基于內殼生長紋的浙江海域曼氏無針烏賊生長特性
2022-08-18 02:40:48謝慕原李鵬飛徐漢祥徐開達劉連為隋宥珍王雅麗
浙江海洋大學學報(自然科學版) 2022年4期
關鍵詞:生長
謝慕原,李鵬飛,徐漢祥,徐開達,劉連為,隋宥珍,王雅麗
(1.浙江海洋大學海洋與漁業研究所,浙江省海洋水產研究所,農業農村部重點漁場漁業資源科學觀測試驗站,浙江省海洋漁業資源可持續利用技術研究重點實驗室,浙江舟山 316021;2.浙江海洋大學水產學院,浙江舟山 316022)
曼氏無針烏賊Sepiella maindroni,俗稱墨魚,屬于軟體動物門Mollusca,在我國沿海地區分布廣泛,主要集中在浙江和福建沿海,一般以浙北群體與浙南群體之稱將其區分開來[1],是除了大黃魚Larimichthys crocea、小黃魚L.polyactis和帶魚Trichiurus lepturus以外的“四大海產”之一[2]。曼氏無針烏賊是沿海地區餐桌上的傳統美食佳肴,曾在市場中占有重要份額,其食用口感純正、營養豐富,深受消費者喜愛。自20 世紀70 年代末以來,由于漁業管理制度的不完善,漁民過度捕撈破壞了沿海生態平衡,導致曼氏無針烏賊資源逐步衰竭,并無法形成漁汛[3-4]。近年來,隨著苗種繁育技術的突破和規模化增殖放流的持續實施[5-11],曼氏無針烏賊資源呈現恢復的跡象[12]。
為更精確地掌握規模化增殖放流背景下自然海域曼氏無針烏賊群體的生長動態,提供更科學的增殖放流管理建議,有必要開展自然海域曼氏無針烏賊生長現狀研究。目前,有關于頭足類日齡與生長的研究大多是以耳石作為研究材料,通過測量與統計耳石的輪紋與其胴背長等系數之間的相關性來計算出生長率,由于烏賊耳石無明顯核心且生長紋不清晰,經打磨處理后雖然生長紋的清晰度提高,但是會因個別生長紋因打磨形成斷層,造成統計誤差,因此,可以選擇通過誤差較小、生長紋清晰的內殼輪紋對部分頭足類的生長率進行計算,通過實驗發現,其結果相較于耳石的輪紋也更貼近實際生長情況[13-16]。瞿俊躍等[17]通過西非沿岸烏賊內殼生長紋來判斷日齡,結合內殼日齡信息,對不同性別烏賊生長特性進行了研究。SCHROEDER,et al[18]對阿根廷滑柔魚Illex argentinus的耳石與其內殼的研究中發現內殼的生長紋增量速度與耳石相同,其生長紋的積累是日周期性。但通過謝慕原等[19]對養殖環境下曼氏無針烏賊日齡與生長紋輪紋數關系的研究發現,其內殼輪紋的生長與其他頭足類研究結果不一致,不符合嚴格的“一日一輪”增長規律,水溫、性腺發育等較多因素都可以影響內殼輪紋的生長,在可調控溫度的養殖環境下,采用內殼輪紋數來鑒定日齡不夠穩定,而在不可調控溫度、食物狀況等復雜的海洋環境下,內殼輪紋的生長可能更加不穩定。由此,內殼生長紋雖然不符合“一日一輪”,但依然可以展現烏賊生長的一般特性,本研究選用浙江海域野生曼氏無針烏賊的內殼作為研究對象,觀察并計數其內殼表面的生長輪紋,分析不同海域曼氏無針烏賊胴背長、體重以及內殼輪紋數的組成,探討內殼長度與胴長和體質量之間的關系以及內殼輪數與各形態指標的關系。
1 材料與方法
1.1 樣品采集
根據周永東等[1]調查歷史數據等資料發現,我國近海曼氏無針烏賊洄游距離短,一般分為浙北種群與浙南種群,故本研究共采集曼氏無針烏賊樣品307 尾,其中浙北海域樣本數205 尾,浙南海域樣本數102 尾,采集時間均為5-12 月(除7 月禁漁期外),其胴背長范圍為2.7~13.7 cm,體重范圍為5.16~293.9 g,輪數范圍為43~145 輪(圖1)。
圖1 采樣站位示意圖Fig.1 Sampling stations
1.2 生物學與內殼測量
將采集的曼氏無針烏賊樣本帶回實驗室解凍后,將浙北海域與浙南海域樣本分開進行測定,并對其的胴背長(ML)、體重(W)、內殼長(GL)、內殼寬(GW)等生物學特性進行測定。胴背長測定精確至0.1 cm,體質量等測定精確至0.01 g。解剖后取出內殼,使用清潔劑清洗存留在內殼表面的有機物,測量內殼殼長、殼寬等精確至0.1 cm,將內殼做好標記備用。在尼康體視解剖鏡下測定內殼輪紋數(N)并采集圖像。
1.3 數據分析
為分析各形態指標與胴長的關系,分別應用線性、冪、指數、對數、增長和邏輯斯蒂[20-22]等函數擬合兩者之間的關系,決定系數(R2)最大者為最佳擬合回歸方程[23]。
擬合模型:
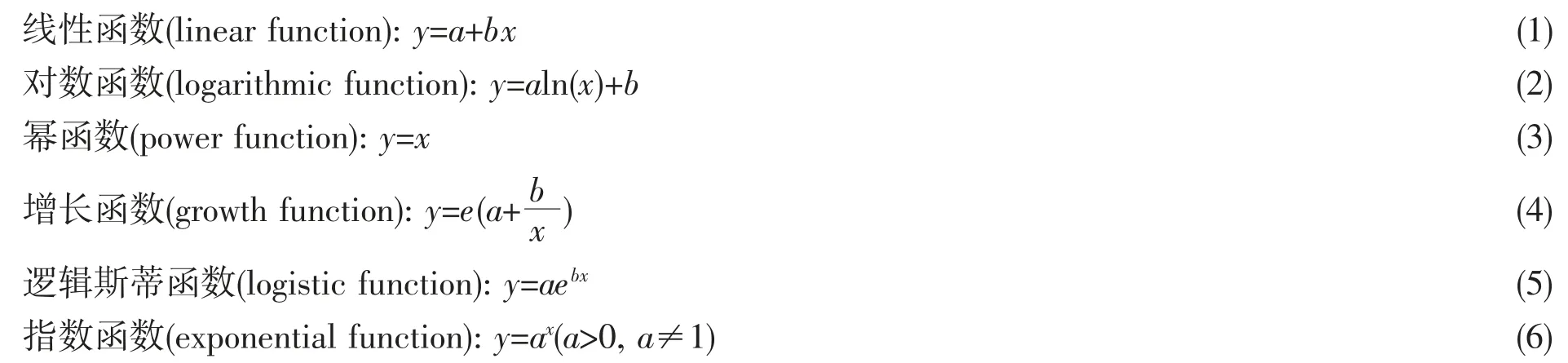
利用SPSS 17.0 軟件對數據進行獨立樣本t 檢驗,對曼氏無針烏賊胴背長、體重、內殼輪紋數等數據進行分析,檢驗其是否存在差異。
2 結果
2.1 群體規格組成
經檢驗,浙北與浙南海域曼氏無針烏賊胴背長與體重均無顯著性差異(P=0.062>0.05)(圖2、3),浙北海域烏賊胴背長分布范圍為27~137 mm,平均胴長為74 mm,標準差25.12,優勢胴背長組為81~100 mm,占整體樣本的33%,體重分布范圍為5.16~288.62 g,平均體重為69.24 g,標準差53.77,優勢體重組為0~30 g,占整體樣本的33%,211~270 是占樣本量總數最少的區間;浙南海域烏賊胴背長分布范圍為30~129 mm,平均胴背長為71 m,標準差23.9,優勢胴背長組為41~60 mm,占整體樣本的49%,體重分布范圍為5.14~154.23 g,平均體重為35.58 g,標準差為46.8,優勢體重組為0~30 g,占整體樣本的44%。
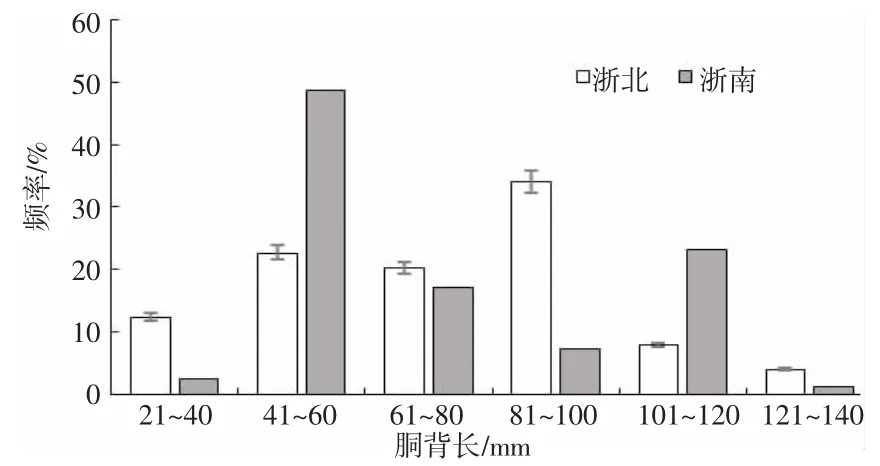
圖2 不同海域烏賊的胴背長分布Fig.2 Distribution of mantle length of S.maindroni in different sea areas
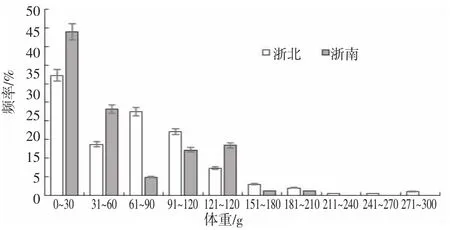
圖3 不同海域烏賊的體重分布Fig.3 Distribution of weight of S.maindroni in different sea areas
2.2 輪數組成
由于個別曼氏無針烏賊內殼取出時已經破損,本實驗共取出浙北群體內殼192 個,浙南群體內殼100個。經檢驗,浙北與浙南群體內殼輪紋數存在顯著差異(P=0.000<0.05),浙北群體內殼輪數分布范圍為58~145輪,平均輪數為93.64 輪,標準差19.48,優勢輪數組為101~120 輪,占整體樣本的35%,占總樣本量最小的組是41~60 輪和141~160 輪;而浙南群體輪數分布范圍為43~138 輪,平均輪數為86.8 輪,標準差為20.17,優勢輪數組為61~80 輪,占整體樣本的45%(圖4)。
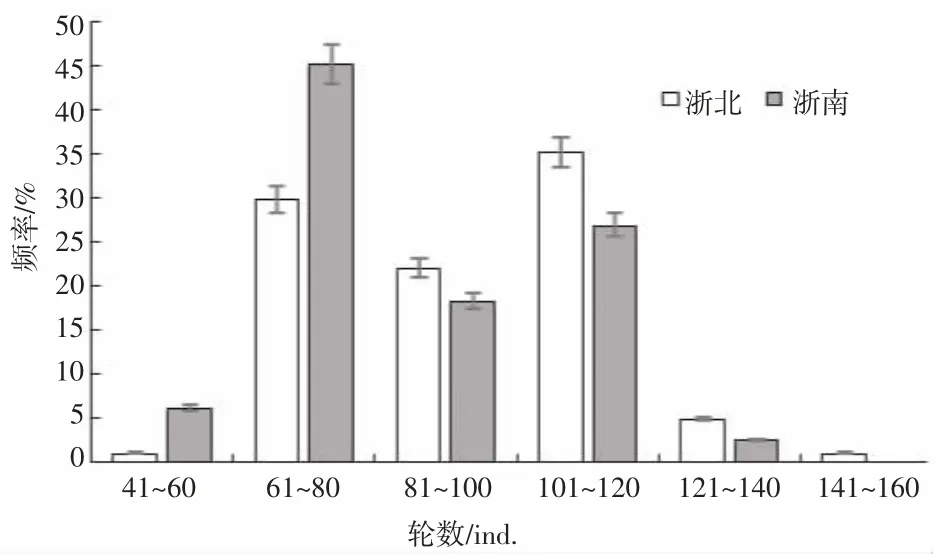
圖4 不同海域烏賊的輪紋數分布Fig.4 Distribution of wheel grain number of S.maindroni in different sea areas
2.3 曼氏無針烏賊胴背長、體重與內殼長的關系
浙北海域曼氏無針烏賊內殼與胴背長呈明顯的線性關系(R2=0.984 8),與體重呈冪函數關系(R2=0.951 4);胴背長與體重呈冪函數關系(R2=0.941 7)。浙南海域曼氏無針烏賊內殼與胴背長呈明顯的線性關系(R2=0.972 8),與體重呈冪函數關系(R2=0.944 8);胴背長與體重呈冪函數關系(R2=0.955 9)(表1、圖5)。

表1 內殼長、胴背長和體重的關系Tab.1 Relationship among cuttlebone length,mantle length and body weight
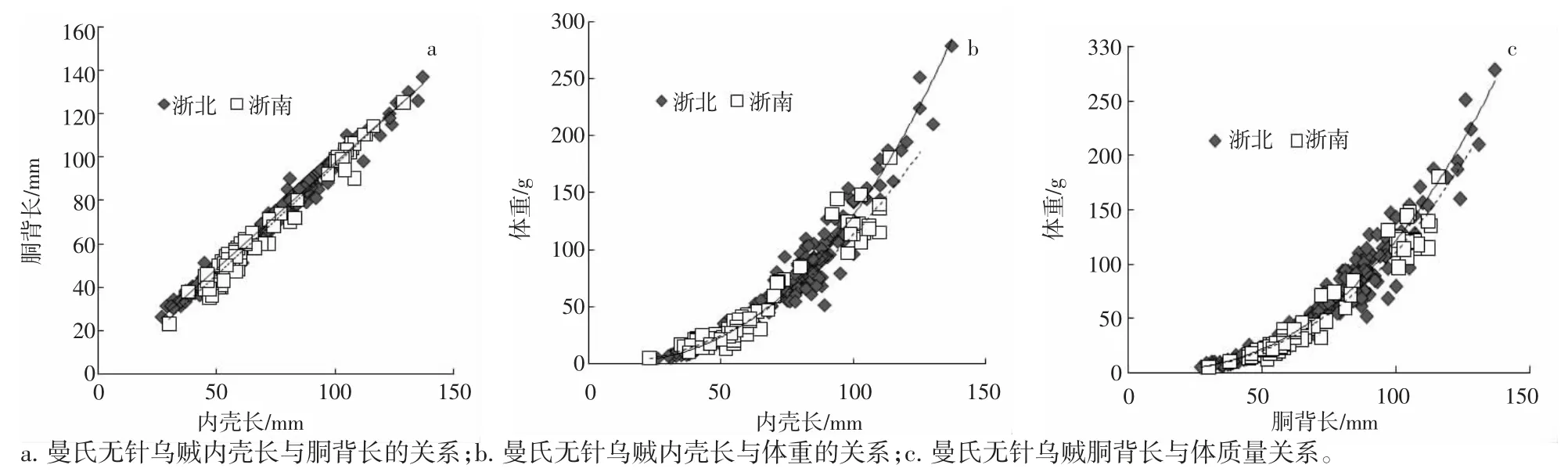
圖5 曼氏無針烏賊內殼長度、胴背長和體重的關系Fig.5 The relationship among the cuttlebone length,the mantle length and the weight of S.maindroni
2.4 曼氏無針烏賊內殼輪數與各形態指標的關系
為進一步分析曼氏無針烏賊內殼輪紋數(N)與各形態指標的關系,將不同海域烏賊ML、W、GL、GW 的生長模型分開研究。根據表2 的結果,比較不同生長模型的判定系數(R2),確定不同地區烏賊群體ML-N、W-N、GL-N、和GW-N 的最優生長模型:
浙北海域:

浙南海域:

結果表明:浙北海域烏賊ML、W、GL、GW 與N 的擬合均以冪函數模型最優,而浙南海域烏賊ML-N和GL-N 以冪函數最優,W-N 和GW-N 以線性函數擬合最優(表2)。
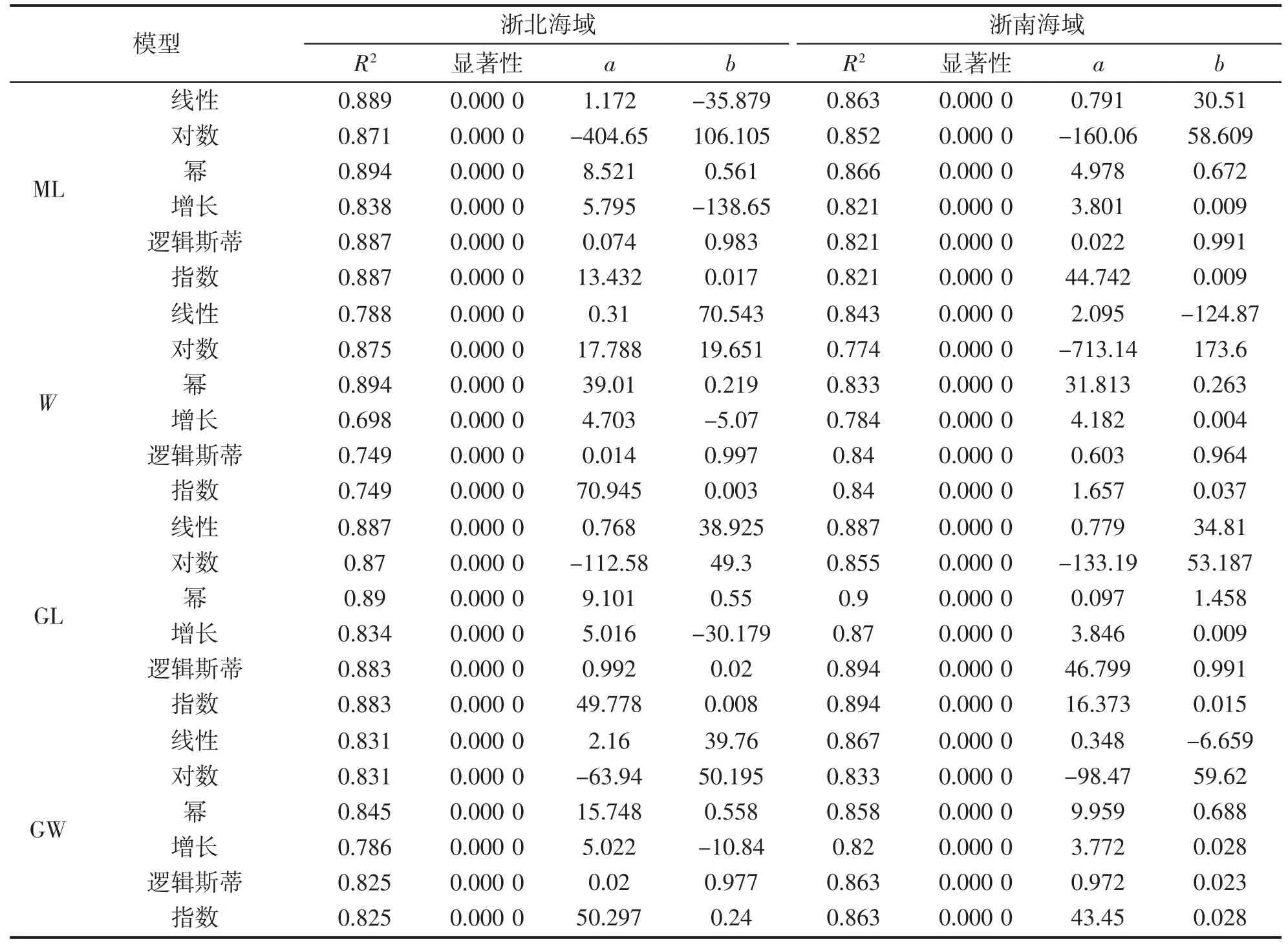
表2 浙江地區曼氏無針烏賊本形態指標與內殼輪紋模型曲線估計結果Tab.2 Model curve estimation results of the model curve between the morphologic indexes and N of S.maindroni in Zhejiang province
3 討論
3.1 群體規格的差異及變化
由于漁業監督與管理中的不足,漁業生產出現了不規范及過度捕撈行為,一些性腺尚未成熟的幼烏賊也被捕撈出海,使得曼氏無針烏賊資源衰竭。近幾年,通過持續地增殖放流曼氏無針烏賊苗種的方式,使得其資源得到一定程度的恢復[24],但與1960 年倪正雅等[25-26]研究發現相比,曼氏無針烏賊體型大小呈下降趨勢,整體小型化較為明顯,其中1960 年浙北曼氏無針烏賊生殖群體最大胴背長為190 mm,最大體重達580 g,而本研究中浙北烏賊群體最大胴背長只有137 mm,體重為294 g,甚至沒有達到1960 年烏賊胴背長和體重的平均值;而1960 年的浙南漁場生殖群體體重高達660 g,胴背長與體重的平均值同樣遠遠高于本研究中2020-2021 年的浙南群體。吳常文等[27]在對曼氏無針烏賊繁殖生物學特性研究時發現,20 世紀80 年代初性成熟的烏賊胴背長和體重要大于60 年代烏賊的體型規格,而80 年代初至今,性成熟的烏賊胴背長、體重在逐漸下降,說明漁場生態是動態變化的,多種因素的變化都會導致其出現連鎖性反應。此外,浙江南部和北部海域海洋環境有一定的差異,且食物網組成以及物種組成存在較大的差異,從而為曼氏無針烏賊提供了迥異的生長環境、棲息地以及食物來源,這可能是導致2 個海域的烏賊胴背長組成有較大差異的原因之一。
3.2 內殼生長輪紋組成
劉必林等[28-32]研究表明,烏賊類內殼生長紋是否具有日周期性仍存在一些爭議,不是所有學者都認為生長輪紋的沉積周期性與其生活環境的溫度息息相關。本研究也曾選用烏賊內殼表面生長輪紋作為研究對象分析其日齡,但BETTENCOURT,et al[19,33]從實驗結果中發現烏賊的全部生命周期,其內殼輪紋并不是勻速生長,內殼輪紋的生長受水環境尤其是溫度變化及自身發育等多種因素的影響,由此說明內殼生長紋不符合“一日一紋”,故通過內殼表面生長輪紋無法準確判斷烏賊的日齡。此外,本研究中烏賊內殼生長輪紋最高達到140~160 區間,而倪正雅等研究發現1960 年浙南浙北海域烏賊內殼輪紋數可達170~180 輪,比本研究中多了20 輪左右。曾經有學者認為烏賊的內殼輪紋數與生活環境、棲息地的溫度密切相關,且近年來海洋溫度在不斷地發生變化,這可能導致烏賊生長紋的減少的原因之一。
3.3 曼氏無針烏賊各項生長指標與內殼長及輪紋數擬合關系比較
研究發現,目前無法通過內殼來準確判斷曼氏無針烏賊的日齡,但內殼生長與烏賊自身的生長密不可分,本研究嘗試用胴背長和體重與內殼長的擬合關系來分析其生長,在用幾種函數模型的擬合對比后,發現曼氏無針烏賊內殼長與胴背長線性模型擬合結果最優,此結果與WANG Xuefeng,et al[34]和郭有俊等[35]在研究鳶烏賊Sthenoteuthis oualaniensis時發現其內殼長與胴背長呈線性相關相似。另外,浙北與浙南海域群體無論是胴背長還是內殼長,與其體重的關系均以冪函數模型擬合最佳,但李楠等[36]在對劍尖槍烏賊Loligo edulis進行研究時發現,烏賊群體胴背長與體重關系在春季產卵時是以Logistic 曲線擬合最佳,而夏季產卵群體胴背長與體重的關系分別以von-Bertalanffy 和Logistic 曲線擬合最佳,這可能是因為劍尖槍烏賊體型較為狹長且分布在外海,而曼氏無針烏賊體型為橢圓狀且分布在近海,不同的棲息環境、餌料和海洋溫度等因素都會影響其形態之間的差異,故擬合結果不一致,后續可以通過增加不同季節的產卵群體的取樣量,對曼氏無針烏賊胴背長和體重等關系做進一步比較。實驗發現烏賊解凍后肉質較軟,或經外力壓迫后體型容易有所改變,可能會在測量時出現一定的誤差,而烏賊內殼是體內的硬組織,形態穩定,不易受到破壞,測量誤差相較胴背長要低很多,所以在烏賊受到外力擠壓后,胴背長無法很好測量時,可以選用內殼長來替換胴背長作為分析指標。由此可見,內殼長與胴背長都可以用來分析生長關系。浙北海域曼氏無針烏賊內殼輪紋數與主要外形參數均為冪函數擬合最優,這與之前學者[19]對養殖曼氏無針烏賊的研究結果一致。這一結果可能是由于近些年在曼氏無針烏賊增殖放流工作加大的影響下,養殖群體的養殖單位近幾年來,在浙北海域每年放流受精卵2 000 萬單位左右;另外,根據徐開達等[12]利用2013-2016 年的放流數據的分析發現曼氏無針烏賊受精卵的放流達到種群修復和增產增收的目的,因此會造成浙北海域取樣的烏賊的研究結果和養殖群體的比較相近;而浙南海域由于種群的差異,內殼輪紋數與主要外形參數并不是均以冪函數最優。此研究結果還需后續的實驗研究來進一步拓展并驗證不同海域間曼氏無針烏賊的差異。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
