氮磷水平對(duì)細(xì)基江蘺氮、磷吸收及生長(zhǎng)的影響*
2022-08-12 04:59:16付家想黃凌風(fēng)藍(lán)文陸黎明民李天深
廣西科學(xué) 2022年3期
付家想,潘 科,黃凌風(fēng),藍(lán)文陸,黎明民,李天深
(1.廣西科學(xué)院廣西紅樹林研究中心,廣西紅樹林保護(hù)與利用重點(diǎn)實(shí)驗(yàn)室,廣西北海 536007;2.廣西壯族自治區(qū)海洋環(huán)境監(jiān)測(cè)中心站,廣西北海 536000;3.深圳大學(xué)高等研究院,廣東深圳 518000;4.廈門大學(xué)環(huán)境與生態(tài)學(xué)院,福建廈門 361102)
近年來,人類生產(chǎn)、生活,尤其是近海養(yǎng)殖活動(dòng)過量投餌給水環(huán)境帶來空前壓力,水體富營(yíng)養(yǎng)化、生態(tài)環(huán)境惡化已成為一大環(huán)境難題[1],其主要原因是氮(N)、磷(P)含量的急劇增加。利用生物濾器大型藻類修復(fù)水質(zhì)成為解決問題的關(guān)鍵,大型海藻不僅能從周圍環(huán)境中大量吸收N、P營(yíng)養(yǎng)元素[1-3]及重金屬元素,還能提高水體溶解氧水平,調(diào)節(jié)水體pH值,從而改善水生生境。N、P是初級(jí)生產(chǎn)者生長(zhǎng)的營(yíng)養(yǎng)基礎(chǔ)[2],在營(yíng)養(yǎng)鹽貧乏水域,加富N、P會(huì)顯著促進(jìn)藻體對(duì)其吸收,然而N、P對(duì)藻類生長(zhǎng)的影響較復(fù)雜,與藻類生長(zhǎng)的環(huán)境因子、氮源和磷源種類、生長(zhǎng)速率、碳代謝、底物酶活性和N/P等因素有關(guān)[3-5]。對(duì)限制性營(yíng)養(yǎng)要素的吸收能力是藻類大量繁殖的關(guān)鍵,適宜的N/P有利于浮游植物的生長(zhǎng)繁殖,反之會(huì)造成營(yíng)養(yǎng)結(jié)構(gòu)失調(diào)、種群結(jié)構(gòu)改變,甚至引發(fā)藻華災(zāi)害。大量研究證實(shí),黃海綠潮災(zāi)害頻發(fā)與海水中營(yíng)養(yǎng)鹽過量密切相關(guān)[6,7]。自20世紀(jì)60年代國(guó)外學(xué)者首次提出Redfield比[8](即維系海洋藻類正常生長(zhǎng)所需的適宜N/P為16/1)后,關(guān)于N、P限制對(duì)大型藻類生長(zhǎng)影響的研究[9-16]已有不少。研究發(fā)現(xiàn),N、P差異顯著會(huì)影響大型海藻江蘺屬(Gracilaria)的生長(zhǎng)狀況和生理特征,例如,錢魯閩等[13]明確了不同形態(tài)的N、N/P會(huì)影響菊花江蘺(Gracilarialichenoides)對(duì)P的吸收,當(dāng)N/P為10/1時(shí)吸收速率最佳;楊芳[14]發(fā)現(xiàn)位于東興、北海和深圳灣3個(gè)海區(qū)的細(xì)基江蘺(G.tenuistipitata)繁枝變型對(duì)不同形態(tài)N的富集能力存在差異,當(dāng)N濃度為20 μmol·L-1時(shí),藻體硝酸還原酶(Nitrate Reductase,NR)、超氧化物歧化酶(Superoxide Dismutase,SOD)活性最高;許忠能等[15]發(fā)現(xiàn)細(xì)基江蘺繁枝變種吸收的N/P隨環(huán)境N/P的不同而變化,當(dāng)N濃度一定時(shí)環(huán)境N/P對(duì)細(xì)基江蘺繁枝變種吸收N的速率無顯著影響;李恒等[16]研究發(fā)現(xiàn),當(dāng)N濃度為50 μmol·L-1時(shí),真江蘺(G.asiatica)和脆江蘺(G.chouae)對(duì)氮的平均吸收速率最大,且脆江蘺對(duì)硝酸鹽和磷酸鹽有更好的去除效果。
目前N、P對(duì)大型海藻江蘺屬的脅迫研究?jī)H見菊花江蘺、細(xì)基江蘺繁枝變型、脆江蘺[15]、真江蘺[16]等,對(duì)細(xì)基江蘺的研究主要集中在繁育、生物學(xué)特性以及與其他藻類相生相克效應(yīng)[17]方面,有關(guān)細(xì)基江蘺在不同N、P水平下的植物營(yíng)養(yǎng)生理生態(tài)研究尚缺乏。本研究以Redfield比[18]為理論依據(jù),研究在不同N、P營(yíng)養(yǎng)條件下細(xì)基江蘺對(duì)營(yíng)養(yǎng)鹽的吸收特性及對(duì)自身生長(zhǎng)的影響,以期為利用大型海藻吸收N、P,降低水體富營(yíng)養(yǎng)水平,改善近岸海域水環(huán)境質(zhì)量提供科學(xué)依據(jù)。
1 材料與方法
1.1 藻種
細(xì)基江蘺(G.tenuistipitata)的采集與保存:實(shí)驗(yàn)所用細(xì)基江蘺采自廣西北海市金海灣潮間帶。選擇生長(zhǎng)狀態(tài)良好、長(zhǎng)勢(shì)相近的藻體,并用海水沖洗除去雜藻、污泥及其他附著生物,室內(nèi)暫養(yǎng)。
暫養(yǎng)條件:每1 L海水添加10 mL f/2培養(yǎng)液,海水為經(jīng)高溫(121℃,20 min)消毒且過0.45 μm醋酸纖維濾膜的自然海水(pH值為8.1,鹽度為28.3),暫養(yǎng)溫度(20±1)℃,光照強(qiáng)度3 000-4 000 lx,光照周期14L∶10D。暫養(yǎng)期間每3 d更換一次海水且24 h充氣,以保證營(yíng)養(yǎng)鹽混合均勻并使藻體保持懸浮狀態(tài),暫養(yǎng)時(shí)間為1周。
實(shí)驗(yàn)用細(xì)基江蘺的準(zhǔn)備:取長(zhǎng)勢(shì)良好的細(xì)基江蘺接種至未加N源、P源的f/2營(yíng)養(yǎng)液中饑餓培養(yǎng)48 h,其他培養(yǎng)條件同上。
1.2 N、P濃度設(shè)置
不同N/P實(shí)驗(yàn):設(shè)置無機(jī)氮(NaNO3∶NH4Cl=1∶1,C∶C)濃度分別為0 μmol·L-1(N0)、40 μmol·L-1(N40)、160 μmol·L-1(N160)、320 μmol·L-1(N320)4個(gè)處理組,各處理組無機(jī)磷(NaH2PO4)濃度均為10 μmol·L-1,每個(gè)處理組各設(shè)置3個(gè)重復(fù),共12組。其余營(yíng)養(yǎng)元素按照f/2培養(yǎng)基的比例添加。
不同N、P濃度實(shí)驗(yàn):固定Redfield比N/P=16/1,設(shè)置5組不同N、P濃度,各處理組設(shè)置3個(gè)平行,共15組。N源、P源和其他營(yíng)養(yǎng)鹽元素與不同N/P實(shí)驗(yàn)設(shè)置相同。具體的N、P濃度設(shè)置見表1。

表1 不同N、P濃度條件下N、P濃度設(shè)置Table 1 Setting of N and P concentrations under different N and P concentrations
1.3 實(shí)驗(yàn)管理

1.4 測(cè)定方法

細(xì)基江蘺對(duì)營(yíng)養(yǎng)鹽的吸收速率v計(jì)算公式[14]如下:
(1)
式中:v為營(yíng)養(yǎng)鹽的吸收速率(μmolg-1h-1),Ct-1、Vt-1為每次取樣時(shí)間間隔的起始培養(yǎng)液中營(yíng)養(yǎng)鹽的濃度和體積,Ct、Vt為每次取樣時(shí)間間隔結(jié)束時(shí)培養(yǎng)液中營(yíng)養(yǎng)鹽的濃度和體積,t為培養(yǎng)時(shí)間(h),W為藻體濕重(g)。
細(xì)基江蘺對(duì)營(yíng)養(yǎng)鹽的去除率r計(jì)算公式如下:
式中:r為營(yíng)養(yǎng)鹽的去除率,C0、V0為實(shí)驗(yàn)開始前培養(yǎng)液中營(yíng)養(yǎng)鹽的濃度和體積,Ct、Vt為每次取樣時(shí)培養(yǎng)液中營(yíng)養(yǎng)鹽的濃度和體積。
稱量前先用紙巾吸去藻體表面的水分,再用精確度為0.001 g的電子分析天平稱量藻體濕重。藻體相對(duì)生長(zhǎng)速率 (Relative Growth Rate,RGR,以濕重計(jì),%d-1)計(jì)算公式[15]如下:
(2)
式中:W0是藻體初始濕重,Wt是t時(shí)刻藻體濕重。
靜態(tài)培養(yǎng)下的大型海藻對(duì)營(yíng)養(yǎng)鹽的吸收過程可以用一級(jí)動(dòng)力學(xué)方程來描述[12],公式[19]如下:
(3)
式中:S0為初始N、P濃度(mg·L-1),Se為t時(shí)刻N(yùn)、P濃度(mg·L-1),t為時(shí)間(d),k為N、P去除率常數(shù)。
1.5 數(shù)據(jù)處理
實(shí)驗(yàn)數(shù)據(jù)為3個(gè)平行樣的平均值,用Origin 9.0軟件處理擬合,用SPSS 19.0進(jìn)行數(shù)據(jù)統(tǒng)計(jì)分析、差異性檢驗(yàn),用one-way ANOVA及Duncan多重比較檢驗(yàn)組間差異(以P<0.05為差異顯著性標(biāo)準(zhǔn))。
2 結(jié)果與分析
2.1 不同N、P組合處理對(duì)細(xì)基江蘺生長(zhǎng)的影響
圖1和圖2展示了不同N、P組合條件下細(xì)基江蘺的生長(zhǎng)情況,不同N、P組合的細(xì)基江蘺濕重及其相對(duì)生長(zhǎng)速率具有明顯差異。細(xì)基江蘺在各處理組的生長(zhǎng)趨勢(shì)基本一致,藻體濕重在整個(gè)實(shí)驗(yàn)階段隨著實(shí)驗(yàn)時(shí)間延長(zhǎng)而增加(圖1)。由圖1(a)可知,在不同N/P下N160組(N初始濃度為160 μmol·L-1)藻體濕重增量比最大,達(dá)48.3%,顯著高于其他3個(gè)實(shí)驗(yàn)組(P<0.01);N0組(N初始濃度為0)藻體濕重增幅最小,僅為12.7%。由圖2(a)可知,不同N/P條件下各處理組間的藻體平均相對(duì)生長(zhǎng)速率大小依次為N160>N320>N40>N0,N40組與N320組無顯著差異(P>0.05),其他組各組間差異顯著(P<0.05)。

圖1 不同N、P水平下細(xì)基江籬藻體濕重隨時(shí)間的變化Fig.1 Variation of wet weight of G.tenuistipitata with time under different N and P levels
由圖1(b)可知,在不同N、P濃度條件下,P10組藻體增重比最高,達(dá)50.8%;P6組藻體增重比最低,為25.7%;P4組、P6組、P25組間差異不顯著,P16組在培養(yǎng)的整個(gè)階段,藻體濕重增長(zhǎng)率持續(xù)增加。由圖2(b)可知,不同N、P濃度條件下各組藻體相對(duì)生長(zhǎng)速率表現(xiàn)為P10>P25>P16>P6>P4,P6組與P16組、P16組與P25組無顯著性差異(P>0.05) ,其他組各組間差異顯著(P<0.05)。

Different letters indicate significant differences between groups (P<0.05)
2.2 細(xì)基江蘺對(duì)的去除
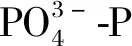

圖3 不同N、P水平下濃度、去除率隨時(shí)間的變化Fig.3 Variation of concentration and removal rate with time under different levels of N and P

表2 在不同初始P濃度下細(xì)基江蘺去除P的一級(jí)動(dòng)力學(xué)擬合結(jié)果Table 2 The first-order kinetic fitting results of P removal by G.tenuistipitata under different initial P concentrations

圖4 不同初始水平下細(xì)基江蘺N、P吸收速率Fig.4 Uptake rates of G.tenuistipitata under different initial levels
2.3 細(xì)基江蘺對(duì)的去除


圖5 不同N、P水平下濃度、去除率隨時(shí)間的變化Fig.5 Variation of concentration and removal rate with time under different N and P levels

表4 在不同初始濃度下細(xì)基江蘺去除N的一級(jí)動(dòng)力學(xué)擬合結(jié)果
2.4 細(xì)基江蘺對(duì)的去除


圖6 不同N、P水平下濃度、去除率隨時(shí)間的變化Fig.6 Variation of concentration and removal rate with time under different initial N and P levels

3 討論
3.1 N、P環(huán)境對(duì)細(xì)基江蘺生長(zhǎng)的影響
本研究結(jié)果表明,在不同的營(yíng)養(yǎng)鹽水平下細(xì)基江蘺都能夠較好地生長(zhǎng)和吸收N、P。在經(jīng)過2 d饑餓處理后長(zhǎng)達(dá)8 d的實(shí)驗(yàn)過程中,即使在氮缺乏的N0組,細(xì)基江蘺仍保持生長(zhǎng),生長(zhǎng)率達(dá)到2%d-1。這可能是細(xì)基江蘺在獲得性外源P的刺激下啟動(dòng)液泡等營(yíng)養(yǎng)儲(chǔ)備庫(kù)中的N源補(bǔ)充藻體生長(zhǎng)對(duì)N的需求。如鄒定輝等[20]研究發(fā)現(xiàn)藻細(xì)胞可以大量吸收并將其存儲(chǔ)在液泡或胞質(zhì)內(nèi),使細(xì)胞內(nèi)氮營(yíng)養(yǎng)鹽水平濃縮105倍以上。李楓[11]研究發(fā)現(xiàn)低氮高磷可促進(jìn)龍須菜(Asparagusschoberioides)的生長(zhǎng),并且與低氮低磷相比,低氮高磷處理提高了龍須菜的光化學(xué)效率,這表明海藻有適應(yīng)氮限制的內(nèi)在機(jī)制。本實(shí)驗(yàn)結(jié)果表明細(xì)基江蘺存在營(yíng)養(yǎng)鹽儲(chǔ)存機(jī)制,并具有較強(qiáng)的儲(chǔ)備營(yíng)養(yǎng)素能力,因此能夠在外源性N、P缺乏時(shí)利用儲(chǔ)備庫(kù)中的營(yíng)養(yǎng)鹽存活并保持一定的生長(zhǎng)率。
本研究結(jié)果還顯示,在不同N/P下,N160組(N/P為16/1)細(xì)基江蘺生長(zhǎng)率最高,藻體濕重增幅達(dá)48.3%,顯著高于其他3個(gè)實(shí)驗(yàn)組(P<0.01),N/P也與Redfield比值(16/1)[18]一致。對(duì)多種藻類的研究[21,22]表明,不同N/P對(duì)藻類的生長(zhǎng)影響顯著,N/P差異會(huì)對(duì)藻細(xì)胞的生長(zhǎng)、胞內(nèi)物質(zhì)的合成與積累產(chǎn)生較大影響。對(duì)藻類的營(yíng)養(yǎng)生理研究發(fā)現(xiàn),適應(yīng)于富營(yíng)養(yǎng)化水質(zhì)的硅藻等在Redfield比值附近生長(zhǎng)狀態(tài)最佳,在介質(zhì)營(yíng)養(yǎng)鹽充足情況下,中肋骨條藻(Skeletonemacostatum)和威氏海鏈藻(Thalassiosiraweissflogii)所需的N/P組成接近16/1,而藍(lán)藻、甲藻等適于寡營(yíng)養(yǎng)環(huán)境的微藻更能適應(yīng)N/P偏離Redfield比值的環(huán)境[23],米氏凱倫藻(Kareniamikimotoi)在N/P為80/1的條件下比生長(zhǎng)速率最大[24],可見N/P對(duì)不同種類海藻的影響還與他們對(duì)營(yíng)養(yǎng)環(huán)境長(zhǎng)期適應(yīng)密切相關(guān)。細(xì)基江蘺主要生長(zhǎng)在富含N、P沖淡水匯入的河口等近岸海區(qū),本研究所用的細(xì)基江蘺也是采集于富營(yíng)養(yǎng)化的潮間帶海區(qū),能適應(yīng)富營(yíng)養(yǎng)的環(huán)境,研究結(jié)果表明N/P對(duì)細(xì)基江蘺生長(zhǎng)有顯著影響,適應(yīng)富營(yíng)養(yǎng)環(huán)境的細(xì)基江蘺傾向于按Redfield比將所吸收N、P用于自身組織生長(zhǎng),偏離Redfield比則對(duì)細(xì)基江蘺的生長(zhǎng)有明顯的抑制效應(yīng)。
3.2 N、P環(huán)境對(duì)細(xì)基江蘺吸收N、P的影響

3.3 細(xì)基江蘺去除N、P的能力
N、P營(yíng)養(yǎng)鹽是海水集約化養(yǎng)殖系統(tǒng)中造成水體富營(yíng)養(yǎng)化、危害養(yǎng)殖對(duì)象的主要因子,目前利用大型海藻去除養(yǎng)殖廢水中N、P已成為國(guó)內(nèi)外的研究熱點(diǎn),學(xué)者們一致認(rèn)為利用大型海藻進(jìn)行水質(zhì)修復(fù)是凈化水體、延緩水體富營(yíng)養(yǎng)化的重要措施之一[29-31]。在江蘺屬的相關(guān)報(bào)道中,在25℃條件下菊花江蘺特定生長(zhǎng)速率為2.3%d-1-3.02%d-1[32],25℃時(shí)脆江蘺相對(duì)生長(zhǎng)速率約為2.6%d-1[15];但金玉林等[33]發(fā)現(xiàn)26℃時(shí)脆江蘺相對(duì)生長(zhǎng)速率在5%以上。分析本研究中細(xì)基江蘺的生長(zhǎng)數(shù)據(jù)得知,細(xì)基江蘺吸收的營(yíng)養(yǎng)鹽主要用于生長(zhǎng),特定生長(zhǎng)率為(5.74±0.42)%d-1,高于同類報(bào)道中的數(shù)值。本研究結(jié)果顯示,在最佳N/P下細(xì)基江蘺在培養(yǎng)后期(第8天)對(duì)培養(yǎng)液中的去除率均可達(dá)96.8%,對(duì)的去除率為83.9%,對(duì)的去除率為94.9%,顯著高于許忠能等[15]研究中細(xì)基江蘺繁枝變種對(duì)和的去除率,也高于吳翔宇等[34]關(guān)于芋根江蘺在28 d時(shí)對(duì)和的去除率。由此可見,細(xì)基江蘺可用于凈化水質(zhì),具有作為生物濾器的潛在可能性。
猜你喜歡
體育科技文獻(xiàn)通報(bào)(2022年3期)2022-05-23 13:46:54
天津外國(guó)語(yǔ)大學(xué)學(xué)報(bào)(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
小讀者(2021年2期)2021-03-29 05:03:48
科技傳播(2019年22期)2020-01-14 03:06:54
瘋狂英語(yǔ)·新悅讀(2019年11期)2019-12-18 05:14:16
華人時(shí)刊(2019年13期)2019-11-17 14:59:54
民用飛機(jī)設(shè)計(jì)與研究(2019年4期)2019-05-21 07:21:24
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
