葶頂生,長10~17 cm,紫色,花白色、帶淡紫色或肉紅色暈,唇瓣上有紫紅色斑點[2]。近年來,由于環境的變化、過度的采挖及自身繁育困難等因素,導致獨花蘭資源急劇減少,1992年被《中國植物紅皮書—稀有瀕危植物( 第一冊) 》列為珍稀瀕危種,成為國家二級保護植物[3]。文章主要針對獨花蘭的分布區域、群落特征、繁育特征、組織培養育苗技術和遺傳與變異五個方面方面進行綜述,以期為獨花蘭的深入研究提供參考依據。
1 分布區域
查閱中國植物志[2]、FRPS圖庫照片分布圖(圖1)及相關文獻[4~8],得出獨花蘭分布在13個省(市)區,主要以湖北、湖南等華中地區為中心,北至河南、陜西南部,南至浙江、江西及廣西等地,西至四川,甘肅南部,東至安徽、河南一帶。此外,還包括重慶、貴州等地。另有報道[9]:獨花蘭于1970年在陜西省平利縣首次發現,之后四十多年再未發現,直到2011年和2012年分別在陜西化龍山國家級自然保護區內被重新發現。2018-2020年,進行中藥資源普查期間,在陜西寧強縣域發現多個獨花蘭野生分布群落[10]。
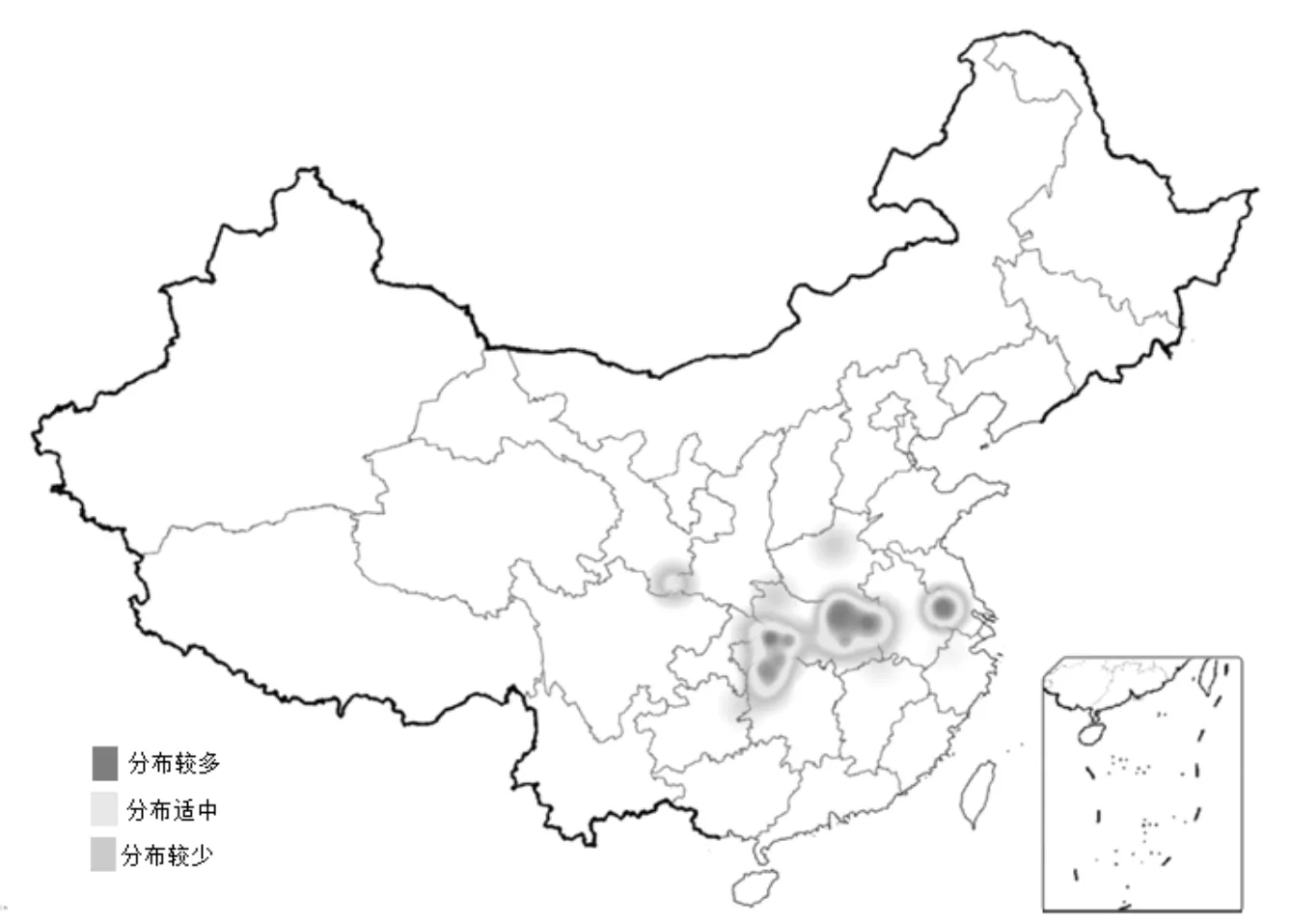
圖1 FRPS圖庫照片分布
2 群落特征
生物群落特征主要包括群落中物種的多樣性、群落的生長形式、結構、優勢種等多個方面。獨花蘭群落一般分布于坡度15~45°的陰坡,海拔范圍常為400~1 700 m,山地黃棕壤,郁閉度40%~75%,林下腐殖質豐富、水分充足的土壤,同時排水性良好。鄔剛等基于鄰接格子法和直接計數法,研究了英山縣現有的168株獨花蘭的種群數量及分布格局,利用方差/均值比率法,得出草盤鎮桃花沖村獨花蘭小種群為集群分布[11]。神農架南坡獨花蘭主要分布的伴生植物,包括以化香樹、四照花、華千金榆等為主的落葉闊葉林及箬竹組成的灌叢下,北溫帶物種為群落中的優勢物種[12]。雞公山保護區獨花蘭主要分布在落葉闊葉林和杉木林中,主要植被有水杉、樸樹、元寶槭、常春藤、絡石、山胡椒、絞股藍等[13]。貴州臺江縣獨花蘭的生境調查發現其生于針闊混交疏林下,主要喬木樹種有杉木、金葉子、鵝掌楸、楠木等,主要灌木有櫟屬,主要草本有苔草屬、百合屬、蝦脊蘭、金蘭等植物[14]。王仕寶等在陜西寧強縣區域內落葉闊葉林或針闊混交林中發現了獨花蘭野生種群,其植被喬木層有殼斗科、松科及樺木科;灌木層主要有小檗科、山茱萸科、忍冬科、漆樹科、冬青科、禾本科(竹亞科)等;草本層主要包括百合科、唇形科、蘭科、鹿蹄草科、薯蕷科、禾本科(禾亞科)、龍膽科等植物[10]。
3 繁育特征
3.1 繁育與傳粉
獨花蘭是自交親和的蟲媒花,正常情況下需借助前來吮吸花粘液分泌物的昆蟲觸及隱藏在藥帽下的花粉或粘盤,將花粉塊帶到柱頭凹陷處的腔室內完成授粉。雖然獨花蘭具有花距,但不分泌花蜜,不能為傳粉者提供回報,是欺騙性傳粉模式[15],并且獨花蘭花期在4月份,海拔較高區域溫度較低,受訪昆蟲偏少,傳粉者較少。研究表明在獨花蘭群落內出現的昆蟲有15種,為其傳粉的只有熊蜂。三條熊蜂是神農架龍門河群體傳粉者,仿熊蜂是神農架關門山群體的唯一傳粉者。可見,傳粉者限制是導致自然結實率低的主要原因[16]。此外,結合文獻[17],對獨花蘭的生理構造研究表明,花藥帽較大,并且包蓋緊密,不利于花粉塊的外露和傳播;花粉塊柄不發達,不能將花粉塊托起將花藥帽頂開;粘盤不發達,無法將花粉塊帶出藥帽。因此,整個花期內,如果沒有昆蟲的幫助,花粉塊將一直包藏在花藥帽下,無法正常完成授粉過程。獨花蘭的這些生物學缺陷導致了其只開花不結果,繁殖率低下。琚煜熙[12]跟蹤調查雞公山自然保護區內14個分布群的340株獨花蘭,發現其開花株數為215株,開花率為 63.2%,結果株數僅有12株,結實率僅為5.6%。桂先群[18]分別于2008年4月24日和2014年4月25日對天堂寨自然保護區內的獨花蘭進行人工授粉。結果顯示,自花受粉后花被枯萎凋落,子房會伸長,果實成熟期結果,未受粉的花被鮮艷,子房不會伸長,果實成熟期不結果,表明獨花蘭是自交親和的異交種。同時,該試驗除進一步證實了獨花蘭的生物學特性是其致危的重要因素外,還對人工授粉技術提高其種群密度的應用提供了依據。
3.2 繁育與菌根
蘭科植物作為最進化復雜的類群之一。在自然環境下,蘭科植物的大多數種子極為細小,缺乏胚乳組織,需要和適宜的真菌共生才能萌發。獨花蘭作為蘭科植物也不例外,種子的胚發育不完全,沒有貯藏種子萌發所需的營養組織,在自然條件下,獨花蘭的種子不能正常萌發;當獨花蘭的種子與特定的真菌形成穩定的共生關系后,才使種子利用真菌提供的營養物質, 繼而萌發并生長成植株。武靜宇根據獨花蘭菌根真菌培養的特點及其形態學特征,初步將其菌根真菌鑒定為蘭科共生絲核菌,后采用分子生物學方法,將其菌根真菌的rDNA ITS序列進行Blast同源比對,進一步得出該真菌屬于膠膜菌屬[19]。有文獻報道獨花蘭具有典型的蘭科植物菌根構造;分離得到的獨花蘭的菌根真菌主要屬于擔子菌和有絲孢菌的菌株,鐮刀菌屬為優勢種屬;利用回接試驗法來判斷從獨花蘭根中分離到的真菌是否為菌根真菌時發現大部分菌株對華石斛組培幼苗生長有不同程度的促進作用[20~21]。基于此,通過篩選適合的真菌進行獨花蘭種子的共生萌發試驗,為有效實現獨花蘭植物的種質保育,進行瀕危野生居群的生態恢復顯得尤為重要。
3.3 繁育與營養體
花的發育和開花對物種繁殖具有重要意義。對于多年生植物, 其生殖生長需要建立在若干年的營養生長的基礎之上。因此,植株營養生長狀況影響著花的發育和開花。獨花蘭的地下部生長有假鱗莖,假鱗莖作為一種變態莖,是獨花蘭植物體的重要營養器官,每個假鱗莖的頂端生有1個葉片,并且是花芽著生的地方。獨花蘭葉片大小能夠體現植株個體大小,反映植株所處的發育階段。葉片大小與地下莖體積和假鱗莖數目呈高度正相關,同時,依據葉片的大小、假鱗莖的數量及地下莖的體積大小可以判斷獨花蘭是否達到有性生殖的狀態;其葉片越大,光合作用產物就會越多,可以滿足有性生殖的需要。熊治廷對安徽天堂寨自然保護區內野生獨花蘭進行了兩月有余的連續定株觀測和群體統計,發現獨花蘭植株開花與否和地下假鱗莖的數目、地下莖的總體積和葉片的面積具有極顯著相關性,絕大多數開花的獨花蘭地下假鱗莖數為3,地下莖總體積通常達8 cm3,葉片的面積達 33 cm3[22]。個體較大的植株,葉片大,光合產物多,開花持續期較長。此外,獨花蘭可依靠自身的假鱗莖進行緩慢的無性生殖[23],其假鱗莖具新老更替特點,經過1個生長季后即不再生葉開花,以后由新的假鱗莖繼續展葉開花,因老的假鱗莖所儲存的營養物供應了抽葶開花的需要,在花謝后老的假鱗莖呈現出萎縮壞死狀[22]。陳戈[24]等在春秋兩季利用假鱗莖,采取組織個體分株法進行獨花蘭繁育,獨花蘭生長2~3年可進行1次分株,分株后至少需要有5個連接的假球莖。
4 組織培養育苗
為解決獨花蘭自然情況下的繁殖困難,傳粉受精與結實率較低的問題,相關學者通過研究,成功提出了獨花蘭的快速繁育方案。Jiang[25]等分別以4月和6月份采集的假鱗莖和節段化假鱗莖為外植體,1/2MS+1.0 mg/L6-BA+0.05 mg/LNAA+10%椰子汁培養基為芽誘導培養基,結果顯示,節段化假鱗莖上沒有芽產生,認為這與假鱗莖切面上有大量酚類物質的滲出,導致節段化的假鱗莖壞死有關;4月份采集的假鱗莖為外植體培養12周可產生腋芽,6月份所采的假球莖為外植體,培養7周后,每個外植體產生1.4個芽,這一現象與獨花蘭4月花期營養物質的消耗有關,同時認為以獨花蘭假鱗莖為外植體時,適宜采收期為6月上旬。最佳生根誘導培養基為1/2MS+1.0mg/LNAA+0.1 mg/L6-BA,平均每芽生根4.5條,最高生根率為88.9%。將4~5 cm高的試管苗移栽到腐殖質:沙土(1:1)混合物的盆栽中,在溫室中生長7周后移栽到大田,之后在野外生長兩個月后成活率約為75%。此外,劉國順[3]、高麗[26]、劉賽思[27]等也研究了獨花蘭的組織培養技術。劉國順發現75%酒精消毒30 s,然后0.2%HgCl消毒6~7 min為外植體消毒的最佳方式,試驗外植體的污染率為14.3%[3]。結合相關文獻,對獨花蘭不同外植體、培養基、誘導率和生根率進行了匯總(見表1)。目前,組織培養育苗技術是實現獨花蘭快速規模化繁殖的一種有效手段。

表1 文獻中記載的獨花蘭組織培養情況
5 遺傳變異特征
基于BLAST1鑒定、遺傳變異、建樹分析,巫偉峰[28]等對12個蘭屬本地樣品和250條GenBank下載的蘭科ITS2序列(其中81條屬于蘭屬)進行了評估。結果表明,ITS2序列在蘭屬植物的分子鑒定、親緣關系、系統發育關系等方面具有一定的應用價值,可作為蘭屬植物DNA條形碼鑒定的輔助條形碼。獨花蘭作為蘭科的單種屬植物,可以結合ITS2序列對同族植物進行研究DNA條形碼和系統發育的關系。為了研究獨花蘭的分類地位,Yi[29]利用下一代測序技術測定了獨花蘭的葉綠體全基因組序列,結果表明獨花蘭與布袋蘭親緣關系較近。Li[30~31]結合傳粉和生態學的研究結果,利用用隨機擴增多態DNA(RAPD)技術對獨花蘭的遺傳多樣性水平和分配進行了研究,結果表明,可能是由于生境破壞致使獨花蘭數量減少甚至局部滅絕,基因流動受限導致獨花蘭群體內變異低而群體間變異大;并提出了獨花蘭棲息地保護和傳粉者保護、人工授粉以及遷地保護的保護管理措施。孫海芹[32]等采用(單因素方差分析法)和UPGMA聚類分析等方法,研究了分布于神農架、廬山和新寧的12個自然居群獨花蘭的形態變異,發現廬山居群13個性狀的平均觀測值均高于新寧和神農架居群,在物種水平上各性狀存在較多的變異,并且營養器官變異性較高。單因素方差分析結果顯示3個地區間多個形態性狀存在極顯著差異;UPGMA聚類分析表明這3個地區分別形成明顯不同的分支。在以傳粉者為媒介自然選擇的影響下,神農架龍門河和關門山兩個地點間存在明顯的形態分化。
6 展望
目前,關于獨花蘭的研究報道相對較少,主要集中在分布區域、群落特征、繁育特征、組織培養育苗技術及遺傳與變異等方面,尚未見關于獨花蘭的化學成分、藥理作用等方面的報道,這可能與獨花蘭為瀕危植物,資源稀缺有較大關系。獨花蘭是一種珍貴的瀕危藥用植物,具有較高的藥用價值;且獨花蘭耐陰又耐寒,植株矮小,花色艷麗,具有較高的觀賞價值,是室內培養的良好的花卉品種,這便導致了獨花蘭采挖過度,資源減少。同時,獨花蘭對傳粉者的高度依賴和專一性,致使傳粉困難,結實率低,還有種子的萌發也需要與菌根真菌形成共生關系,在適宜的溫度和濕度下才能萌發,生態環境的破壞可能影響其傳粉者和菌根真菌,從而影響獨花蘭的自然生長與繁育,這又進一步導致了獨花蘭資源的稀缺。針對現狀,應該積極采取應對措施保護獨花蘭生境,利用人工授粉和組織培養育苗技術提高獨花蘭的繁育率,豐富其資源。在此基礎上,對獨花蘭進行更深入的研究,以期在藥用、生態、遺傳及觀賞等方面體現更高的綜合價值。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
