烘焙預處理對小球藻熱解特性與產物分布的影響
2022-08-17 06:38:14謝東明
可再生能源 2022年8期
謝東明, 鐘 典, 李 俊, 曾 闊, 彭 靜
(1.廣西壯族自治區特種設備檢驗研究院, 廣西 南寧 530200; 2.華中科技大學 能源與動力工程學院, 煤燃燒國家重點實驗室, 湖北 武漢 430074; 3.華中科技大學 中歐清潔與可再生能源學院, 湖北 武漢 430074)
0 引言
藻類作為第三代生物質,因其具有生長速度快、光合效率高、不占用耕地、生物油產率高、固碳能力強等優點,近些年受到了人們的重視[1]~[4]。微藻中含氮量為5.2%~16.3%[2],[4],[5], 其熱解生物油中包括酰胺類、胺類、吡咯類、喹啉類、吡啶類、吲哚類等含氮物質。 早先的研究集中在將含氮物質從中裂解出去,生成烴類、氨等物質作為運輸燃料。 近年來,這些含氮物質可作為一些精細化學品的重要前驅體而受到了研究者的廣泛關注[4],[5]。如何通過更優的處理方式實現藻類生物質熱解高值化利用成為了新的挑戰。
烘焙(200~300 ℃)是生物質常見的預處理方式,可有效地提高生物質原料的能量密度,改善其機械性能, 并能在一定程度上解決生物質,特別是藻類生物質含水率高、熱值低、不易儲存等問題,降低運輸、處理、儲存以及作為能源利用的成本[6]。 烘焙預處理對生物質熱解也具有深遠的影響。 研究表明,隨烘焙溫度的升高,熱解生物油的品質有所提高,水分含量最多減少了42.8%,稠環化合物及酚類產物的含量有所增加,熱值約升高34%,但是生物油的產率大為降低,焦炭的含量增加,其化學組成逐漸向煤過渡[7]。 此外,烘焙時間越長,烘焙溫度越高,熱解時的交聯反應和碳化縮合反應就越多, 使熱解過程中水分和CO2析出減少, 且水的析出溫度由120 ℃提升至200℃[8]。這表明,需要對整個生產過程進行綜合考慮,確保生物油含氧含水量高的缺點被合理補償。 文獻[9]指出,在較高烘焙溫度下,木質素的側鏈斷裂,導致具有側鏈的酚類產量減少,不含有側鏈的酚類產量增加。 宇妍[10]探究了不同脫水方式與烘焙聯合預處理對生物質燃料特性的影響, 經過聯合預處理進一步提升了生物質的能源化利用品質。 藻類生物質與木質纖維素類生物質成分差異較大, 烘焙預處理對其熱解特性與產物分布的影響是否類似, 以及對含氮產物生成的影響仍有待研究。
本文在不同烘焙預處理條件下對小球藻熱解特性的影響規律進行了研究。 通過在不同升溫速率下的熱重實驗,探究其在40~900 ℃下的熱重反應特性, 采用分布式活化能模型(DAEM)計算烘焙后小球藻的熱解活化能,揭示烘焙預處理對其熱解動力學的影響;在升溫速率為10 ℃/min 的條件下采用熱重-紅外聯用儀分析烘焙后小球藻熱解氣體中含氮化合物的變化規律;采用Py-GC/MS 在不同熱解溫度(500 ℃和900 ℃) 下探究烘焙預處理對其熱解生物油的影響規律。
1 實驗原料與方法
1.1 實驗原料
本文所用的小球藻(干燥基)使用前先在105℃烘箱中干燥24 h,再手工磨碎,并用30 目篩網篩分后得到粒徑小于0.613 mm 的干燥原料,其工業分析 (GB/T 28731—2012)、 元素分析(Vario EL cube,德國元素分析系統公司)以及高位熱值(微電腦熱量計系統,WZR-1T-CII & IKAC2000)結果如表1 所示。

表1 小球藻原料性質Table 1 Proximate and ultimate analysis of Chlorella
1.2 實驗儀器與設備
烘焙預處理實驗裝置如圖1 所示。
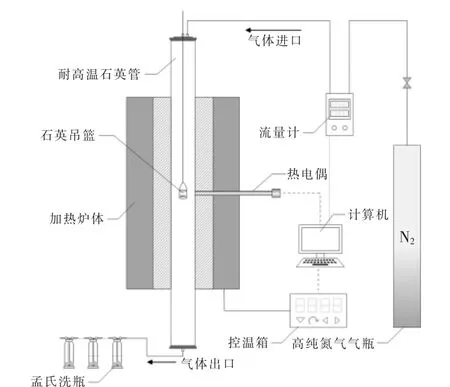
圖1 烘焙實驗裝置示意圖Fig.1 Schematic diagram of the experimental setup
稱取2~5 g 干燥好的小球藻粉末, 裝入石英吊籃中;將石英吊籃固定在耐高溫石英管頂部,通入氮氣 (純度99.999 9%,245 mL/min)30 min,對石英管進行排空; 將管式爐預熱至所需溫度(200,225,250,275,300,325 ℃, 溫度間隔25 ℃)并保持恒溫;待爐體溫度達到所需溫度后,降低載氣流量至170 mL/min,同時將裝入樣品的石英吊籃推入反應器中部, 分別保持相應的停留時間(30,60,90 min)。 在實驗過程中,采用三段式洗滌去除尾氣中的污染物和焦油, 之后排放至室外(250 mL 孟氏洗瓶, 前兩個裝入異丙醇和玻璃珠的混合物,第3 個裝入變色硅膠)。實驗結束后,取出吊籃進行稱重,獲得烘焙前后的質量變化。每次實驗至少重復3 次,且相對誤差不超過5%,取3次的平均值計算質量產率、 相對能量密度和能量產率。
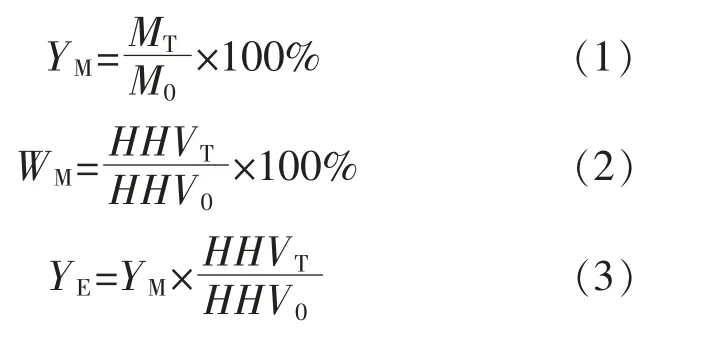
式中:YM為生物質烘焙后的質量產率,%;WM為生物質烘焙后的相對能量密度,%;YE為生物質烘焙后能量產率,%;MT為烘焙后收集的生物質質量,g;M0為烘焙前實驗稱取的生物質質量,g;HHV0,HHVT分別為烘焙前、 后生物質的高位熱值,kJ/kg。
對不同條件下烘焙預處理的產物與原樣進行熱重實驗。 升溫速率設置為5,10,20 ℃/min,在氬氣氛圍下從40 ℃升溫至900 ℃,同時采用同步熱分析-紅外光譜聯用儀(STA449F3Jupiter,德國耐馳)對揮發分進行紅外光譜分析。 采用裂解器-氣相色譜質譜聯用儀(CDS5200,美國CDS-Agilent)對相同幾組樣品進行熱解實驗, 熱解溫度設置為500 ℃和900 ℃,升溫速率為2 000 ℃/s,保持30 s。氣相色譜-質譜儀(GCMS)在線檢測樣品的揮發分組成,采用氬氣氣氛,色譜柱以30 ℃/min程序升溫至300 ℃,分流比為1∶50,質譜檢測的核質比為35~600 m/z。
2 結果與討論
2.1 烘焙對小球藻熱解動力學的影響
2.1.1 小球藻烘焙前后的熱解失重特性
選取烘焙時間為60 min,烘焙溫度為225,250,275 ℃時的烘焙產物及小球藻原樣4 個樣品進行熱重動力學測試, 得到生物質隨溫度的變化關系。 為了更好地對熱重數據進行分析,將DTG曲線中的最低點定義為峰值失重率, 所對應的溫度稱為峰值失重溫度。
圖2 為小球藻原樣及3 個烘焙樣品的TG 與DTG 曲線。 由圖2 可以看出,在不同升溫速率下,焦炭產率、峰值失重率都發生了變化,且開始失重的溫度隨升溫速率的升高而逐漸升高。 對比不同烘焙條件下的DTG 曲線可以看出:隨著烘焙溫度的上升, 樣品在所有升溫速率下的主失重峰(300~400 ℃)均出現滯后現象,對應的溫度升高(未烘焙樣品:278~312 ℃;225 ℃烘焙后:302~327℃;250 ℃烘焙后:325~352 ℃;275 ℃烘焙后:356~383 ℃);樣品在主失重峰的失重率也隨烘焙溫度的上升而減小, 而對應的400~500 ℃處肩峰的失重率則會上升。 研究指出,微藻三組分的熱重曲線會出現2~3 個峰, 在300~400 ℃的主峰主要是由于糖類和蛋白質的熱降解脫出揮發分所致,400~500 ℃的肩峰是由于脂類的熱降解所致[2],[3]。從DTG 曲線還可以看出, 隨著烘焙溫度的升高,主峰與肩峰的間距變小,由于脂類熱解區間較高,在烘焙過程中基本不發生變化, 而較易降解的糖類和蛋白質在該階段部分發生脫揮發分反應[11],烘焙使小球藻的均一性得到提高。
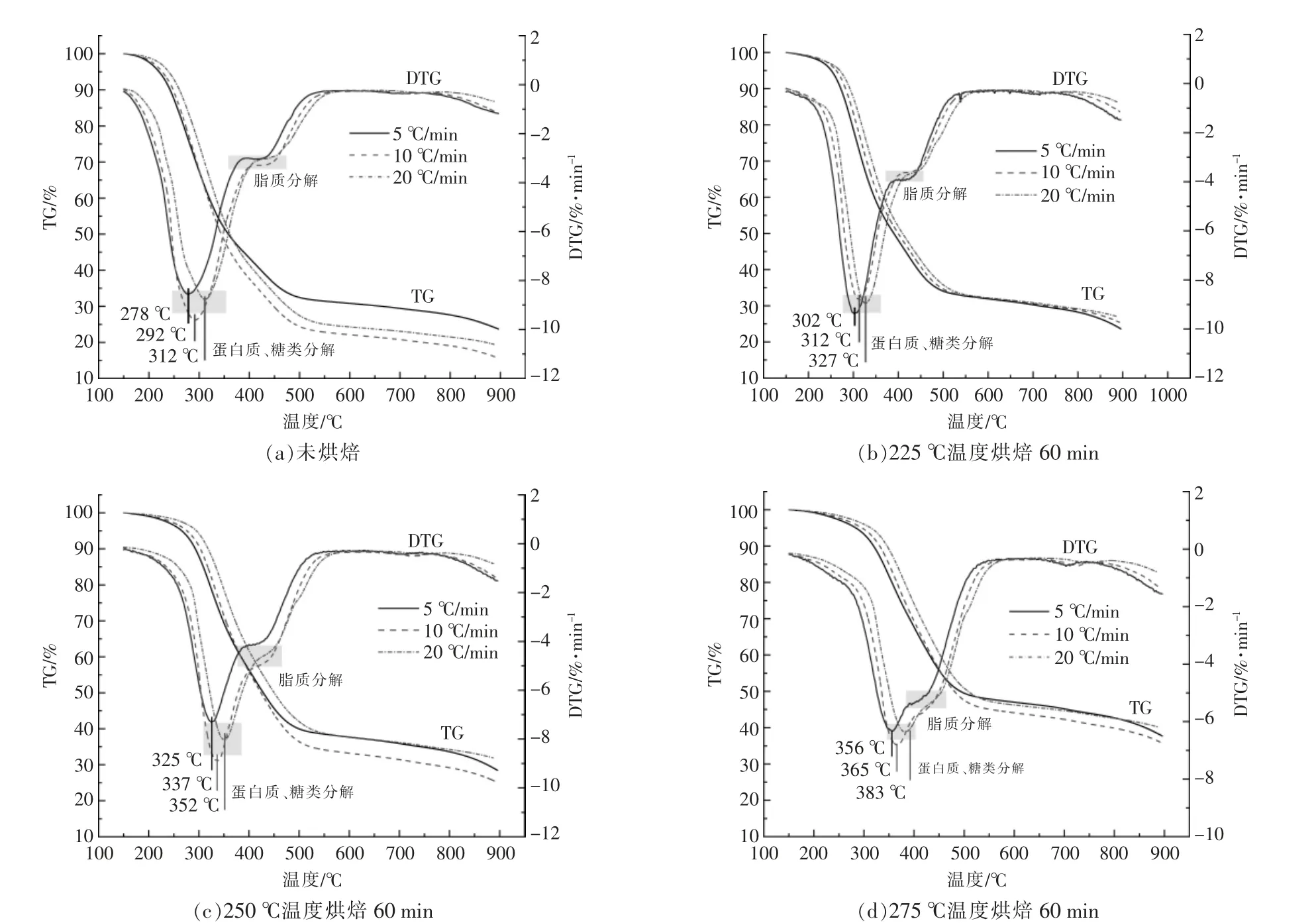
圖2 小球藻在不同升溫速率下TG 和DTG 曲線Fig.2 TG and DTG curves of Chlorella at different heating rates
不同熱解條件后的焦炭產率計算式分別為

式中:φrel為相對焦炭產率,%;φabs為絕對焦炭產率,%;Mresidual為焦炭質量,g。
不同樣品在不同升溫速率下的小球藻焦炭產率如圖3 所示。從圖可以看出,樣品最終焦炭產率受到烘焙條件和升溫速率變化的影響, 且烘焙條件的影響占主要地位。 以烘焙后的樣品質量計算相對焦炭產率, 烘焙處理明顯提高了相對焦炭產率,且隨著溫度升高而升高,以升溫速率為10℃/min 為例, 當烘焙溫度從225 ℃提高至275 ℃時,小球藻相對焦炭產率由15.76%增加至35.91%[圖3(a)]。 提高升溫速率會使原始樣品與經過烘焙處理后樣品的相對焦炭產率差異加大。
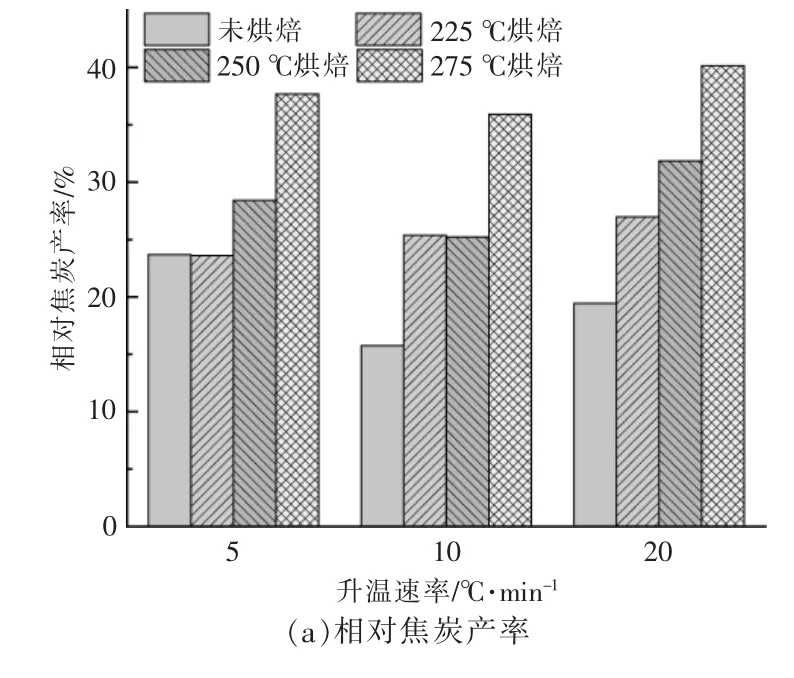
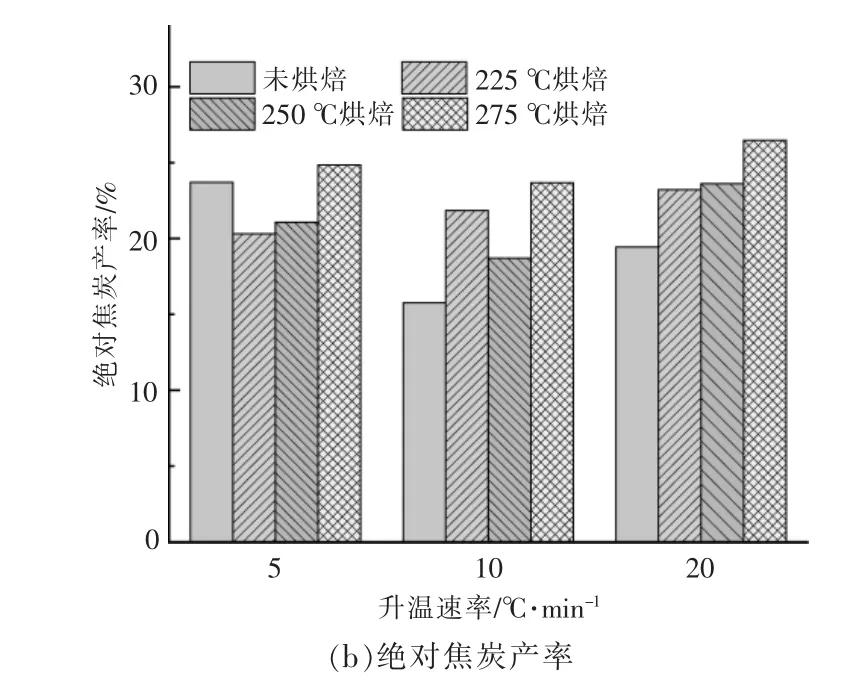
圖3 在不同升溫速率下小球藻烘焙前后的焦炭產率Fig.3 Residual rates of Chlorella before and after torrifying at different heating rates
若考慮烘焙過程中脫出的揮發分, 即以烘焙前原始樣品的質量計算絕對焦炭產率,如圖3(b)所示。此時隨著烘焙溫度的升高,絕對焦炭產率依舊呈現上升趨勢, 說明烘焙溫度升高會促進最后熱解焦的生成。 在升溫速率為5 ℃/min 時,225 ℃和250 ℃烘焙的小球藻的絕對焦炭產率甚至低于未烘焙原料,說明225~250 ℃烘焙處理后,小球藻在低升溫速率(5 ℃/min)下熱解更有利于揮發分的析出。
2.1.2 小球藻烘焙前后的熱解動力學
熱解活化能是動力學的重要參數, 生物質熱解是眾多官能團在熱能作用下一系列復雜連續的反應。由多條熱重曲線得出的分布式活化能,計算結果較準確且隨反應的加深而變化, 避免了單一表觀活化能生物質復雜反應的局限性[2],[12]~[14]。 本文采用活化能分布模型(DAEM)研究小球藻熱反應過程,探討熱解活化能與轉化率的關系[2],[12]~[14]。
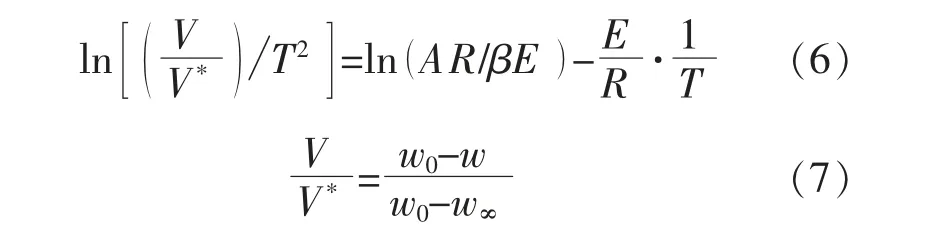
式中:V/V*為熱解轉化率,%;T 為溫度,K;A 為指前因子,s-1;R 為氣體常數,R=8.314 J/(mol·K);E為活化能,kJ/mol;w 為τ 時刻樣品質量,g;w0為樣品初始質量,g;w∞為樣品熱解后最終剩余量,g。
對熱重實驗中升溫速率為5,10,20 ℃/min 的熱重數據進行計算整理后,得到小球藻的Arrhenius譜圖及熱解活化能隨轉化率的變化關系, 分別如圖4,5 所示。 從圖4 可以看出,當轉化率為0.05~0.80 時, 擬合曲線的相關系數R2整體上均大于0.98 000,說明擬合效果較好,計算得到的活化能也較為可靠。從圖5 可以看出,烘焙預處理總體上會提升小球藻的熱解活化能, 但不同反應階段的影響不盡相同。 當轉化率低于0.2 時,僅225 ℃烘焙后的樣品初始階段活化能稍微大于原樣品,其余烘焙溫度樣品的活化能相比原樣差別不大;當轉化率為0.2~0.55 時, 不同烘焙溫度下小球藻的熱解活化能與原樣相比均明顯增大, 但烘焙溫度對活化能的增加影響不大。 此階段大致與圖2 中最大失重峰對應,即蛋白質與糖類的分解階段,說明烘焙處理使得糖類和蛋白質經歷了預熱解過程, 生成了一些更難以分解的產物; 當轉化率為0.55~0.75時,不同烘焙溫度下樣品的熱解活化能出現差異,這一階段中225 ℃烘焙的活化能最高,但隨烘焙溫度升高活化能反而逐漸降低, 與原始樣品接近。此階段大致與圖2 中肩峰對應,即脂質的分解階段, 說明雖然烘焙處理會使脂質變為更難分解的產物,但提高烘焙溫度反而會逐漸消除這一影響,使熱解反應更容易進行。 在最后階段,烘焙處理的樣品活化能均出現激增, 以275 ℃烘焙樣品為代表,此階段對應了焦炭的生成,說明烘焙處理后焦炭產物增加,難以再進行分解。這也對應了圖3 中的結論。
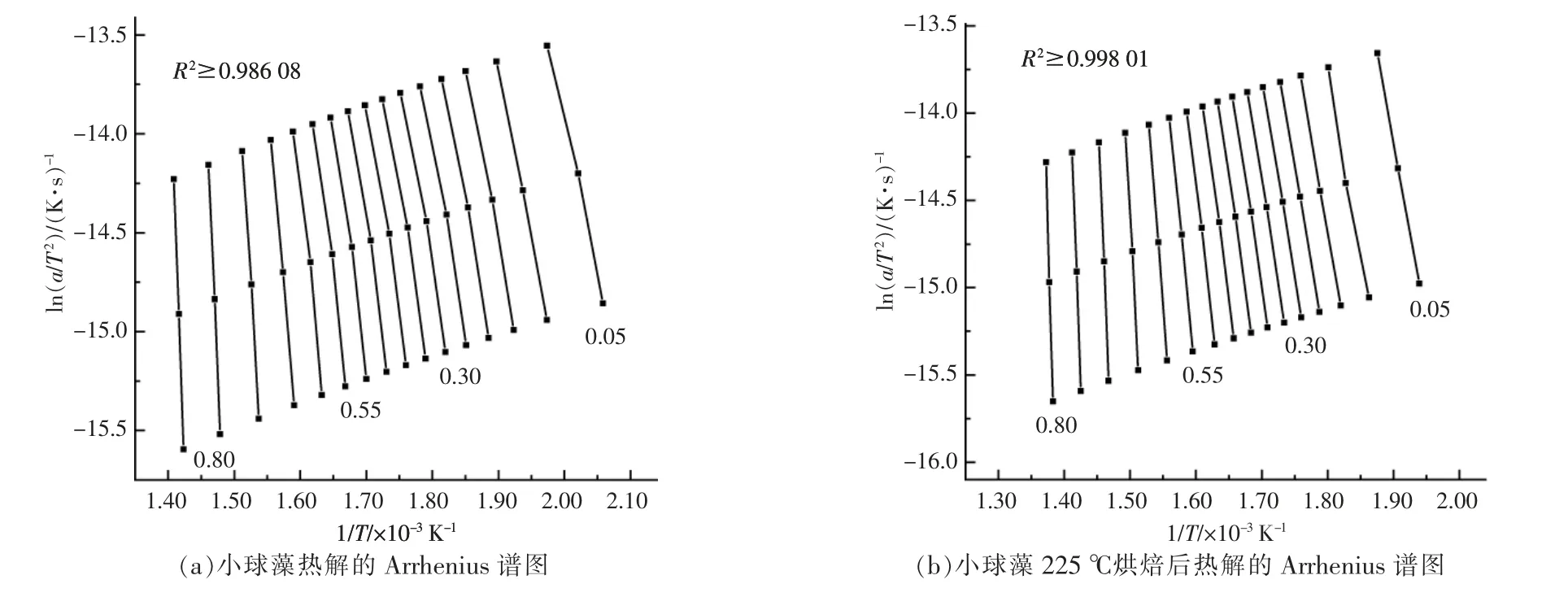
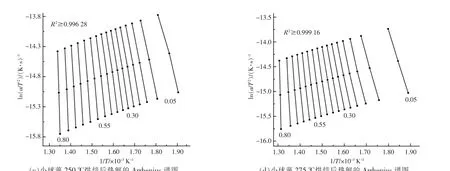
圖4 小球藻熱解動力學計算Fig.4 Calculation of pyrolysis kinetics of Chlorella vulgaris
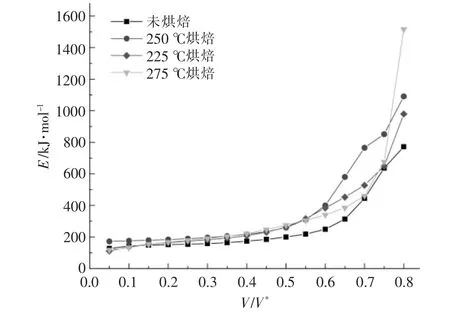
圖5 不同烘焙溫度下小球藻的熱解活化能隨轉化率的變化關系Fig.5 Changes of activation energy of Chlorella vulgaris at different torrification temperatures
2.2 烘焙對小球藻熱解氣體產物的影響
根據Lambert-Beer 定律[15],各揮發分氣體的紅外光譜的吸光度與氣體的濃度呈線性關系,特征吸收峰越高,說明該氣體的相對含量越高。 圖6為小球藻原樣及烘焙預處理后熱重-紅外聯用儀所得到的紅外三維立體圖。由圖可以看出,各個樣品熱解揮發分的紅外譜圖中均可見幾個顯著的吸收峰, 分別在3 200~3 400 cm-1,2 950 cm-1,2 340 cm-1,1 754 cm-1,1 500~1 600 cm-1,900~1 100 cm-1,669 cm-1。 其中大部分吸收峰隨反應時間變化,在熱解40 min 左右均出現最大值, 對應溫度為280~300 ℃, 結合各組樣品DTG 曲線可以發現,該溫度區間為最大失重峰出現的溫度區間。
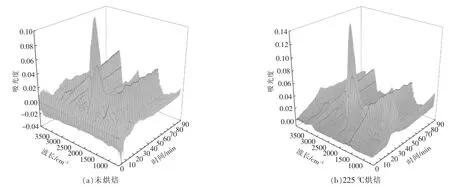
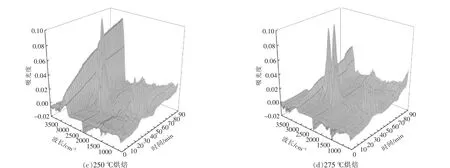
圖6 小球藻烘焙預處理后在10 ℃/min 下熱解時氣體的FTIR 分析Fig.6 FTIR analysis of pyrolytic volatile fraction of Chlorella vulgaris after pretreatment at 10 ℃/min
圖7 展示了上述4 種樣品在失重速率最大處的揮發分鑒定:①3 200~3 400 cm-1為羥基的吸收峰,對應揮發分中結合水的生成;②2 950 cm-1為C-H 鍵的伸縮振動峰,對應揮發分中的輕質烷烴類物質的生成;③2 340 cm-1與669 cm-1對應CO2吸收峰,標志熱解脫-CO 以及脫-COOH 反應;④1 754 cm-1對應有機組分中的C=O 結構, 主要存在于脂質熱解產物與有機酸產物中;⑤1 500~1 600 cm-1主要是芳烴中的C=C 鍵的吸收峰,對應產物中芳烴類物質的生成;⑥900~1 100 cm-1附近的峰由含氧官能團的吸收峰組成, 如有機組分中C-O 和無機組分中的P-O-P,Si-O,對應醇類、酚類以及無機硅磷酸鹽[16]。
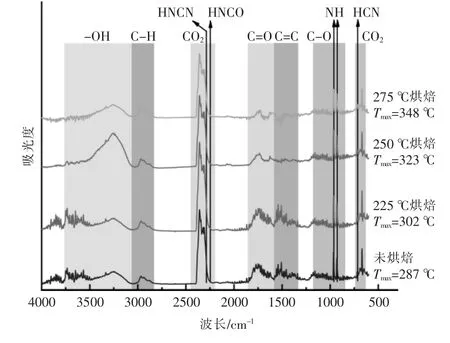
圖7 小球藻烘焙預處理前后在失重速率最大處的揮發分鑒定Fig.7 Identification of volatile fraction at the maximum rate of weight loss of Chlorella before and after torrification
對比烘焙處理前后以及不同烘焙溫度下的樣品熱解揮發分的紅外譜圖(圖7)可以發現,相比未烘焙的原樣,烘焙后樣品熱解產生的水變少,含氧官能團如羧酸、醛、酮等的析出減少,芳烴類物質減少,醇、醚以及烴類則沒有明顯變化。此外,隨著烘焙溫度的升高,CO2的含量經歷了一個先增加后減少的過程, 在烘焙溫度為225 ℃時達到峰值,但總的強度仍高于未烘焙原樣。
小球藻熱解產物中的4 種主要含氮官能團(NH,HCN,HNCO 和HNCN) 隨熱解溫度的變化如圖8 所示。總體來看,烘焙處理使小球藻熱解含氮物質增加, 但同時烘焙溫度的升高也會降低后續熱解含氮產物的生成。 圖8(a),(b)分別為不同烘焙溫度樣品的NH3和HCN 的生成強度, 可以看到,烘焙處理對其強度影響規律相似,生成的含氮物質強度由高到低依次為225 ℃烘焙>250 ℃烘焙>275 ℃烘焙>未烘焙。 這是由于烘焙溫度升高,部分含氮的產物在烘焙階段就提前釋放,導致熱解階段的含氮產物析出減少;圖8(c),(d)中,HCNO 和H2NCN 的含量由高到低依次為225 ℃烘焙>275 ℃烘焙>250 ℃烘焙>未烘焙, 其中烘焙時間均為60 min。 但這兩種相對復雜的含氮物質的析出強度并不和烘焙溫度呈線性相關, 而是在250 ℃出現極小值。 這可能是由于生成過程會受到脂質與糖類烘焙產物的影響, 進而影響了在后續熱解過程中交互脫氮的過程[17]。
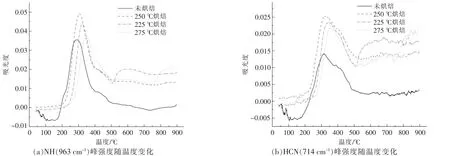
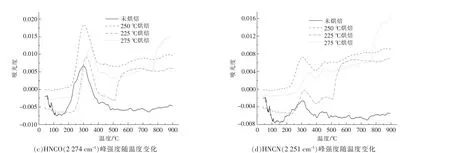
圖8 小球藻烘焙前后含氮物質隨溫度的變化關系Fig.8 Relationship between nitrogenous substances and temperature before and after torrification of Chlorella vulgaris
2.3 烘焙對小球藻熱解生物油組分的影響
為了便于分析, 根據原樣和烘焙樣品在不同熱解溫度下總離子流色譜圖峰面積的占比, 選取了數十種主要熱解產物。 將生成產物分為輕質揮發分、雜環化合物、脂肪族含氧化合物、其他含氧化合物及其他產物5 大類, 各類產物的占比如圖9 所示。
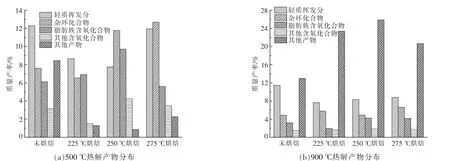
圖9 小球藻及烘焙樣品在不同溫度下的熱解產物分布Fig.9 Distribution of pyrolysis products of Chlorella and torrified samples at different temperatures
由圖9 可以看到, 熱解溫度為900 ℃下的熱解產物中,輕質揮發分、雜環化合物、含氧化合物所占比例均低于500 ℃下的熱解產物(烘焙溫度為250 ℃時的輕質揮發分除外),然而其他產物的占比大幅度增加,導致最終生物油的產率增大。這主要是由于500 ℃下原先檢測不到的重質生物油在熱解溫度提升至900 ℃的高溫環境下發生裂解,生成了更多以輕質芳烴和不飽和烴類物質(碳原子個數小于6)為主的輕質揮發分。
小球藻的熱解產物中含氮產物較多, 含氮物質主要包含在雜環類物質中, 這是由于藻類的構成與木質纖維素類生物質不同,蛋白質、脂類、糖類是其主要成分[11],而蛋白質中的氨基酸側鏈是N 元素的主要來源。 對于在500 ℃下的熱解產物而言,雜環類化合物主要由吲哚、吡咯類、喹啉類、吡啶類、肽類和吡喃類組成,脂肪族含氧化合物主要由乙酸、羥基丙酮、長鏈脂肪酸及其衍生物長鏈脂肪酰胺類組成, 其他含氧化合物主要由苯酚類組成,此外還有部分C-O-N=C 結構及酯類結構,其他產物主要由胺類、甲苯類、腈類(C≡N 鍵)等組成。在900 ℃下的熱解產物,不同之處在于其他產物中的種類增加,主要由苯、甲苯、萘、茚等芳香類產物以及二烯類、丙烯腈等組成。
當小球藻在500 ℃下熱解時,由圖9 可以看出:隨著烘焙溫度的升高,小球藻熱解產物中的雜環化合物逐漸增加,只有烘焙溫度為225 ℃時的產率低于未經烘焙預處理小球藻原樣;含氧化合物先增加后減少,在烘焙溫度為250 ℃時達到最高; 其他產物先減少后增加, 在烘焙溫度為250 ℃時產率達到最低, 且均低于未經烘焙預處理原樣。 當小球藻在900 ℃下熱解時,雜環化合物占比在烘焙溫度為250 ℃時達到最低,略高于未經過烘焙預處理的原樣產率;含氧化合物和其他產物隨烘焙溫度的升高先增加后減少,在烘焙溫度為250 ℃時達到最高,二者的不同之處在于含氧化合物的所占比例變化不大,而其他產物遠遠高出未烘焙原樣的產率。 這說明,高溫下重質生物油的熱裂解過程幾乎不會產生含氧化合物,以輕質芳烴和烯烴類為主, 經過烘焙預處理后,更容易發生重質生物油的裂解,最佳烘焙溫度為250 ℃。
3 結論
本文通過TG-FTIR 與Py-GC/MS 方法,探究了烘焙預處理和不同烘焙溫度對小球藻熱解動力學及其產物的影響,得到以下結論。
①烘焙預處理的峰值失重溫度隨烘焙溫度的升高而升高,同時兩個主要失重峰的差距減小,提高了原料組分的均一性; 提升烘焙溫度能降低部分活化能, 但烘焙處理后的小球藻總體熱解反應活化能仍會增加。
②烘焙預處理促進了小球藻熱解脫羧過程,產生更多CO2; 同時會使小球藻在熱解過程中含氮產物的生成顯著增加, 提升烘焙溫度能夠抑制這一趨勢。
③小球藻的熱解生物油中含氮物質較多,以吲哚、吡咯類、喹啉類為主,隨著烘焙溫度的升高,生物油中的雜環化合物比例增加。 當小球藻在900 ℃下發生熱解時,以生成輕質芳烴和烯烴類產物為主,幾乎不產生含氧化合物,且烘焙預處理更容易使生物油發生裂解,最佳烘焙溫度為250 ℃。
