蕾期低溫脅迫對棉花光合特性的影響
2022-08-10 02:10:28鐘信念梁其干李有忠趙建華馬海霞李志博
新疆農業科學 2022年7期
鐘信念,梁其干,李有忠,3,趙建華,馬海霞,李志博
(1.石河子大學農學院/新疆兵團綠洲生態農業重點實驗室;2.新疆農業科學院/海南三亞農作物育種試驗中心,海南三亞 572014;3.新疆農墾科學院生物技術研究所/作物種質創新與基因資源利用兵團重點實驗室,新疆石河子 8320032)
0 引 言
1 材料與方法
1.1 材 料
供試材料為3個新疆北疆不同年代北疆主栽陸地棉棉花品種新陸早10號(1999年)、新陸早36號(2007年)、新陸早61號(2013年),棉花種子均由新疆兵團綠洲生態農業重點實驗室棉花育種室提供。
1.2 方 法
1.2.1 棉苗培養及處理
試驗于2019年11月至2021年1月于新疆石河子市石河子大學綠洲生態實驗室內購置的浙江寧波儀器廠人工智能氣候箱中進行。選用大小相同的新發芽盒,3個品種各挑選300粒飽滿健康且已經經過硫酸脫絨洗凈的新陸早10號、新陸早36號、新陸早61號棉籽作為材料,用0.4%的高錳酸鉀溶液對種子進行消毒,消毒時間為10~15 min,然后用蒸餾水560將表面殘留的高錳酸鉀溶液沖洗3~4次,沖洗后放置于經75%酒精消毒的燒杯中備用。放置于墊有干凈濾紙(防止種子根部穿過紗布網眼)及紗布的發芽盒中,用56℃溫水泡種20~25 min(加快萌發),每個發芽盒放40粒種子置于加有足以潤濕發芽床的少量水,放入28℃培養箱培養,觀察種子發芽長勢,當芽長1 cm時取出種入花盆。
采用室內人工氣候箱盆栽法培養棉花幼苗,選取320x250 mm規格的花盆進行播種育苗。培養土采用花卉市場采購的花土與蛭石以4∶1的比例進行混配后作為基土栽培種植。每盆保苗4株,株距10 cm,各溫度每個品種設置4盆作為重復(3盆處理、1盆對照)。出苗后每盆保留4棵長勢一致的棉苗繼續生長,每3 d以適宜的水肥澆灌于托盤從底部滲透進基土以防止出現土壤板結等問題及便于植株吸收,同時適宜的蹲苗防止出現弱苗、高腳苗、根扎不深等栽培問題。實時觀察及時進行蟲害管理。圖1

圖1 實驗培養箱
前期培養光照強度設置為50 000xl、晝夜溫度為28/20℃、光周期16 h、濕度為60%;待實驗材料長至初始現蕾期時,將長勢一致的實驗材料放入人工智能氣候箱內低溫處理培養。 試驗條件濕度均設置為60%,低溫條件分別設置為①光照強度20 000 lx(光照強度根據低溫強度設置設置相應的弱光進行調節)晝夜溫度20℃/15℃(白天20℃晚上15℃)光周期16 h、②光照強度15 000 lx晝夜溫度15℃/10℃光周期16 h、③光照強度10 000 lx晝夜溫度10℃/5℃光周期16 h三個低溫梯度處理。
對照組設置光照強度為50 000 lx、晝夜溫度28℃/20℃光周期16 h進行處理,于實驗材料現蕾期開始給予低溫脅迫至初花期,期間每處理5d后選取棉花倒4功能葉進行光合及葉綠素熒光參數的測定。去掉一個最高和最低測定值,取其余的平均值作為相應的測定值。
1.2.2 指標測量
使用美國LI-COR 公司便攜式光合儀 LI-6400XT測定植物的凈光合速率(Pn)、氣孔導度(Gs)、胞間 CO2濃度(Ci)和蒸騰速率(Tr),測定光強為 1 000 μmol/(m2·s),CO2濃度為400 μmol/mol,葉室溫度為 28℃,流速為 500 μmol/s。選取蕾期棉花倒四片功能葉,并做標記,測定時避開葉脈。之后每次測定使用相同葉片,每枚葉片測定2次,每個處理6 株重復。
Nguyen(2012)發現當首席執行官和董事屬于同一社會網絡時,CEO不太可能因為業績不佳而被解雇,董事會成員和CEO之間密切的社會關系會影響董事會職能的發揮;Chahine et al.(2014)發現只有在控制董事會成員與高管的社交聯系后,董事會獨立性才能發揮效應。那么關于社會關系的度量又該如何展開呢?國外大部分學者的研究集中在工作經歷、畢業院校、老鄉、專業方面 (Kramarzy et al.2007、 Hwang et al.2009、Liu 2010、Nguyen 2011、Hoitash 2011、Fracassi et al.2012等)。
1.3 數據處理
使用SPSS 26.0軟件處理本試驗數據統計分析,P<0.05表示差異顯著,應用Excel 2019、origin2021等畫圖軟件綜合分析作圖。
2 結果與分析
2.1 蕾期低溫脅迫對凈光合速率(Pn)的影響
研究表明,隨著處理溫度的降低,不同品種的凈光合速率均呈顯著下降趨勢,且溫度越低則降幅越大;隨低溫脅迫天數的增加,不同品種的凈光合速率顯著下降,脅迫時間越長Pn值越小。T1(20℃/15℃)處理下,各品種相對于對照,Pn緩慢下降。T2(15℃/10℃)低溫下Pn顯著低于對照和T1(20℃/15℃),T2(15℃/10℃)處理 10 d的Pn大幅度下降,極顯著低于處理0 d、5 d;T3(10℃/5℃)的Pn顯著低于T1(20℃/15℃)、T2(15℃/10℃)且隨脅迫時間的延長Pn值越來越小。
當低溫脅迫溫度高于20℃/15℃時新陸早61號的凈光合速率(Pn)下降的幅度較新陸早36號、新陸早10號趨勢小,T1(20℃/15℃)處理下,同一時間3個蕾期棉花品種葉片的Pn值由大到小依次為新陸早61號>新陸早36號>新陸早10號>。當低溫脅迫溫度低于15℃/10℃時新陸早36號相對于新陸早10號、新陸早61號下降的幅度較小。T2(15℃/10℃)、T3(10℃/5℃)處理下,同一時間3個蕾期棉花品種葉片的Pn值由大到小依次為新陸早36號>新陸早61號>新陸早10號>,中度、重度低溫脅迫下新陸早36號相對其它2個品種更耐低溫。 同一個低溫下,隨著低溫脅迫時間的延長,凈光合速率(Pn)下降加劇;且同一個處理時間段內(15 d),低溫脅迫強度越大,凈光合速率(Pn)受到的影響越大。圖2
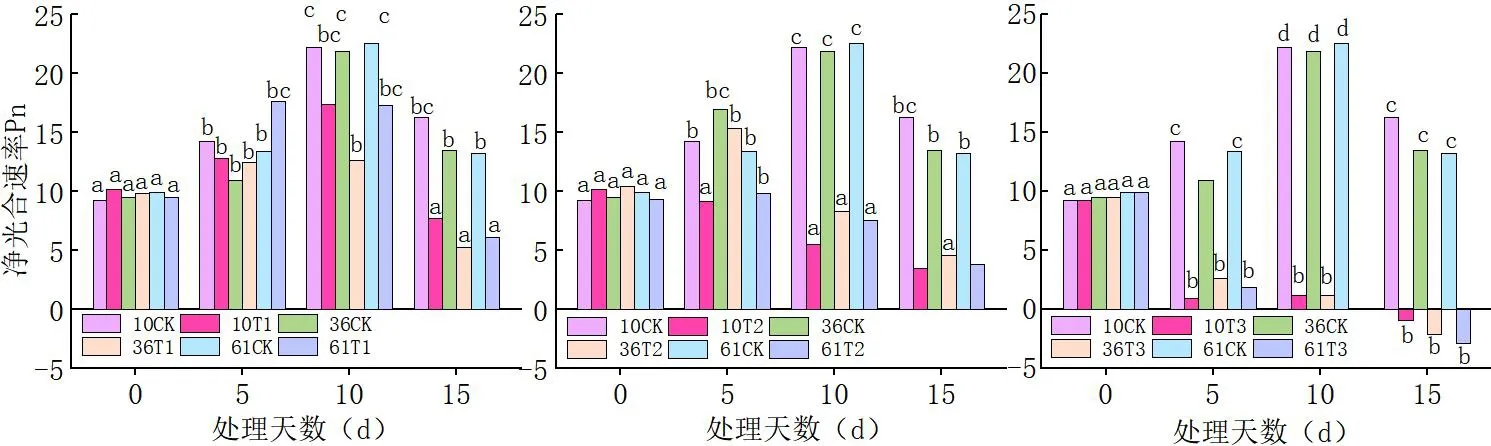
圖2 低溫脅迫下棉花凈光合速率(Pn)變化
2.2 蕾期低溫脅迫對氣孔導度(Gs)的影響
研究表明,T1(20℃/15℃)與T2(15℃/10℃)、T3(10℃/5℃)存在顯著差異,總體上溫度下降氣孔導度值變小;同一溫度下不同時間各處理也差異明顯,T1、T2處理下Gs值先升后降。T3處理下Gs值直線下降。脅迫溫度從 20℃/15℃降至10℃/5℃時,蕾期棉花參試品種的Gs含量均大幅度下降,低于 15℃/10℃的低溫處理葉片的Gs含量雖呈下降趨勢,但降幅較小,10℃/5℃處理10 d 植株死亡。脅迫 15d 各處理的Gs含量均下降到最低水平,不同天數間差異顯著。從單變量全因子分析的結果來看,溫度、時間對蕾期棉花葉片氣孔導度的變化均有影響,而且溫度與處理天數有交互作用,P<0.001。溫度和時間產生的低溫危害效應與Pn相似。圖3
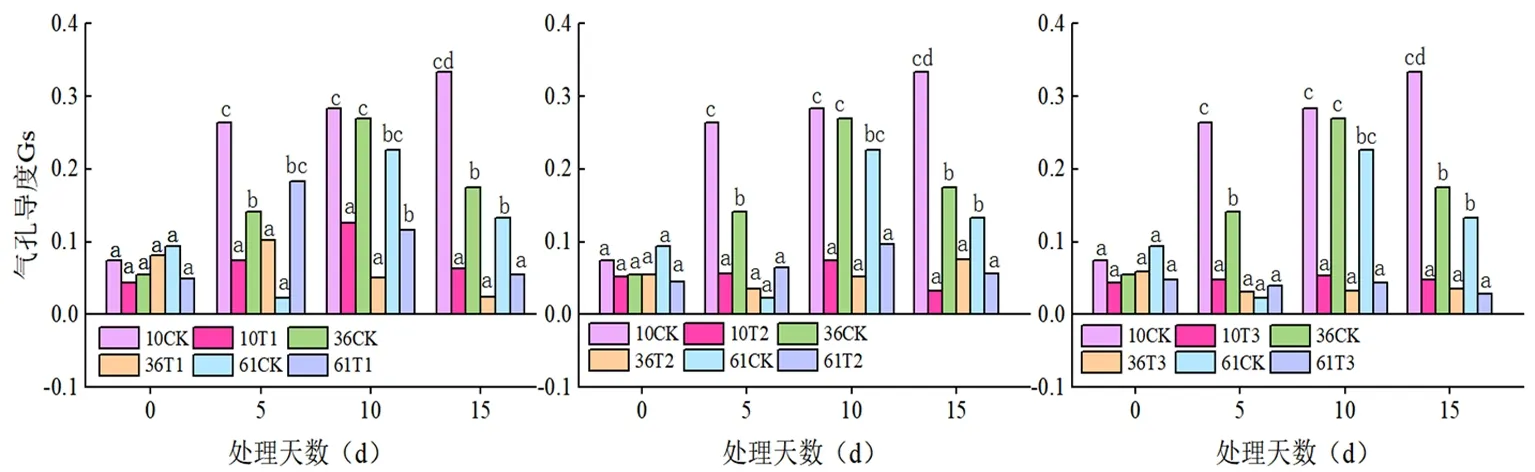
圖3 低溫脅迫下氣孔導度(Gs)變化
2.3 蕾期低溫脅迫對細胞間CO2濃度(Ci)影響
研究表明,不同低溫脅迫初期(5 d)各品種的Ci值差異不顯著,脅迫后期(15 d)各品種Ci值差異較為顯著。T1脅迫10 d內各品種蕾期棉花的Ci差異不明顯,T3脅迫下同一處理溫度不同脅迫天數之間差異較明顯。T2、T3脅迫下各品種的Ci與CK組的差異均隨處理持續時間的延長而加大,T1(20℃/15℃)處理下Ci值呈升-降-升的變化趨勢,T2(15℃/10℃)處理下Ci呈先升后降的變化趨勢,T3(10℃/5℃)處理下Ci的變化趨勢呈急劇上升,總體上各低溫脅迫組的Ci值均大于對照。T3(10℃/5℃)脅迫 15 d 時達最大值。新陸早61號的Ci值上升幅度最小,新陸早61號蕾期的抗冷性較好。圖4
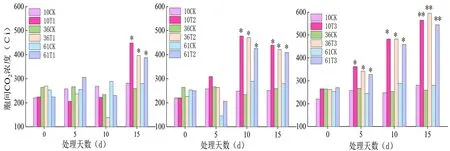
圖4 低溫脅迫下細胞間CO2濃度(Ci)變化
2.4 蕾期低溫脅迫對蒸騰速率(Tr)的影響
研究表明,各低溫處理組的Tr均呈降低趨勢,各處理間差異顯著。同一低溫條件下,隨低溫處理時間的延長,3個蕾期棉花的Tr均顯著下降。各處理中,新陸早61號的Tr值隨處理時間延長降幅顯著低于其余3個品種,而新陸早36號的Tr值下降速度最快;T2(15℃/10℃)低溫下相對新陸早36號的Tr值最低,新陸早61號最高;T3(10℃/5℃)低溫下相對其它種質新陸早10號的Tr值最低,新陸早36號最高。在T2(15℃/10℃)低溫脅迫10 d 時,新陸早36號Tr值出現一個躍升,顯著高于其他2個品種。T1、T2低溫脅迫下新陸早61號的抗寒性較強,T3低溫脅迫下新陸早36號抗寒性較強,新陸早10號抗寒性最弱。Tr于T3(10℃/5℃)低溫脅迫10 d 和15 d 下均表現為緩慢下降。時間和溫度均對蕾期棉花的冷害起較重要的效應作用,本研究中蕾期棉花的Tr值極顯著(P<0.01)受“溫度×時間”互作的影響,且時間和溫度對不同蕾期棉花品種的危害效應同于上述Pn和Gs。圖5

圖5 低溫脅迫下蒸騰速率(Tr)變化
2.5 蕾期低溫脅迫對水分利用效率(WUE)影響
研究表明,水分利用效率WUE是植物生產中單位水分的能量轉化率指標。T1低溫脅迫處理時,3個品種的蕾期棉花葉片水分利用效率增大, T2處理時先升高后下降、T3處理時降低。低溫脅迫強度大于T2及低溫脅迫時間超過10 d時蕾期棉花可造成較大的影響。圖6

圖6 低溫脅迫下水分利用效率(WUE)變化
T1處理的10 d、 T2處理的5 d、T3處理的10 d、15 d均與CK之間差異顯著。T1、T2處理5 d時,新陸早61號、新陸早36號發生了較大的躍升,推測是新陸早61號、新陸早36號可通過提升水分利用效率(WUE)來增強植株的抗冷性,推測T1、T2處理5 d內為蕾期棉花植株可調節恢復的范圍。T3處理下的蕾期棉花的水分利用效率(WUE)表現為急劇降低,當T3脅迫15 d時為負值,表明T3的低溫脅迫處理對蕾期棉花可造成較大的影響,且脅迫超過10 d可造成植株死亡。當T1、T2處理持續0~5 d時,WUE不斷增加,即葉片WUE在T1、T2處理持續5 d時達到最大,之后開始迅速下降。T1、T2處理持續5 d是蕾期棉花受低溫脅迫的極限, T3脅迫強度或 T1、T2脅迫強度持續5 d以上,都會使葉片無法通過調節氣孔的開張和蒸騰速率緩解低溫對植株造成的傷害。圖6
研究表明蕾期棉花在輕度(T1-20℃/15℃)低溫脅迫下水分利用率(WUE)呈上升趨勢的現象一致,表明蕾期棉花可在輕度(T1-20℃/15℃)低溫脅迫下存活。中度(T2-15℃/10℃)下,處理前5 d水分利用率(WUE)升高,而處理持續時間超過5 d后水分利用率(WUE)下降,中度(T2-15℃/10℃)低溫脅迫持續5 d是兩蕾期棉花的生存閾值。
2.6 蕾期低溫脅迫對氣孔限制值(Ls)的影響
研究表明,T1處理時,處理5d時的氣孔限制值Ls下降比較急劇,低溫脅迫初期會迫使蕾期棉花葉片對光能的利用效率急劇下降。T1處理過程中表現為降升降的趨勢。T1和T3處理Ls峰值出現在第10 d,T1處理5 d新陸早10號、新陸早36號、新陸早61號氣孔限制值分別為0.205、0.068、0.099;T2處理5 d新陸早10號、新陸早36號、新陸早61號氣孔限制值分別為0.270、0.570、0.236; T3處理5 d新陸早10號、新陸早36號、新陸早61號氣孔限制值分別為0.256、0.063、0.228,各處理間差異不顯著(P>0.05)。圖7
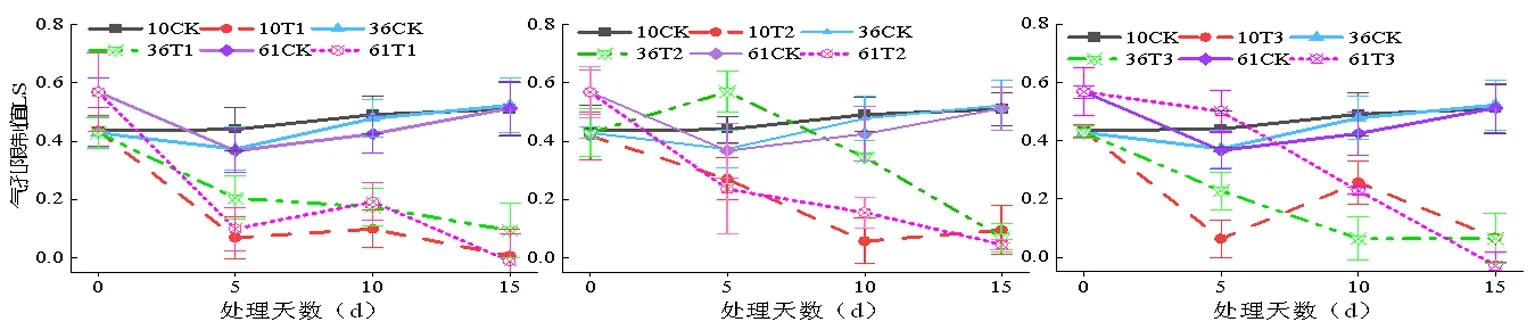
圖7 低溫脅迫下水分利用效率(WUE)變化
T1和T2處理下新陸早36號的氣孔限制值Ls受影響最小,T3處理則新陸早61號的氣孔限制值Ls表現為受影響最小。而T1、T2和T3處理的新陸早10號的氣孔限制值Ls都表現為受的影響最大。新陸早10號品種的Ls平均值在T1、T2和T3脅迫下分別比CK組下降了53.41% 、38.64%和41.82%。蕾期棉花在遭受低溫脅迫時會表現出Ls大幅度降低。同一低溫脅迫強度處理下,隨著低溫處理時間的延長,蕾期棉花氣孔限制值Ls越小,與CK處理組的差異越來越大。低溫脅迫時非氣孔限制是導致蕾期棉花光合速率降低的主要因素[20]。
3 討 論
Pn是光合系統功能的直接體現,大多數的植物在遭遇低溫脅迫時凈光合速率Pn都表現出明顯下降[21]。不僅環境因子和氣孔導度會導致Pn下降,非氣孔限制因素也是可能降低Pn的因素。植物的光合與蒸騰受氣孔的制約,氣孔對植物與大氣進行水汽和 CO2交換起到很重要的作用[23]。
氣孔限制因素引起的Pn下降表現為Gs、Tr下降,Ci也相應下降,非氣孔限制因素引起的Pn下降表現為Gs、Tr下降的同時Ci升高[24-25];這與研究中氣孔導度Gs、凈光合效率Pn下降時Ci升高的結果一致,表明本研究中限制蕾期棉花Pn的主要原因是非氣孔因素,與郭菊蘭等[26]表明非氣孔因子限制是迫使秋茄幼苗凈光合速率Pn下降的主要因素。蕾期棉花相同脅迫時間下,T1、T2的溫度處理Pn下降主要受氣孔限制因素引起光合作用受限制所致,而T3以下低溫處理Pn下降很大程度來源于非氣孔限制因素。
低溫脅迫下,凈光合速率(Pn)、蒸騰速率(Tr)、氣孔導度(Gs)均逐步下降,低溫脅迫越重,光合參數下降幅度越大;重度低溫已嚴重損害葉片光合機構,極大抑制了光合作用[22]。與研究中蕾期棉花在低溫脅迫后, 光合速率(Pn)、蒸騰速率(Tr)、氣孔導度(Gs)均比對照明顯下降。且下降趨勢隨脅迫程度加大而增大,表現為T3>T2>T1的結果相一致。研究發現蕾期棉花在中度低溫脅迫(T2-15℃/10℃)下處理超過5 d及重度(T3-10℃/5℃)低溫脅迫下水分利用率(WUE)呈下降趨勢。與鄭芊彤等[18]對三七進行低溫處理7 d后發現重度處理的WUE先上升后下降,輕度處理的WUE隨持續時間的延長而上升的結果一致。水分利用率(WUE)的變化趨勢不僅取決于脅迫的溫度,還與脅迫持續時長有著密切的關聯。
試驗中蕾期棉花隨低溫脅迫時間的延長,Ls呈下降趨勢,低溫環境使得蕾期棉花葉片對光能的利用效率下降,與許耀照等[16]表明T2(15℃/8℃)與T1(19℃/12℃)低溫處理下氣孔限制值的降低程度達到顯著差異的研究結果一致。
4 結 論
4.1可根據蕾期棉花的氣孔導度(Gs)、蒸騰速率(Tr)、細胞間二氧化碳濃度(Ci)、水分利用率(WUE)來判斷其受到的低溫脅迫程度。輕度脅迫下蕾期棉花光合速率降低是由氣孔限制因素導致;中度脅迫下蕾期棉花光合速率降低的主要原因先為氣孔因素后轉為非氣孔因素;重度脅迫下非氣孔因素是限制蕾期棉花光合作用的主要因素。
4.2低溫脅迫對蕾期葉片的光合作用有抑制的作用。輕度脅迫時新陸早61號的耐低溫性較好,中度、重度脅迫時新陸早36號相比于新陸早61號和新陸早10號耐低溫性更好。脅迫溫度越低,對蕾期棉花生長的抑制作用越明顯,且相同低溫度值處理下,低溫處理的時間持續越長,對蕾期棉花生長的抑制作用越明顯。
