熟地黃對6-羥基多巴胺誘導的多巴胺能神經細胞氧化應激的影響
2022-08-25 13:20:36王家鵬孫曉杰
中國老年學雜志 2022年16期
王家鵬 孫曉杰
(山東中醫藥高等專科學校 1中醫系,山東 煙臺 264199;2招生就業處)
帕金森病是一種神經退行性疾病,其病理特征為腦黑質致密區域多巴胺能神經元缺失,神經元缺失可誘導紋狀體多巴胺異常減少,誘導基底神經節異常,進而誘發行動遲緩、平衡功能障礙、肢體僵硬等運動癥狀,氧化應激被認為是多巴胺能神經細胞功能紊亂的重要原因〔1〕。目前常見的帕金森病治療藥物有多巴胺受體激動藥、抗膽堿藥等,這些藥物多數為對癥治療,只能部分改善運動缺陷,且不能阻止病情進展,研發有效藥物改善帕金森病備受關注〔2〕。熟地黃是玄參科植物地黃塊根經過炮制加工而成,具有抗衰老、抗氧化、改善記憶力、調節免疫等作用〔3〕。有研究〔4〕顯示,熟地黃能夠緩解帕金森病模型大鼠異動癥狀。現階段尚不清楚熟地黃對6-羥基多巴胺(OHDA)誘導的多巴胺能神經細胞的影響。本實驗研究熟地黃對6-OHDA誘導的多巴胺能神經細胞氧化應激的影響和機制。
1 材料與方法
1.1材料 細胞:多巴胺能神經細胞SH-SY5Y(貨號:258028)購自寧波明舟生物科技有限公司。多巴胺能神經細胞SH-SY5Y培養在含有10%胎牛血清的DMEM細胞培養液內,細胞培養條件為37℃,飽和濕度,5% CO2培養箱。藥物:熟地黃(北京索萊寶科技有限公司,編號:DYS8270,規格:1 g)。試劑與儀器:6-OHDA(Sigma公司,美國,貨號:H4381-100MG);p38MAPK激活劑Anisomycin(Sigma公司,美國,貨號:A5862-0.5ML);兔抗p-p38MAPK抗體(Cell Signaling Technology公司,美國,貨號:4511);活性氧(ROS)檢測試劑盒(北京百奧萊博科技有限公司,貨號:HR8786);兔抗剪切的含半胱氨酸的天冬氨酸蛋白水解酶(酶切Caspase)-3抗體(Abcam公司,美國,貨號:ab2302);兔抗p38MAPK抗體(北京百奧萊博科技有限公司,貨號:YT820);兔抗Ki-67抗體(Abgent公司,美國,貨號:A-AJ1427c);丙二醛(MDA)檢測試劑盒(北京索萊寶科技有限公司,貨號:BC0020);DMEM培養基(Sigma公司,美國,貨號:DF-042-B);兔抗Bcl-2相關X蛋白(Bax)抗體(Abcam公司,美國,貨號:ab53154);超氧化物歧化酶(SOD)檢測試劑盒(碧云天生物技術研究所,貨號:S0101S);胎牛血清(浙江天杭生物科技股份有限公司,貨號:11011-8611);乳酸脫氫酶(LDH)檢測試劑盒(碧云天生物技術研究所,貨號:C0017);流式細胞儀(Becton,Dickinson and Company公司,美國,LSRFortessa X-20型);酶標儀(北京普凱瑞生物科技有限公司,SpectraMax iD5型)。
1.2分組及給藥方法 取處于對數生長期的多巴胺能神經細胞,根據不同的實驗需求接種到細胞培養板內,24 h后,添加不同濃度的含藥培養液,對照組細胞培養液不添加任何藥物,6-OHDA組、熟地黃低劑量組、熟地黃中劑量組、熟地黃高劑量組、熟地黃高劑量+Anisomycin組細胞培養液中均添加100 μmol/L的6-OHDA〔5〕,同時熟地黃低、中、高劑量組細胞培養液中分別添加120、240、480 μmol/L的熟地黃,熟地黃高劑量+Anisomycin組細胞培養液中添加480 μmol/L的熟地黃和2 ng/ml的p38MAPK激活劑Anisomycin〔6〕。各組細胞培養24 h后進行相關檢測。
1.3檢測指標與方法 (1)CCK-8實驗檢測細胞增殖:多巴胺能神經細胞按照每個孔內4 000個細胞接種到96孔板內,細胞培養24 h后,分別在每個孔內添加10 μl CCK-8溶液,繼續孵育10 min,用酶標儀測定波長為450 nm的吸光度(OD)值,以OD值表示細胞增殖活性。(2)Western印跡檢測相關蛋白表達:多巴胺能神經細胞按照每個孔內接種5×104個細胞接種到24孔板內,24 h后,用磷酸鹽緩沖液(PBS)洗滌細胞2次,添加含10%苯甲基磺酰氟(PMSF)的放射免疫沉淀(RIPA)溶液,放在冰上裂解,30 min后,在4℃條件下高速離心,收集蛋白上清,經二喹啉甲酸(BCA)方法檢測蛋白濃度后,與2倍結合緩沖液混合煮沸5 min。在每個十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳(SDS-PAGE)上樣孔內添加40 μg蛋白樣品,先以90 V的電壓電泳30 min后,再以120 V的電壓電泳1.5 h。以200 mA恒流轉膜1.5 h。將NC膜放置5%牛血清白蛋白溶液中封閉2 h。NC膜放在一抗溶液(p-p38MAPK、酶切Caspase-3抗體以1∶800稀釋,p38MAPK抗體以1∶1 500稀釋,Ki-67、Bax抗體以1∶1 000稀釋)中,4℃搖床過夜。NC膜放在1∶2 000稀釋后的二抗溶液內,在室溫中結合2 h。ECL發光。Image J分析比較灰度值,以GAPDH校正。(3)流式細胞術檢測凋亡:多巴胺能神經細胞按照每個孔內接種5×104個細胞接種到24孔板內,24 h后,收集細胞,用PBS洗滌細胞2次,每個測定樣品取1×106個細胞,懸浮于500 μl的結合緩沖液中,分別添加5 μl的膜連蛋白(Annexin)V-FITC和碘化丙啶(PI)溶液,轉移至避光條件下結合20 min,用流式細胞儀檢測凋亡變化。(4)ROS、MDA、SOD含量和培養液上清中LDH檢測:多巴胺能神經細胞按照每個孔內接種5×104個細胞接種到24孔板內,24 h后,收集細胞和培養液上清,以ROS檢測試劑盒(CellROX Green熒光探針法)、MDA檢測試劑盒(可見分光光度法)、SOD檢測試劑盒(黃嘌呤氧化法)分別測定細胞中ROS、MDA、SOD含量,ROS檢測結果以對照組為參照,分析其余各組ROS含量變化百分比;以LDH檢測試劑盒(比色法)測定培養液上清中LDH含量,步驟均按照試劑盒說明書進行。
1.4統計學方法 采用SPSS23.0軟件進行單因素方差分析。
2 結 果
2.1熟地黃對多巴胺能神經細胞增殖影響 與0 μmol/L(0.56±0.06)比較,120、240、480 μmol/L的熟地黃處理后多巴胺能神經細胞增殖活性(0.51±0.08、0.49±0.06、0.47±0.07)沒有變化(P>0.05);960、1 920 μmol/L熟地黃處理后多巴胺能神經細胞增殖活性(0.40±0.06、0.32±0.04)顯著降低(P<0.05)。選擇對多巴胺能神經細胞沒有毒性的120、240、480 μmol/L的熟地黃進行后續實驗。
2.2熟地黃可抑制6-OHDA誘導的多巴胺能神經細胞中p38MAPK信號激活 與對照組(0.15±0.03)比較,6-OHDA組多巴胺能神經細胞中p-p38MAPK/p38MAPK表達量(0.89±0.09)顯著升高(P<0.05);與6-OHDA組比較,熟地黃低、中、高劑量組表達量(0.63±0.05、0.41±0.03、0.29±0.02)逐漸顯著降低(P<0.05);與熟地黃高劑量組比較,熟地黃高劑量+Anisomycin組表達量(0.45±0.06)顯著升高(P<0.05)。見圖1。
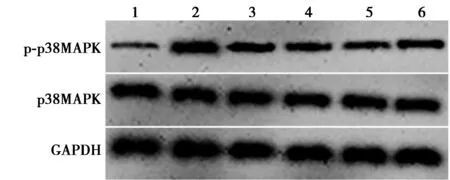
1~6:對照組、6-OHDA組、熟地黃低劑量組、熟地黃中劑量組、熟地黃高劑量組、熟地黃高劑量+Anisomycin組;同下圖圖1 各組多巴胺能神經細胞中p38MAPK信號 相關蛋白表達
2.3熟地黃調控p38MAPK對6-OHDA誘導的多巴胺能神經細胞增殖、凋亡影響 與對照組比較,6-OHDA組多巴胺能神經細胞增殖活性顯著降低,凋亡率顯著升高(P<0.05);與6-OHDA組比較,熟地黃低、中、高劑量組多巴胺能神經細胞增殖活性逐漸顯著升高,凋亡率逐漸顯著降低(P<0.05);與熟地黃高劑量組比較,熟地黃高劑量+Anisomycin組多巴胺能神經細胞增殖活性顯著降低,凋亡率顯著升高(P<0.05)。見圖2,表1。
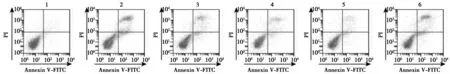
圖2 各組多巴胺能神經細胞凋亡情況
表1 各組多巴胺能神經細胞增殖活性和凋亡率及Ki-67、Bax、酶切Caspase-3蛋白表達量比較
2.4熟地黃調控p38MAPK對6-OHDA誘導的多巴胺能神經細胞增殖、凋亡相關蛋白表達影響 與對照組比較,6-OHDA組多巴胺能神經細胞中Ki-67蛋白表達量顯著降低,Bax、酶切Caspase-3蛋白表達量顯著升高(P<0.05);與6-OHDA組比較,熟地黃低、中、高劑量組多巴胺能神經細胞中Ki-67蛋白表達量逐漸顯著升高,Bax、酶切Caspase-3蛋白表達量逐漸顯著降低(P<0.05);與熟地黃高劑量組比較,熟地黃高劑量+Anisomycin組多巴胺能神經細胞中Ki-67蛋白表達量顯著降低,Bax、酶切Caspase-3蛋白表達量顯著升高(P<0.05)。見表1,圖3。
2.5熟地黃調控p38MAPK對6-OHDA誘導的多巴胺能神經細胞氧化應激影響 與對照組比較,6-OHDA組多巴胺能神經細胞中SOD含量顯著降低,ROS、MDA含量顯著升高,培養液上清中LDH含量升高(P<0.05);與6-OHDA組比較,熟地黃低、中、高劑量組多巴胺能神經細胞中SOD含量逐漸顯著升高,ROS、MDA含量逐漸顯著降低,培養液上清中LDH含量逐漸顯著降低(P<0.05);與熟地黃高劑量組比較,熟地黃高劑量+Anisomycin組多巴胺能神經細胞中SOD含量顯著降低,ROS、MDA含量顯著升高,培養液上清中LDH含量顯著升高(P<0.05)。見表2。

圖3 各組各組多巴胺能神經細胞中增殖和凋亡 相關蛋白表達
表2 各組多巴胺能神經細胞中ROS、MDA、SOD含量和培養液上清中LDH含量比較
3 討 論
帕金森病屬于神經退行性疾病,多發生于中老年人,也是中老年殘疾的重要誘因〔7〕。帕金森病常見病理變化為腦黑質致密帶多巴胺神經細胞脫落、缺失,目前尚不清楚帕金森病發生的原因,遺傳因素、興奮性因素、自身免疫因素、氧化應激、環境因素等均與其有關,其中氧化應激因素研究最多〔8,9〕。6-OHDA是常見的體外構建帕金森病細胞模型的誘導因子〔10〕。帕金森病條件下,多巴胺能神經細胞中合成大量ROS,這些ROS不能被抗氧化酶SOD等及時清除,誘導ROS聚集,而過量的ROS可促進脂質發生過氧化,生成MDA;脂質是細胞膜的重要組成部分,其過氧化后可誘導細胞膜完整性破壞,導致原本多存在于細胞內的LDH釋放至細胞外,因此檢測MDA、LDH水平可間接反映氧化應激水平〔11,12〕。過量的ROS還可以激活細胞凋亡蛋白如Bax、酶切Caspase-3,促進細胞凋亡發生〔13〕。Ki-67是細胞增殖標志蛋白,其表達變化與細胞增殖水平有關〔14〕。本實驗結果表明,6-OHDA誘導多巴胺神經細胞氧化應激,激活細胞凋亡,成功構建了體外帕金森病細胞模型。
熟地黃是常見中藥材,具有滋陰補腎的功效,現代藥理學證明熟地黃有調節免疫力、改善骨質疏松、降血糖、抗衰老、抗氧化、改善腎功能等作用〔15〕。熟地黃對神經系統有保護作用,可以抑制阿爾茨海默病大鼠海馬神經細胞損傷,減少Caspase-3活化〔16〕;熟地黃治療后的注意缺陷多動障礙大鼠神經元發育明顯改善〔17〕。熟地黃對帕金森病可能有治療功效,復方地黃處理后的帕金森病動物模型運動障礙癥狀減輕〔18〕;熟地黃處理后的MPP+誘導的神經細胞凋亡減少,Bax和酶切Caspase-3蛋白表達減少〔19〕。本實驗結果說明熟地黃能夠減輕6-OHDA誘導的多巴胺能神經細胞氧化應激,進而減少細胞凋亡,與以前的研究報道結果一致〔12〕,均提示熟地黃可能是帕金森病治療的有效藥物。
熟地黃作用機制與信號途徑的轉導有關〔20〕。本實驗發現,熟地黃處理可降低多巴胺能神經細胞中p38MAPK磷酸化水平,熟地黃作用機制可能與p38MAPK有關。p38MAPK是MAPK信號通路的重要分支,在細胞增殖、氧化應激、細胞凋亡、細胞衰老、炎癥、能量代謝過程中發揮作用〔21〕。有研究〔22〕表明,p38MAPK在帕金森病進展中過度激活,p38MAPK具有促進帕金森病發生的作用。p38MAPK在6-OHDA誘導的多巴胺能神經細胞中過度磷酸化,且抑制p38MAPK可改善多巴胺能神經細胞損傷〔23〕。本文以p38MAPK激活劑進一步驗證熟地黃的作用機制,結果發現,p38MAPK激活劑能夠逆轉熟地黃對6-OHDA誘導的多巴胺能神經細胞增殖、凋亡和氧化應激的作用,提示熟地黃通過抑制p38MAPK信號發揮作用。
綜上,熟地黃可能是帕金森病治療的潛在藥物,其能夠通過抑制p38MAPK信號改善6-OHDA誘導的多巴胺能神經細胞氧化應激,進而減少細胞凋亡。關于熟地黃通過何種具體靶向機制影響p38MAPK信號進而發揮作用還有待進一步研究。
猜你喜歡
課堂內外·初中版(科學少年)(2023年10期)2023-12-10 00:43:06
全科護理(2022年10期)2022-12-26 21:19:15
中國合理用藥探索(2022年1期)2022-11-26 00:22:32
保健醫苑(2022年6期)2022-07-08 01:25:28
中國民間療法(2021年10期)2021-07-22 02:32:40
成都醫學院學報(2021年2期)2021-07-19 08:35:14
鄉村科技(2021年33期)2021-03-16 02:26:54
國際放射醫學核醫學雜志(2021年10期)2021-02-28 08:41:58
活力(2019年22期)2019-03-16 12:47:04
醫學研究雜志(2015年5期)2015-06-10 06:43:26
