打頂對黃精種子生長生理及萌發特性的影響
2022-08-11 06:34:50張紅瑞孟盼盼劉國彬何立威高致明
江蘇農業科學 2022年15期
張紅瑞,孟盼盼,劉國彬,何立威,高致明,王 茜
(1.河南農業大學農學院,河南鄭州 450046;2.河南省三門峽市農業技術推廣站,河南三門峽 472000;3.河南省農業科學院長垣分院,河南長垣 453400)
黃精(Red.)為百合科黃精屬多年生草本植物,是中藥黃精的基源植物之一。黃精以根莖入藥,始載于《名醫別錄》,曰:“黃精,味甘,平,無毒。主補中益氣,除風濕,安五臟。久服輕身、延年、不饑”。傳統中醫認為黃精性甘、味平;歸脾、肺、腎經;具有補氣養陰、健脾、潤肺、益腎的功效。現代研究表明,黃精含有多糖、低聚糖、黃酮、皂苷、生物堿、揮發油等,具有降血糖、降血脂、抗腫瘤、抗脂肪肝、保護心臟、保護腎臟、抗氧化、抗阿爾茨海默癥等藥理作用。近年來,隨著人們保健意識的增強,黃精作為藥食同源中藥材需求量逐漸增大,野生資源無法滿足市場需求,栽培資源逐漸成為主要商品來源。研究發現,種子繁殖具有繁殖系數高的優勢,黃精為無限花序,花開放時間不一致,一般為先下后上,種子成熟度不一致,質量參差不齊,又具有休眠特性,自然狀態下種子繁殖萌發期較長。打頂是促進作物優質高產的重要措施,有關黃精打頂方面的研究鮮有報道。本試驗研究了打頂對黃精種子生理及萌發特性的影響,以期明確打頂對黃精種子發育進程的影響及適宜打頂的葉位,為黃精良種繁育提供依據。
1 材料與方法
1.1 試驗材料
試驗于2020年在河南省洛陽市嵩縣黃精種植基地(112.11°E,33.80°N)進行,海拔670 m。以栽培3年的黃精為試驗材料,經河南農業大學高致明教授鑒定,為百合科黃精屬植物黃精(Red.)。
1.2 試驗設計
選栽培3年、長勢均勻、無病蟲害的黃精,現蕾期標記莖粗(8±1)mm的植株200株,由于黃精一般在植株從下往上數第3輪葉開始開花結實,并隨著葉位升高結果數逐漸降低,分別在第4輪、第6輪、第8輪、第10輪葉位上方約1 cm處打頂,以不打頂為對照,每處理40株。7月31日開始第1次采種取樣(整株采收),每15 d取1次樣,每次取樣每處理隨機抽取5株進行采種。
1.3 測定項目及方法
用于酶活性測定的樣品采收后立即保存于液氮,其余分別用信封保存備用。測定完百果質量和果實直徑后搓去果肉洗凈,吸干種子表面水分,測定鮮黃精種子千粒質量及可溶性糖、淀粉、脂肪含量。用培養皿法進行發芽試驗,用穴盤在自然環境下育苗。冷凍樣品用于測定超氧化物歧化酶(superoxide dismutase,SOD)、過氧化物酶(peroxidase,POD)、過氧化氫酶(catalase,CAT)活性及可溶性蛋白含量。
1.3.1 黃精果實及種子農藝性狀的測定 果實和種子顏色參考前人研究,采用目測法劃分。每個處理隨機抽取100顆黃精果實稱質量,重復3次,計算百果質量。用游標卡尺測量垂直于果柄和柱頭的連線,3次重復,每個重復隨機抽取10顆果實測量后取平均值,計算果實直徑。采用百粒法測定黃精種子千粒質量。
1.3.2 黃精種子能量物質的測定 可溶性蛋白含量用考馬斯亮藍法測定;可溶性糖含量用蒽酮-硫酸法測定;淀粉含量用蒽酮比色法測定;粗脂肪含量用油重法(索氏提取法)測定。
1.3.3 黃精種子生理指標的測定 SOD活性使用氮藍四唑(NBT)光化還原法測定;POD活性用愈創木酚法測定;CAT活性用紫外吸收法測定。稱取新鮮黃精種子5 g左右,105 ℃下烘至恒質量,稱質量,重復3次,計算含水率,含水率=(鮮質量-干質量)/鮮質量×100%。
1.3.4 發芽參數的測定 將黃精果實搓去果肉,洗凈,隨機數30粒種子,均勻擺放于鋪有濕潤的雙層濾紙的培養皿中。每個處理3次重復,每個重復30粒種子,用培養皿法進行發芽試驗。將培養皿置于恒溫恒濕培養箱內,溫度(26±0.5)℃,每4 d統計發芽種子數(胚根突破種皮2 mm視為發芽)、初生根莖數(胚芽基部有明顯的膨大視為形成初生根莖)、發霉種子數。發芽時間為60 d。發芽率=發芽種子數/30×100%,發芽勢=40 d內發芽的種子總數/30×100%,發芽指數=∑(當天發芽種子數/發芽日數)。
1.3.5 出苗率的測定 每次樣品搓去果肉洗凈后,隨機數50粒種子,播種在穴寬5 cm、每盤50穴的穴盤中,重復3次,基質選用品氏基質。穴盤置于林下并覆蓋地膜,翌年3月下旬統計出苗率。出苗率=出苗數/50×100%。
1.4 數據分析
數據主要采用Excel 2016和SPSS 25.0進行統計分析。
2 結果與分析
2.1 打頂對黃精果實和種子農藝性狀的影響
2.1.1 打頂對黃精果實百果質量的影響 由圖1可知,不同時期、不同打頂高度對黃精果實百果質量均有影響。7月31日第10輪(65.40 g)、第8輪(54.39 g)、第6輪(52.15 g)葉處打頂的百果質量極顯著大于對照(48.13 g)。8月15日僅第10輪(61.31 g)葉處打頂百果質量極顯著大于對照(52.40 g)。8月30日第10輪(63.72 g)和第8輪(59.84 g)葉處打頂百果質量極顯著大于對照(52.46 g)。9月14日第10輪(62.99 g)和第8輪(62.40 g)葉處打頂百果質量與對照(62.05 g)無顯著差異,但均極顯著大于第6輪(55.38 g)和第4輪(54.92 g)葉打頂。9月29日不同打頂高度與對照均無顯著差異。可知,在黃精第10輪葉處打頂能提高8月30日前果實百果質量,截去的黃精葉輪數越多其果實百果質量越小。
2.1.2 打頂對黃精果實直徑的影響 由圖2可知,黃精果實直徑在各個時間變化不大。7月31日第8輪葉處打頂果實直徑最大,為11.33 mm,極顯著大于對照(10.27 mm)和第10輪(10.16 mm)、第6輪(10.53 mm)葉處打頂;8月15日對照果實直徑最大,為11.07 mm,極顯著大于第6輪(10.08 mm)葉處打頂且顯著高于另外3個打頂高度;8月30日、9月14日、9月29日3個時期不同打頂程度之間均無顯著差異性。說明打頂僅在8月15日前能增大果實直徑,且第8輪葉處打頂最明顯。


2.1.3 打頂對黃精種子千粒質量的影響 由圖3可知,黃精種子千粒質量隨生長時間變久而變大。在第10輪和第8輪葉處打頂黃精種子千粒質量較大,大于對照和其他打頂高度。7月31日第10輪、第8輪葉處打頂種子千粒質量分別為40.76、40.73 g,極顯著高于其他處理,第4輪葉處打頂黃精種子千粒質量(34.47 g)顯著小于對照(36.60 g)。8月15日第10輪、第8輪葉處打頂黃精種子千粒質量分別為46.93、46.73 g,差異不顯著,但都極顯著大于其他打頂高度及對照(40.57 g)。8月30日第10輪葉處打頂黃精種子千粒質量最大,為47.57 g,極顯著大于對照(43.23 g)和第6輪(41.20 g)、第4輪(41.73 g)葉處打頂;第8輪葉處打頂黃精種子千粒質量為45.20 g,極顯著大于第6輪和第4輪葉處打頂。9月14日第10輪葉處打頂黃精種子千粒質量最大,為48.13 g,顯著大于對照(44.97 g)并極顯著大于其他打頂高度。9月29日仍為第10輪葉處打頂黃精種子千粒質量最大,為49.27 g,極顯著大于對照(46.63 g)和第6輪(44.00 g)、第4輪(42.93 g)葉處打頂,與第8輪(48.63 g)葉處打頂差異不顯著。從千粒質量來看打頂高度應在第8~10輪葉處。
2.2 打頂對黃精種子物質積累的影響
2.2.1 打頂對黃精種子可溶性糖含量的影響 由圖4可知,7月31日第10輪葉處打頂的可溶性糖含量(30.95 mg/g)極顯著高于對照(27.24 mg/g),第8輪(28.04 mg/g)葉處打頂與對照無顯著差異,第6輪(23.86 mg/g)和4輪(20.83 mg/g)葉處打頂的都極顯著低于對照,且極顯著低于第10輪和第8輪。8月15日所有打頂處理的黃精種子可溶性糖含量均顯著低于對照(27.58 mg/g),且第8輪(24.54 mg/g)、6輪(23.65 mg/g)、4輪(17.95 mg/g)葉處打頂極顯著低于對照。8月30日第10輪葉處打頂的黃精種子可溶性糖含量(20.10 mg/g)與對照(20.86 mg/g)之間無顯著差異,但顯著高于第8輪(17.38 mg/g)、6輪(17.98 mg/g)、4輪(17.92 mg/g)葉處打頂處理。9月14日只有第4輪葉處打頂的黃精種子可溶性糖含量極顯著低于對照和其他處理,其他處理間及與對照均無顯著差異。9月29日對照與處理間均無顯著差異。


2.2.2 打頂對黃精種子可溶性蛋白含量的影響 由圖5可知,7月31日不同打頂處理黃精種子可溶性蛋白含量無顯著差異。8月15日不同打頂處理黃精種子表現為4輪(142.58 mg/g)>6輪(130.93 mg/g)>8輪(127.36 mg/g)>10輪(115.62 mg/g)>對照(110.06 mg/g),從第4輪葉處打頂的黃精種子顯著高于其他處理,第6輪和8輪葉處打頂高于第10輪葉處和對照,但未達到顯著水平。8月30日亦是第4輪打頂的可溶性蛋白含量最高,顯著高于其他處理,且極顯著高于對照。9月14日和9月29日不同打頂處理黃精種子的可溶性蛋白含量無顯著差異。

2.2.3 打頂對黃精種子淀粉含量的影響 從圖6可以看出,7月31日對照(141.57 mg/g)與第10輪葉處打頂黃精種子淀粉含量(141.00 mg/g)無顯著差異但都極顯著高于第6輪(119.94 mg/g)、第4輪(118.09 mg/g)葉處打頂。8月15日對照黃精種子淀粉含量(145.28 mg/g)顯著高于第10輪(136.45 mg/g)、第8輪(133.68 mg/g)葉處打頂,極顯著高于第6輪(126.18 mg/g)、第4輪(123.44 mg/g)葉處打頂。9月14日第10輪葉處打頂黃精種子淀粉含量(67.37 mg/g)最低,極顯著低于對照(91.23 mg/g)、第6輪(81.53 mg/g)和第4輪(92.27 mg/g)葉處打頂。8月30日、9月29日各處理間淀粉含量無顯著差異。
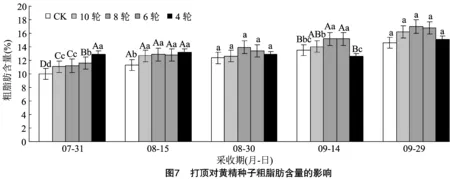
2.2.4 打頂對黃精種子粗脂肪含量的影響 由圖7可以看出,7月31日處理組黃精種子粗脂肪含量均極顯著高于對照,除第10輪和第8輪葉處打頂處理,其他均兩兩間有極顯著差異性,表現為第4輪葉處打頂(12.94%)>6輪(11.57%)>8輪(11.20%)>10輪(11.05%)>對照(10.00%)。8月15日處理組均顯著高于對照,但處理間無顯著差異。8月30日、9月29日不同打頂處理黃精種子粗脂肪含量均無顯著差異。9月14日,第8輪(15.15%)、6輪(15.15%)葉處打頂的黃精所結種子粗脂肪含量極顯著高于對照(13.54%),第10輪(14.02%)葉處打頂高于對照,第4輪(12.55%)葉處打頂低于對照,但均未表現出差異性。

2.3 打頂對黃精種子生理指標的影響
2.3.1 打頂對黃精種子SOD活性的影響 由圖8可以看出,黃精種子SOD活性先升高后降低,在第4輪葉處打頂于9月14日達到峰值,其余處理均在8月30日達到峰值。7月31日第4輪葉處打頂(91.02 U/g)黃精種子SOD活性顯著高于第10輪(72.63 U/g),極顯著高于對照(68.80 U/g),第6輪葉處打頂(87.24 U/g)顯著高于第10輪和對照。8月15日第4輪葉處打頂(234.08 U/g)極顯著高于對照(131.29 U/g)及其他打頂處理,第6輪(172.49 U/g)和第8輪葉處打頂(165.62 U/g)極顯著高于對照,顯著高于第10輪葉處打頂(142.17 U/g)。8月30日對照(705.49 U/g)和第10輪葉處打頂(745.51 U/g)黃精種子SOD活性極顯著高于其他打頂處理,第8輪(613.54 U/g)、第6輪葉處打頂(588.65 U/g)極顯著高于第4輪打頂(432.78 U/g)。9月14日第4輪葉處打頂(459.23 U/g)黃精種子SOD活性顯著高于對照(359.18 U/g)和第10輪(380.38 U/g)。9月29日第4輪葉處打頂(303.69 U/g)黃精種子SOD活性極顯著高于對照(174.94 U/g)和第10輪(229.00 U/g),打頂處理均顯著高于對照。

2.3.2 打頂對黃精種子POD活性的影響 由圖9可以看出,黃精種子POD活性先升高后降低再升高,不同打頂高度達到峰值的時間略有不同。7月31日打頂處理黃精種子POD活性與對照均無顯著差異,第6輪葉處打頂(4.56 U/g)顯著高于第10輪(3.37 U/g)。8月15日打頂處理黃精種子的POD活性均高于對照(5.01 U/g),第6輪葉處打頂(6.63 U/g)達到顯著水平,第4輪(9.19 U/g)達到極顯著水平。8月30日以第8輪葉處打頂黃精種子POD活性最高,為8.81 U/g,極顯著高于對照(6.02 U/g)及其他處理水平,第10輪(6.76 U/g)、6輪(6.41 U/g)葉處打頂所得黃精種子的POD活性與對照無顯著差異,而第4輪葉處打頂(4.61 U/g)黃精種子POD活性極顯著低于對照。9月14日打頂黃精種子POD活性均低于對照(6.90 U/g),且第6輪(4.83 U/g)、4輪(4.76 U/g)葉處打頂的黃精種子POD活性達到顯著水平。9月29日不同打頂處理之間及與對照之間均無顯著性差異。

2.3.3 打頂對黃精種子CAT活性的影響 由圖10可知,黃精種子CAT活性總體呈先升后降趨勢。7月31日打頂處理黃精種子CAT活性均極顯著高于對照,且第4輪葉處打頂(408.89 U/g)極顯著高于其他處理。8月15日黃精種子CAT活性為4輪(496.67 U/g)>6輪(491.11 U/g)>8輪(422.22 U/g)>10輪(324.44 U/g)>對照(262.22 U/g),打頂處理種子CAT活性均顯著高于對照,且8輪、6輪、4輪達到極顯著水平。8月30日打頂處理仍然均顯著高于對照(435.56 U/g),而第6輪(608.89 U/g)和4輪(620.00 U/g)葉處打頂種子CAT活性均極顯著低于第8輪(706.67 U/g)。9月14日第4輪葉處打頂種子CAT活性與對照(462.22 U/g)無顯著差異,且均極顯著低于第10輪(673.33 U/g)、8輪(666.67 U/g)、6輪(704.44 U/g)。9月29日各處理黃精種子CAT活性為對照(677.78 U/g)>10輪(593.33 U/g)>8輪(560.00 U/g)>6輪(526.67 U/g)>4輪(488.89 U/g),打頂處理均顯著低于對照,且第4~8輪葉處打頂達到極顯著水平。

2.3.4 打頂對黃精種子含水率的影響 由圖11可知,黃精種子含水率總體呈下降趨勢。7月31日對照黃精種子含水率最高,為65.57%,極顯著高于打頂處理,第10輪(63.56 g)、第8輪(63.11 g)葉處打頂黃精種子含水率顯著高于第4輪(61.70 g),第10輪葉處打頂達到極顯著水平,第6輪(62.59 g)葉處打頂與第4輪葉處打頂黃精種子含水率無顯著差異。8月15日不同打頂處理與對照均無顯著差異。8月30日對照黃精種子含水率最高,為44.02%,顯著高于第6輪(41.71 g)、第8輪(42.21 g)葉處打頂,且極顯著高于第4輪(39.85 g)。9月14日和9月29日不同打頂處理和對照均無顯著差異。

2.4 打頂對黃精種子萌發特性的影響
從表1可以看出,在7月31日打頂處理黃精種子發芽率均高于對照,且隨著打頂葉位降低發芽率呈升高趨勢;第8輪、第6輪、第4輪葉處打頂均顯著高于對照,且第4輪達到極顯著水平。8月15日對照發芽率明顯升高,這與不同時期黃精種子的發芽規律相同。8月30日打頂處理黃精種子比之上一時期發芽率更低,且此時對照的種子發芽率也開始降低。9月14日所有打頂處理所得的黃精種子發芽率又有所提高,且極顯著高于對照。9月29日對照種子發芽率升高幅度較小,而打頂處理黃精種子發芽率回升幅度較大。

表1 打頂對黃精種子萌發特性的影響
7月31日以第10輪葉處打頂黃精種子發芽指數(3.79)最高,顯著高于對照和其他打頂處理。8月15日各處理與對照均無顯著差異,但第8輪葉處打頂黃精種子發芽指數極顯著高于第10輪、4輪。8月30日各處理間及與對照均無顯著差異。9月14日對照黃精種子發芽指數顯著高于第10輪、8輪葉處打頂。9月29日不同打頂處理與對照間的發芽指數均無顯著差異。
前4個時期不同處理間發芽勢與對照相比均無顯著差異。9月29日第8輪葉處打頂的黃精種子發芽勢最高,顯著高于對照,其他處理間及與對照均無顯著差異。
7月31日打頂處理黃精種子初生根莖形成率均顯著高于對照,不同處理間無顯著差異,這與打頂處理后黃精種子發芽率高有關。8月15日除第10輪葉處打頂外初生根莖形成率均顯著低于對照,且第4輪葉處打頂達到極顯著水平。8月30日各打頂處理與對照均無顯著差異。9月14日第4輪葉處打頂初生根莖形成率與對照間形成顯著差異,其余處理與對照間差異均不顯著。9月29日打頂處理初生根莖形成率均高于對照,且第8輪葉處打頂達到極顯著水平,同時,第8輪葉處打頂初生根莖形成率極顯著高于第10輪、顯著高于第6輪葉處打頂,說明通過打頂可以改變黃精種子在不同時期初生根莖形成率,提高第2年出苗的潛力。
從發霉率來看,7月31日黃精種子發霉率隨著打頂葉位降低呈下降趨勢,對照最高(47.78%),顯著高于第10輪(36.67%)、8輪(35.56%)葉處打頂,極顯著高于第6輪(30.00%)、4輪(26.67%)葉處打頂。8月15日為對照種子發霉率最高(24.44%),第10輪葉處打頂(18.67%)次之,兩者之間無顯著差異,第8輪(14.44%)、第6輪(13.33%)、第4輪(14.67%)葉處打頂的種子發霉率均顯著低于對照。其后3個時期,不同處理間及與對照相比均無顯著差異,說明打頂可以降低7月底至8月中旬采收的黃精種子發霉率。
3 結論與討論
打頂處理可以促進黃精種子的發育進程,在第8~10輪葉處打頂較為適宜。第10輪、第8輪葉處打頂能顯著增高7月31日采收的黃精果實百果質量和種子千粒質量;打頂對黃精果實直徑影響不大;8月30日前打頂葉位越低種子含水率、可溶性糖含量、淀粉含量越低,可溶性蛋白含量、粗脂肪含量越高,8月30日后差別不大;不同時期黃精種子SOD、POD、CAT活性均呈先升后降的趨勢,打頂可使POD、CAT活性提前達到峰值;打頂對黃精種子的萌發特性有顯著影響,7月31日打頂葉位越低發芽率越高,8月15日打頂葉位越低發芽率越低,9月14日和9月29日打頂黃精種子發芽率均高于對照。打頂可以提高7月底至8月中旬以前采收的黃精種子發芽率、發芽勢、初生根莖形成率,降低發霉率。打頂葉位過高促進作用不明顯,打頂葉位過低造成葉片過少,光合速率降低,不利于黃精種子的發育。種植3年的黃精宜在第 8~10輪葉處打頂,可使種子提前達到生理成熟,采收期提前,為黃精種子當年發芽爭取高溫時間。
打頂作為一項常用的栽培措施,對多種作物的優質高產意義重大。打頂的目的主要有控制高度、促進側枝發育、控制開花時間、促進坐果等,其本質都是消除頂端優勢,調控源-庫關系。目前有關黃精打頂的研究較少,王東輝研究認為,黃精葉片的凈光合速率隨葉位的變化呈單峰曲線,第11輪葉片凈光合速率最高,葉位高于17輪葉時葉片光合速率顯著下降,建議去頂以利于黃精地下部分生長。而本研究發現,打頂還可促進種子的發育,和抗倒伏作用明顯。楊清平等對多花黃精進行研究發現,摘花和打頂措施可以調控其庫-源關系,實現光合產物和養分的定向運輸,能顯著提高多花黃精一年生塊莖生物量、生長速度。中藥材育種也常通過打頂來改善種子質量,如桔梗、肉蓯蓉、甘草、丹參等,打頂不僅能促進黃精種子發育,適宜高度打頂也可以防止倒伏和促進地下部分生長。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國生殖健康(2019年2期)2019-08-23 08:12:08
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
產品可靠性報告(2017年7期)2017-09-05 09:49:12
汽車觀察(2016年3期)2016-02-28 13:16:26
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
