鼠傷寒沙門氏菌感染機制及其檢測方法研究進展
2022-08-11 06:11:46吳婭芳王英林田亞晨黃志強姜嘉燁黃笑天馬文文
工業微生物 2022年3期
吳婭芳, 王英林, 田亞晨, 黃志強, 姜嘉燁, 黃笑天, 馬文文, 劉 箐
上海理工大學健康科學與工程學院,上海 200093
沙門氏菌是一種常見的食源性致病菌,在世界范圍內的細菌性食物中毒事件中,由沙門氏菌引起的食物中毒死亡人數高達3 300萬[1]。據世界衛生組織報告,沙門氏菌感染是造成腹瀉病的主要原因之一,每年約有5.5億人患病[2]。沙門氏菌分布十分廣泛,常寄居于蔬菜、蛋類、禽類肉制品等食品中。目前已鑒定的沙門氏菌血清型超過2 650多種[3],且從食品分離株中最常見的血清型是鼠傷寒沙門氏菌(Salmonellatyphimurium)[4-5]。近年來,鼠傷寒沙門氏菌感染禽類肉制品問題愈加嚴重,該致病菌不僅阻礙畜牧業的發展,而且危害人類健康。通過對鼠傷寒沙門氏菌逃避先天免疫機制及其在宿主間轉移、擴大感染等方面進行研究,可以為預防鼠傷寒沙門氏菌感染以及疫苗、藥物研發提供新的思路。此外,鼠傷寒沙門氏菌是食源性致病菌重點監控對象之一[6],對食品中鼠傷寒沙門氏菌進行準確、快速檢測也是預防和控制沙門氏菌的有效措施。隨著光譜、納米等技術的發展,各種高通量、高靈敏的檢測方法也應運而生,為實現食品中鼠傷寒沙門氏菌的快速檢測提供了新的手段。因此,本綜述主要從鼠傷寒沙門氏菌感染機制及其檢測技術兩個方面進行闡述,以期為鼠傷寒沙門氏菌的防治提供研究基礎與方向。
1 鼠傷寒沙門氏菌感染機制
1.1 鼠傷寒沙門氏菌逃避先天免疫的機制
鼠傷寒沙門氏菌通過逃避宿主防御系統的生存機制來對抗宿主攻擊,從而增加發病率和死亡率。黏附在宿主表面的鼠傷寒沙門氏菌利用宿主的信號通路對宿主細胞骨架結構重排并破壞正常的上皮細胞刷狀邊界,誘導膜皺褶的形成[7]。膜皺褶可以將病原體內吞到一個低酸堿度和營養缺乏的隔室中,即SCV(Salmonellacontaining vacuole)。鼠傷寒沙門氏菌分泌蛋白SopB已被證明可以激活細胞膜上的宿主激酶蛋白AKT1,進而磷酸化AS160。而AS160是一種潛在的rab14-GTPase激活蛋白,其磷酸化形式無法與吞噬體膜結合,rab14在SCV中保持激活狀態,從而抑制SCV與溶酶體融合[8]。入侵之后,鼠傷寒沙門氏菌毒力島-2(SPT-2)編碼的T3SS(type III secretion system)可以轉移SCV,將其定位在緊鄰高爾基體的宿主細胞核周區,有助于菌體攔截宿主內吞作用和胞吐囊泡作用,以便獲取營養物質和膜片段[9]。此外,SPI-2 T3SS在SCV內被誘導,可以防止抗微生物宿主因子輸送到SCV,改變宿主細胞骨架組織,影響囊泡運輸,維持SCV的完整性,從而保證沙門氏菌在宿主細胞內始終被SCV包裹著并進行生存、繁殖[10]。
1.2 鼠傷寒沙門氏菌感染與宿主之間的關系
1.2.1調節宿主細胞的遷移
SseI是由侵入細胞內的鼠傷寒沙門氏菌表達的一種分泌型效應物,通過SPI-2-T3SS轉運進入宿主細胞胞質溶膠[11]。有研究證實SseI是通過調節巨噬細胞和樹突細胞(DC)的遷移來維持小鼠長期全身性的感染[12]。而抑制遷移需要宿主因子含有GTPase激活蛋白1的IQ基序(IQGAP1),它是宿主細胞遷移的重要調節因子。當SseI與IQGAP1結合,就會抑制對激活T細胞至關重要的有效抗原呈遞細胞DC向趨化因子CCL19/21遷移(見圖1)[13]。雖然SseI調節遷移的確切機制仍不清楚,但它能夠阻斷宿主免疫細胞向T細胞的遷移,從而削弱了宿主清除全身細菌的能力。
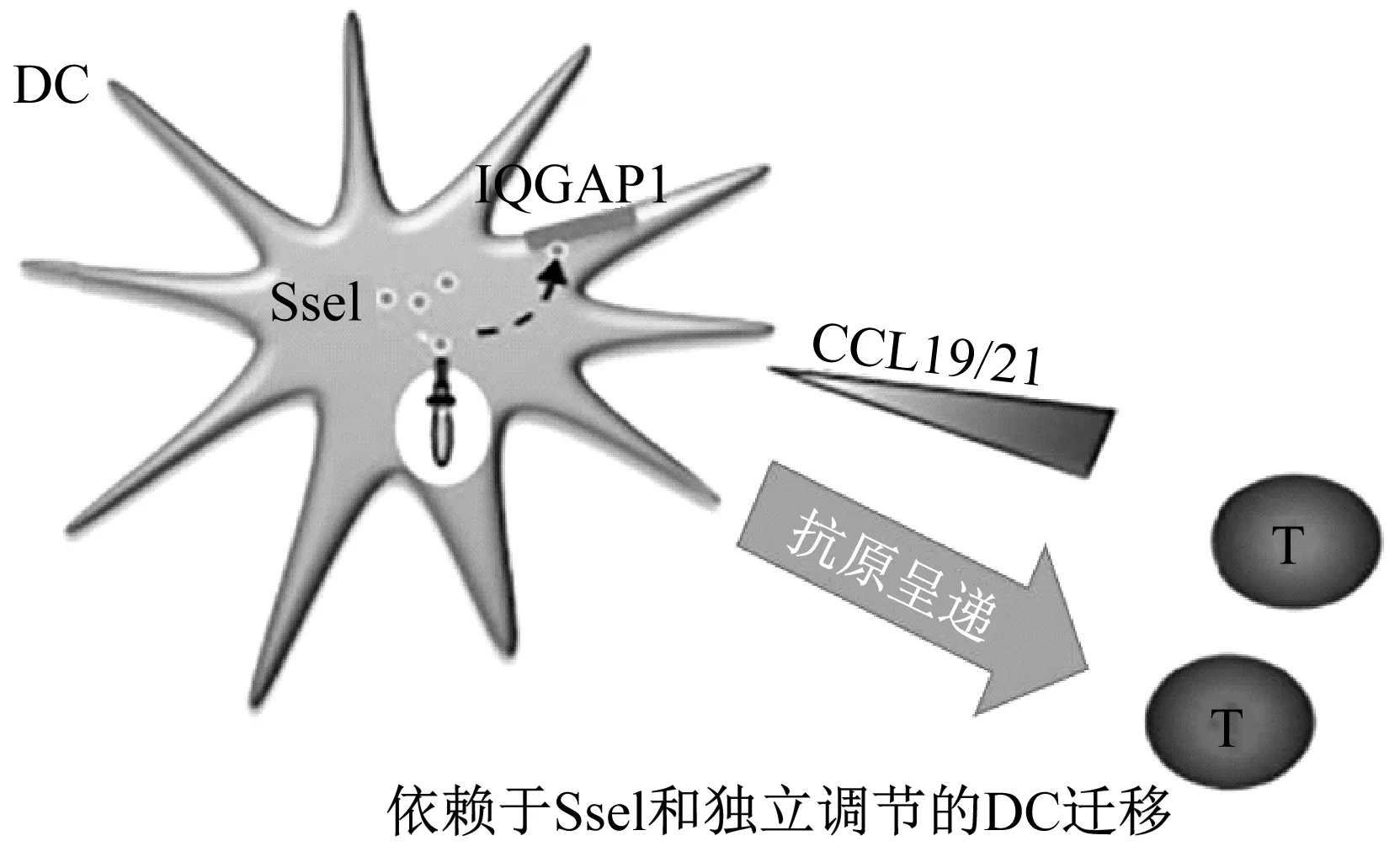
圖1 沙門氏菌抑制適應性免疫反應[13]
1.2.2宿主細胞的凋亡和鼠傷寒沙門氏菌的擴大感染
在宿主細胞感染初期,鼠傷寒沙門氏菌毒力島-1(SPI-1)編碼的SipB蛋白與細胞凋亡誘導的執行因子Caspase(cysteine protease)家族的Caspase-1結合,使其活化[14]。Caspase-1的活化不僅促使細胞凋亡,還是引起腸道組織炎癥的主要原因之一[15]。此外,有報道稱,如果沙門氏菌感染源自Caspase-1基因敲除小鼠的巨噬細胞,那么在感染初期Caspase-2也會誘發細胞凋亡[16]。Caspase-2的活化可能也和SipB蛋白有關。Caspase與SipB蛋白結合機制尚不清楚,有學者認為可能是SipB蛋白直接貫穿SCV膜與Caspase結合。在感染晚期,SPT-2的表達產物會誘導巨噬細胞的凋亡[17]。細胞凋亡發生在肝臟和脾臟,死細胞被相鄰的吞噬細胞吞噬,通過細胞內增殖進一步擴大了感染的范圍,所以細胞凋亡對于沙門氏菌擴大感染是必要的。
2 檢測方法研究進展
對鼠傷寒沙門氏菌簡單、快速、準確的檢測是預防和控制沙門氏菌感染的有效手段。目前,沙門氏菌的檢測方法主要包括傳統分離方法、免疫學方法、分子生物學方法以及近年來多學科交叉發展起來的新型檢測方法,如生物傳感器技術、光譜技術、納米技術等。這些方法為沙門氏菌快速、準確、靈敏的檢測提供了新的方向。本文對鼠傷寒沙門氏菌的檢測方法進行了概述,為鼠傷寒沙門氏菌的預防和監控以及新型檢測技術的研發提供一些參考。
2.1 傳統分離方法
傳統培養方法是根據病原菌的生長繁殖特性,利用選擇性培養基對細菌進行篩選和分離,再結合生物特性對其進行鑒定。根據(GB 4789.4—2016)《食品安全國家標準 食品衛生微生物學檢驗 沙門氏菌檢驗》[18],其具體流程包括8 h~18 h的預增菌、18 h~24 h的增菌、選擇性平板分離、生化試驗和血清學鑒定五個步驟。傳統分離方法準確性高,對儀器要求低,是鑒定鼠傷寒沙門氏菌的金標準。但是操作繁瑣,檢測周期一般長達4 d~7 d,不適用于即時現場檢測。細菌抗原表位的改變或丟失,也會導致同種血清型沙門氏菌的抗原性不同,降低檢測靈敏度,從而出現假陰性[19]。另外,由于存在細菌“活的不可培養”狀態(viable but non-culturable, VBNC)的情況,從而導致實驗結果出現假陰性[20]。因此,構建更加準確、快速、便捷的檢測方法對于鼠傷寒沙門氏菌的預防具有重大意義。
2.2 免疫學檢測技術
免疫學檢測技術是通過抗原抗體特異性結合來檢測目標致病菌。目前,該技術廣泛用于鼠傷寒沙門氏菌的檢測。根據檢測原理的差異性可以將免疫學檢測技術分為標記免疫學檢測技術和非標記免疫學檢測技術。大致分為酶聯免疫技術、熒光免疫技術、免疫磁珠分離技術和放射性免疫技術等。
2.2.1酶聯免疫吸附檢測法(Enzyme-Linked Immuno Sorbent Assay, ELISA)
酶聯免疫吸附技術是目前較為成熟的一種檢測方法,其基本原理是將抗體(抗原)吸附在固相載體表面,依次加入受檢樣品和酶標記的抗原(抗體),它們通過特異性結合形成抗原抗體復合物,用洗滌的方式將未結合的物質除去,然后加入酶反應底物,底物經酶催化變為有色物質,最后根據顏色的深淺對樣本進行定性或定量分析。該檢測方法周期較短、靈敏度高。WANG W等[21]制備針對鼠傷寒沙門氏菌鞭毛蛋白和脂多糖的單克隆抗體,建立了基于抗鼠傷寒沙門氏菌脂多糖的雙抗體夾心ELISA法對目標菌進行檢測,檢出限達到104CFU/mL。 CHUNGLOK W等[22]利用單壁碳納米管(SWCNTS)構建的標記平臺來固定辣根過氧化物酶(HRP)標記的鼠傷寒沙門氏菌抗體,形成的Ab/SWCNTs/HRP偶聯物用于ELISA試驗。該檢測方法的檢測靈敏度比傳統ELISA方法提高了1 000倍。
2.2.2免疫層析檢測技術(Immunochromatography Assay, ICA)
免疫層析法的基本原理是將特異性抗體固定在硝酸纖維素膜上的某個區域,然后將干燥的一端插入待檢樣品中,在毛細血管力的作用下,樣品向前移動,當到達固定有抗體的區域時,抗原與抗體發生特異性結合,若用膠體金或者酶染色,則會出現肉眼可見的條帶,從而得出檢測結果。相較于ELISA,免疫層析法操作簡單,方便快捷。WU Y X[23]等構建了一種能夠檢測沙門氏菌的廣譜性膠體金試紙條。將制備的抗鼠傷寒沙門氏菌單克隆抗體用膠體金標記,因為該抗體可以與自身配對,所以也用作捕獲抗體,從而形成一個三明治檢測平臺。該試紙條可以檢測18種沙門氏菌種,與其他14種食源性病原菌無交叉,對鼠傷寒沙門氏菌的檢測限達到4×105CFU/mL,具有較高的廣譜性和特異性,但其靈敏度有待提高。REN Y R[24]等開發了一種基于鹽誘導聚集金納米顆粒的免疫層析試紙條來檢測樣品中的鼠傷寒沙門氏菌,該方法的檢測線為103CFU/mL,靈敏度較上種提高了100倍。
2.3 分子生物學檢測技術
分子生物學檢測技術是通過檢測分子水平的核酸序列來橫向比較不同物種、同物種不同個體、同個體的不同細胞或不同生理(病理)狀態的差異[25]。隨著現代分子生物學技術的快速發展,人們能夠深入到分子層面對食源性致病菌進行診斷。常見的分子生物學檢測技術主要包括聚合酶鏈式反應(Polymerase Chain Reaction, PCR),環介導等溫擴增技術(loop-mediated isothermal amplification, LAMP),核酸探針技術和基因芯片技術等。這些檢測技術具有穩定的準確性、較高的靈敏度和特異性,被廣泛用于食源性致病菌的檢測。
2.3.1PCR
PCR技術是由Kary Mullis在1985年發明的一種體外擴增特定DNA片段的檢測方法,將微量DNA片段進行成倍增加。具體流程包括:變性、退火和延伸。常見的PCR方法包括實時熒光PCR技術、免疫PCR技術、多重PCR技術和逆轉錄PCR。XU J X[26]等利用多重PCR技術檢測肉類中的鼠傷寒沙門氏菌,檢測限達100 CFU/25 μL。相較于傳統培養方法,該技術檢測時間短,可同時檢測多種不同類型的致病菌。GWAK S H[27]等將過濾、DNA濃縮和實時PCR技術聯合使用,建立了一種快速檢測甘藍中鼠傷寒沙門氏菌的方法。這種聯合方法能夠在5 h內快速檢出食品中(1~4) CFU/25 g的活菌。因此,該方法有望促進分子生物學檢測技術的發展,從而實現快速檢測。
2.3.2LAMP
LAMP法是一種新型核酸擴增技術,其基本原理是對靶基因上的六個區域設計四條引物,在恒溫條件下,15 min~60 min內實現109~1010倍的擴增,通過肉眼觀察白色擴增產物即焦磷酸鎂白色沉淀的有無來判斷靶基因是否存在。與傳統PCR相比,不需要熱變性、溫度循環和電泳觀察等步驟。LAMP法具有快速、靈敏、準確性高等特點。DEHGHANI Z[28]等將鉑/鈀納米顆粒(Pt/Pd NP)和DNA適配體雙修飾的磁珠與LAMP技術相結合,來檢測食品中鼠傷寒沙門氏菌的含量。該方法可以快速檢測食品中的鼠傷寒沙門氏菌,檢測線低至(3~10) CFU/mL,為鼠傷寒沙門氏菌的檢測提供了一個全新的思路。CHUN H J[29]等將LAMP技術和后反射Janus粒子(RJP)相結合,開發了一種檢測鼠傷寒沙門氏菌的分子生物傳感器。RJPs作為光信號探針可以與LAMP產物結合,擴增的LAMP產物濃度與結合的RJPs數量成正比。因此,通過測量熒光強度可以得到鼠傷寒沙門氏菌的濃度,其檢測限為102CFU/mL。該方法具有高特異性、快速和簡便的特點,并為解決傳統LAMP不能定量的問題提供新的思路。
2.4 新型檢測技術
2.4.1生物傳感器檢測技術
生物傳感器,是一種對生物物質敏感并將其濃度轉換為電信號進行檢測的儀器。它主要由三個部分組成:識別元件、信號轉換器和放大器。識別元件由特定與所需要的分析物相互作用的生物材料組成(包括酶、抗體、抗原、微生物、細胞、組織、核酸等生物活性物質),信號轉化器(如氧電極、光敏管、場效應管、壓電晶體等)能夠將生物響應轉換為可測量的信號,產生的信號則由放大器進行放大[30]。生物傳感器檢測沙門氏菌是以抗原抗體的特異性識別、核酸或適配體對菌體的特異性識別為基礎,將生物信號放大并轉化為光學或者電化學信號,通過建立化學信號與沙門氏菌濃度之間的關系達到檢測的目的[31]。生物傳感器根據其結構上使用的識別元件以及其信號轉換器的不同進行分類,可以分為酶傳感器、免疫傳感器、電化學生物傳感器、基因傳感器、光敏傳感器等[32]。CHAI等[33]針對雞蛋殼表面的鼠傷寒沙門氏菌設計了一種噬菌體磁彈性(ME)生物傳感器。該傳感器根據細菌附著在傳感器表面后的共振頻率變化來檢測蛋殼上的鼠傷寒沙門氏菌,并在20 min內完成對蛋殼樣品表面的鼠傷寒沙門氏菌的檢測。SPR生物傳感器是一種利用表面等離子體共振實時監測抗體-抗原相互作用的光學檢測儀器。它通過分析物與固定在SPR傳感器上的生物識別分子相結合的方式導致折射率變化,由生物分子的含量與折射率的線性關系反映樣品中沙門氏菌的濃度[31]。BHANDARI D等[34]開發了一種基于SPR檢測萵苣葉中鼠傷寒沙門氏菌的傳感器,該傳感器利用了雙抗體夾心的原理,將針對鼠傷寒沙門氏菌鞭毛蛋白的單克隆抗體(MAb 1E10)固定在傳感器表面,然后用另一株捕獲抗體(MAb 1C8)與檢測樣品孵育半小時,孵育過程中形成鼠傷寒鞭毛蛋白-MAb 1C8復合物。將鞭毛蛋白-MAb 1C8復合物引入傳感器中進行培養,培養過程中固定在傳感器表面上的MAb 1E10會捕獲不同濃度的鞭毛蛋白-MAb 1C8復合物,從而構建鞭毛蛋白濃度與SPR響應的劑量關系曲線(見圖2[34])。該方法檢測人工污染的長葉萵苣樣品檢出4.7×105CFU/mL。在提高檢測靈敏性方面,XU Y等[35]設計了一種Ω型光纖局部SPR生物傳感器用于雞肉樣品中鼠傷寒沙門氏菌的快速檢測,檢測限達到128 CFU/mL,是一種靈敏度相對較高的檢測方法。

圖2 基于表面等離子體共振生物傳感器檢測鼠傷寒沙門氏菌示意圖[34]
2.4.2光譜技術
光譜技術,是指利用光與物質的相互作用研究分子結構及動態特性的一門技術,即通過獲取光的發射、吸收與散射信息來獲得與樣品相關的信息。基于光學原理的鼠傷寒沙門氏菌檢測是將光學材料與電子材料、生物學結合起來,利用如抗體、適配體等特定識別分子來分析待測樣品,以材料光學特性的變化反映細菌濃度的變化,實現對鼠傷寒沙門氏菌的檢測。光學材料應用于食品中致病菌檢測方面的技術包括光學生物傳感器技術、熒光納米免疫層析試紙條技術等。將光譜技術與常規檢測技術相結合,具有便捷,靈敏度高的優點。SRINIVASAN S等[36]開發了一種基于熒光篩選的鼠傷寒沙門氏菌無標記適配體傳感器。羅丹明(rhodamine,RB)與適配體和AuNPs共存時,RB熒光由于熒光共振能量轉移(fluorescence resonance energy transfer, FRET)而發生猝滅。鼠傷寒沙門氏菌加入體系后與適配體特異性結合,RB熒光強度增強。樣品中鼠傷寒沙門氏菌濃度與熒光強度有良好的線性關系,檢出限為464 CFU/mL。ZHANG等[37]開發了一種基于EuPSNs的熒光免疫層析試紙條。將EUPSN-單克隆抗體與鼠傷寒沙門氏菌進行孵育,孵育過程中標記病原菌。然后將標記的病原體放置在試紙條(預噴涂抗體)上,形成三明治結構(抗體-病原體-抗體),并使用發光閱讀器對IST條帶進行分析。該試紙條對鼠傷寒沙門氏菌的檢測限為103CFU/mL,檢測時間小于1.5 h,是一種快速、高靈敏度的檢測方法。FANG S Y等[38]構建了一種新型倏逝波雙色熒光適配體傳感器。基于雙色倏逝波熒光原理和微/納米尺寸效應的機理可用于同時檢測大腸桿菌O157:H7和鼠傷寒沙門氏菌。將520 nm和635 nm的激發光交替引入光纖納米探針,交替激發兩種熒光標記的適配體Cy3-apt-E.coli和Cy5.5-apt-S.typhimurium,并由一個基于時間分辨效應的光電探測器同時進行檢測。由于倏逝波的穿透深度有限,且細菌和納米孔的尺寸差異顯著,纖維納米探針能夠有效區分游離核酸傳感器和與病原菌結合的核酸傳感器。隨著病原菌濃度的增加,更多的核酸適配體與各自的靶菌特異結合,從而導致較低的熒光強度。根據細菌數量與熒光強度之間的線性關系,可以對兩種致病菌進行定量分析。該檢測方法對大腸桿菌O157:H7和鼠傷寒沙門氏菌的檢測限分別340 CFU/mL和180 CFU/mL。倏逝波雙色熒光適配體傳感器可以實時檢測兩種病原菌,而無需繁瑣的培養、濃縮和分離步驟,較其他生物傳感器更為方便、快捷。
2.4.3納米技術
隨著納米技術的迅速發展,各種納米材料由于其獨特的磁性、光學和其他相關性質,為不同的目標分析物提供了高效的化學/生物傳感平臺[39-42]。納米材料的表面體積比較大,有利于分析物通過共價或非共價的方式附著在各種識別部分,從而控制和改變其性質[43-44]。由于尺寸和表面結構的良好可控性,納米材料的分析性能(如成本、反應動力學、準確度和重復性等)得到顯著提高,更容易實現僅需少量樣本量的快速現場檢測[45-47]。與傳統的病原體檢測方法相比,基于納米材料的檢測方法具有成本低、快速、準確度高等優點,特別是在高通量篩選、無標簽和實時響應方面,能夠充分滿足食品行業、醫院和監管機構的需要。ZHENG Z等[48]構建了一種基于磁性納米材料的磁性和光熱效應的快速檢測鼠傷寒沙門氏菌的方法(圖3[48])。鼠傷寒沙門氏菌和磁性納米材料與抗鼠傷寒沙門氏菌抗體偶聯物進行孵育,通過膜過濾去除游離磁性納米材料和磁分離步驟去除非特異性細菌,然后用激光筆照射鼠傷寒沙門氏菌保留膜上的磁性納米材料,使其產生溫度變化。根據溫度變化反映鼠傷寒沙門菌的濃度,其檢測限為300 CFU/mL。CHO I H等[49]基于磁性納米粒子Fe3O4設計了一種原位熒光免疫磁傳感器,使其能夠同時檢測鼠傷寒沙門氏菌在內的三種病原體。目標細菌被IMBs和抗體結合的有機熒光團捕獲并鑒定,形成三明治復合結構。隨后,在蛋白酶K的作用釋放熒光物質,通過熒光顏色和強度反映病原菌的種類和濃度。與傳統的免疫檢測方法相比,該磁珠的表面積大,有利于對目標細菌的捕獲,是一種快速、靈敏的檢測方法。

圖3 (A)免疫磁性納米材料識別鼠傷寒沙門氏菌示意圖;(B)基于免疫磁納米材料光熱效應的鼠傷寒沙門氏菌捕獲、熱傳感器檢測和滅活程序[48]
傳統檢測技術準確性高,但仍有較多缺陷。以膠體金免疫層析試紙條為例,膠體金產品存在批次差異大,靈敏度低,標記物不穩定等問題,而且該技術的特異性高度依賴捕獲抗體的質量,而捕獲抗體獲取過程本就極為繁瑣[50]。此外,傳統檢測技術檢測周期長,對操作人員以及環境要求較高。因此,隨著單一檢測技術弊端的出現,新型檢測技術逐漸進入人們的視野。該方法在一定程度上可以彌補單一檢測技術產生的缺陷。將傳統檢測技術與其他分析檢測方法相結合,極大地豐富了鼠傷寒沙門氏菌檢測技術的內容,例如將發光納米材料與免疫層析試紙條相結合,通過檢測熒光強度,實現對病原菌的定量檢測,解決了傳統膠體金免疫層析試紙條只能定性或半定量的問題。新型檢測技術的誕生,促進了鼠傷寒沙門氏菌檢測技術的迅速發展,但在實際應用中仍然存在較多缺陷。表1總結了現有檢測技術的優缺點。

表1 傳統檢測技術和新型檢測技術優缺點
3 總結與展望
鼠傷寒沙門氏菌作為一種胞內病原菌,在逃避免疫系統清除以及在宿主細胞內繁殖方面有著非常靈活的表現。目前,針對鼠傷寒沙門氏菌的防治,主要采用抗生素治療和疫苗預防。常用的鼠傷寒沙門氏菌抗生素藥物是氟喹諾酮類或廣譜頭孢菌素[51]。雖然抗生素的殺菌能力強,特異性高,但是由于近年來抗生素的濫用,出現了沙門氏菌的耐藥性增強,耐藥譜變寬等問題[52]。與此同時,大劑量使用抗生素也會破壞宿主腸道菌群的平衡。這表明采用傳統抗生素治療方法的效果大不如前,而且會直接影響肉制品的衛生與安全,最終危及到人類自身健康。因此,深入了解鼠傷寒沙門氏菌的感染機制,開發新型綠色藥物來防治鼠傷寒沙門氏菌的感染對公共安全與經濟發展有著重大意義。
隨著鼠傷寒沙門氏菌在畜禽類及其制品中的感染逐漸增多,人們對鼠傷寒沙門氏菌的關注度隨之提高,同時也促進鼠傷寒沙門氏菌檢測方法的發展。以準確度為標桿,傳統方法處于檢測技術中的領先地位,但由于其檢測周期長這一弊端,在一些即時檢測現場并不能發揮作用。生物傳感器技術、光譜技術、納米技術和噬菌體庫檢測技術等新型檢測技術的出現,為鼠傷寒沙門氏菌的即時檢測提供了有力的技術支撐,但仍需不斷地完善改進。因此,開發方便、快捷的檢測技術是食源性致病菌檢測技術的發展方向,也是當今社會的迫切需要。相信在不久的將來,會出現更多快速、便捷、特異性強、高靈敏度、高通量和低成本的鼠傷寒沙門氏菌檢測技術,為人們的食品安全保駕護航。
猜你喜歡
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
兒童故事畫報(2019年5期)2019-05-26 14:26:14
海峽科技與產業(2016年3期)2016-05-17 04:32:12
Coco薇(2016年2期)2016-03-22 02:42:52
Coco薇(2015年1期)2015-08-13 02:47:34
小雪花·成長指南(2015年7期)2015-08-11 15:03:12
