中華絨螯蟹4個養殖群體遺傳多樣性與遺傳結構分析
2022-08-27 10:59:56胡玉婷潘庭雙周華興
江蘇農業科學 2022年16期
胡玉婷, 凌 俊, 江 河, 汪 煥, 潘庭雙, 周華興
(安徽省農業科學院水產研究所/水產增養殖安徽省重點實驗室,安徽合肥 230031)
中華絨螯蟹()別稱河蟹,俗稱大閘蟹,是我國久負盛名的名貴水產品,自然分布于我國長江、黃河、遼河和甌江等河川及附近地區。20世紀80年代初,隨著河蟹育苗技術的突破和養殖技術的開發和推廣,我國河蟹養殖進入前所未有的嶄新階段,產量逐年增加,已成為世界首個產蟹大國,同時從蟹苗一直到成蟹培育,全部采用人工方法,這在世界上也是絕無僅有的。目前,中華絨螯蟹養殖已遍布我國從遼寧至浙江的各沿海省份及廣大內陸地區,是我國最重要的經濟養殖蟹類,2020年全國養殖總產量為77.6萬t。但隨著中華絨螯蟹養殖產業的規模化發展,跨流域引種和增殖放流等生產活動,造成不同水系中華絨螯蟹種質資源嚴重混雜和養殖性能退化,尤其受利益驅使和技術所限,中華絨螯蟹育苗場一般選擇100 g左右小規格親本繁殖,且不考慮親本遺傳差異,導致蟹苗質量差,已成為當前養殖生產發展的瓶頸問題。如蘇雨等采用微衛星和線粒體標記(COⅠ、b和D-loop)分析了來自長江、黃河和遼河野生與養殖群體的遺傳多樣性和遺傳結構,結果一致表明,三個水系群體間的遺傳分化程度較低,野生群體的遺傳分化與地理距離具有不完全相關性,提示不同水系的中華絨螯蟹可能與其跨流域引種及養殖群體逃逸造成其種質混雜有關。
一般認為,遺傳多樣性的大小及其群體遺傳結構跟一個物種的進化潛力和抵御不良環境的能力密切相關。鑒于此,本研究擬在分析養殖群體遺傳背景的基礎上,從中選取遺傳多樣性高且遺傳距離適度的群體作為育種基礎群,采取大規格親本進行繁育的方法培育中華絨螯蟹良種。江蘇和安徽兩省是目前中華絨螯蟹重要的養殖區域,成蟹養殖產量分別位列全國第一和第三,占比分別為46.3%、12.9%。本研究利用10對微衛星標記對偶數年的江蘇、安徽4個主要養殖群體的遺傳多樣性和遺傳結構進行分析,旨在為中華絨螯蟹的遺傳選育、種質資源保護與利用工作提供基礎資料。
1 材料與方法
1.1 樣品來源
2020年8月,中華絨螯蟹樣品采集于安徽省蕪湖市無為縣泉塘鎮、江蘇省南京市高淳區椏溪鎮、江蘇省宜興市楊巷鎮和江蘇省張家港市鳳凰鎮魏莊村,4個養殖群體的親本是當地養殖場于2018年底采集(或購買)的大規格野生長江蟹(雌蟹約 300 g,雄蟹約400 g)。每個養殖群體隨機選取30只成蟹樣品。
1.2 總基因組DNA提取
剪取上述中華絨螯蟹的附肢肌肉樣品,保存于無水乙醇中備用。提取河蟹總基因組DNA,提取方法參照基因組提取試劑盒[DP324,天根生化科技(北京)有限公司]說明書進行。提取的基因組DNA分裝后于-20 ℃冰箱保存備用。
1.3 微衛星位點的擴增
由表1可知,經預試驗,從參考文獻[7,9]中選取能夠穩定擴增、等位基因個數較多、多態性較好的10對微衛星引物,由生工生物工程(上海)股份有限公司合成熒光引物。

表1 中華絨螯蟹微衛星引物信息
PCR反應總體系20.0 μL:先加入超純水,依次補足模板DNA,10×Buffer緩沖液(TaKaRa) 2.0 μL,dNTP(2.5 mmol)1.5 μL,MgCl(2.5 mol/L)2.0 μL,上下游引物(5 μmol/L)各1.0 μL,DNA聚合酶0.3 U。PCR反應條件:96 ℃預變性3 min;94 ℃變性45 s,由表1可知,最佳退火溫度下保持45 s,72 ℃延伸1 min,32個循環;循環后72 ℃延伸10 min,4 ℃結束反應,-20 ℃保存備用。擴增產物經ABI-3730XL全自動DNA測序儀進行測定,然后使用軟件Genemarker 2.2讀取等位基因大小進行基因分型,內標為LIZ500。
1.4 數據分析
運用軟件CONVERT 1.31把基因型數據轉換為各種軟件格式。使用Popgene 1.32計算觀測等位基因的數目、有效等位基因數目、觀測雜合度、期望雜合度、近交系數及群體間Nei’s遺傳距離,采用MEGA 7.0軟件構建前述遺傳距離的UPGMA系統樹;用PIC_CALC 0.6小程序計算各位點及群體的多態信息含量。運用Arlequin 3.5軟件計算成對群體之間的遺傳分化系數(genetic differentiation coefficient,),并采用分子方差分析(analysis of molecular variance,AMOVA)統計變異來源。哈迪-溫伯格平衡檢驗通過Genepop在線軟件(http://genepop.curtin.edu.au/)進行。根據各位點等位基因頻率,應用Bottleneck軟件中的符號檢驗(Sign test)和符號秩次檢驗(Wilcoxon sign-rank),檢驗在3種突變模型(IAM、TPM和SMM)假設下是否偏離突變-漂移平衡(Mutation-drift equilibrium)。利用軟件Strcture 2.3.4對4個養殖群體進行遺傳聚類分析,確定最佳值,結果使用軟件CLUMPP打開,利用小程序DISTRUCT繪圖。
2 結果與分析
2.1 微衛星位點和群體遺傳多樣性
10對微衛星位點和4個養殖群體的遺傳信息比較見表2。有效等位基因數(:4.980~26.620)均>4,觀測雜合度(:0.585~0.990)、期望雜合度(:0.803~0.967)及多態信息含量(PIC:0.778~0.961)值均>0.5,符合群體遺傳評估的多樣度要求。
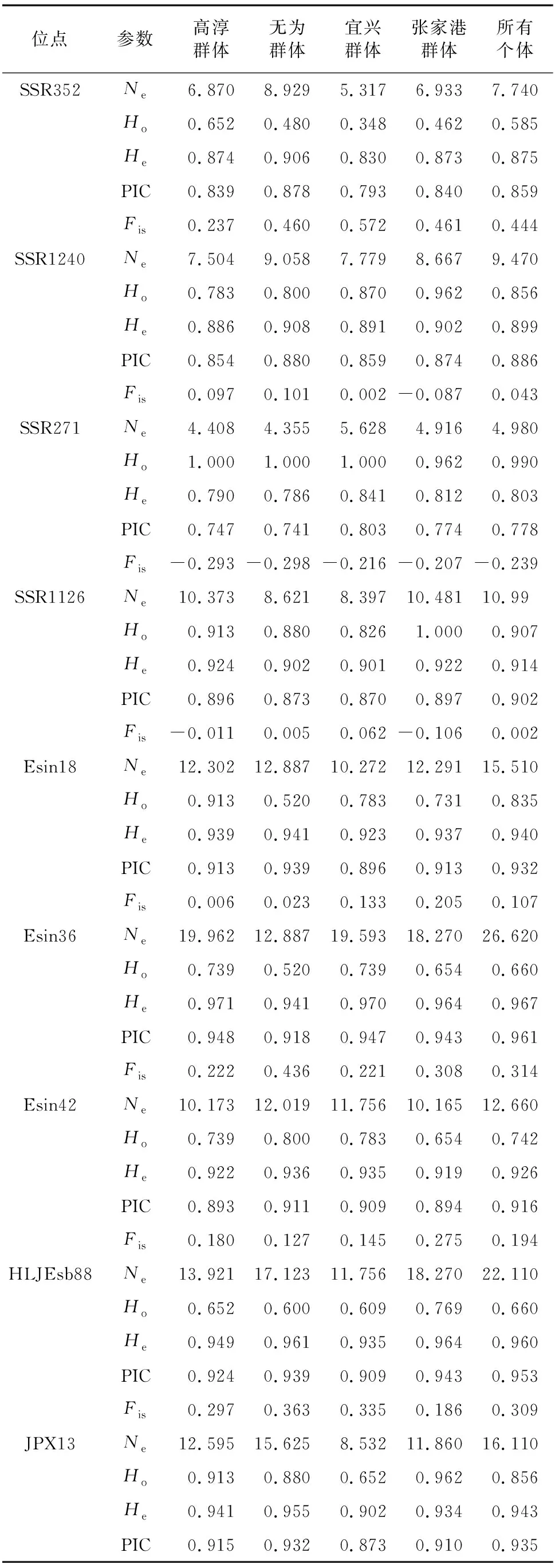
表2 微衛星位點的遺傳多樣性參數
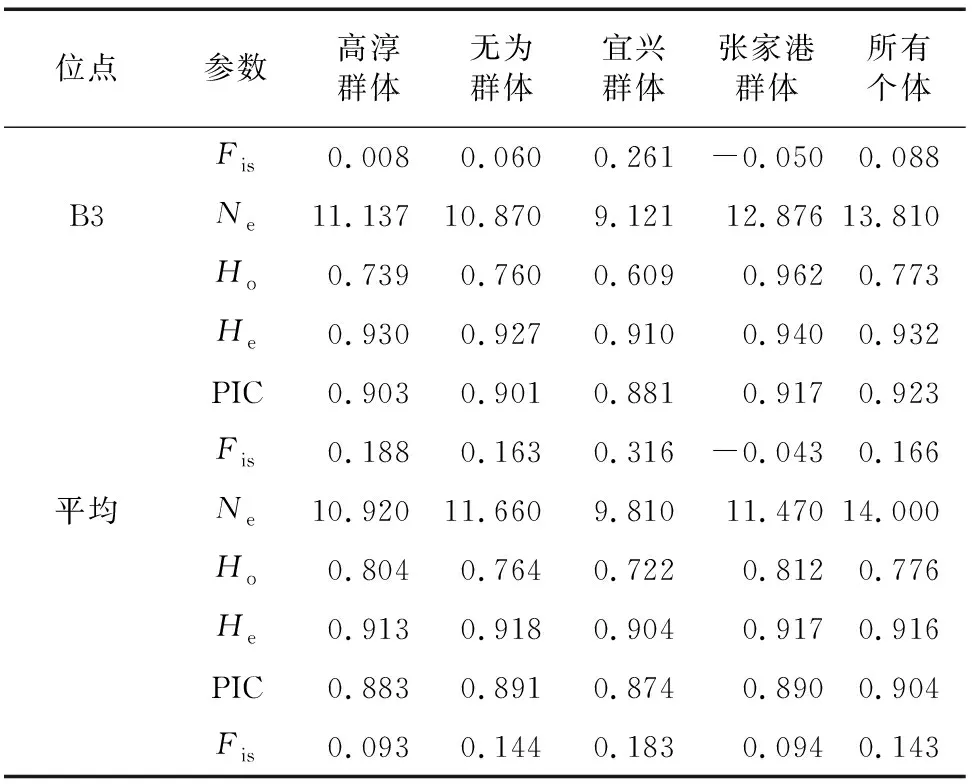
表2(續)
各養殖群體均顯示出較高的遺傳多樣性水平(=0.722~0.812,=0.904~0.918,PIC=0.874~0.891),無為泉塘群體期望雜合度及多態信息含量相對較高,宜興楊巷群體期望雜合度及多態信息含量相對較低。綜合各遺傳多樣性參數結果,認為各群體的遺傳多樣性大小順序為:無為群體>張家港群體>高淳群體>宜興群體。40個群體-位點的組合中,大部分組合(31個)近交系數()為正值且相應的<,所有群體的10個微衛星位點平均近交系數()值均為正。
2.2 群體遺傳距離和分化
對4個養殖群體間的成對遺傳距離和遺傳分化指數()進行分析。由表3、表4可知,遺傳距離較小(0.121 0~0.189 0),均低于0.3;群體間遺傳分化指數(0.001 5~0.008 2)較小,遠低于0.05,其中,遺傳分化程度最大的為無為群體和宜興群體(0.008 2),其遺傳距離也最遠(0.189 0)。AMOVA結果顯示, 群體之間的遺傳變異占總變異的比例僅為0.42%,而來自每個養殖群體內的遺傳變異占比高達99.58%,表明4個中華絨螯蟹養殖群體遺傳分化程度極低,無顯著遺傳分化。
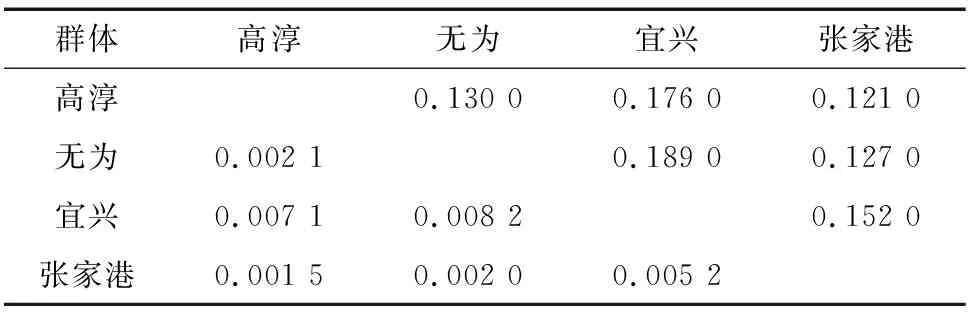
表3 中華絨螯蟹4個群體的遺傳距離(對角線上)和遺傳分化指數(對角線下)
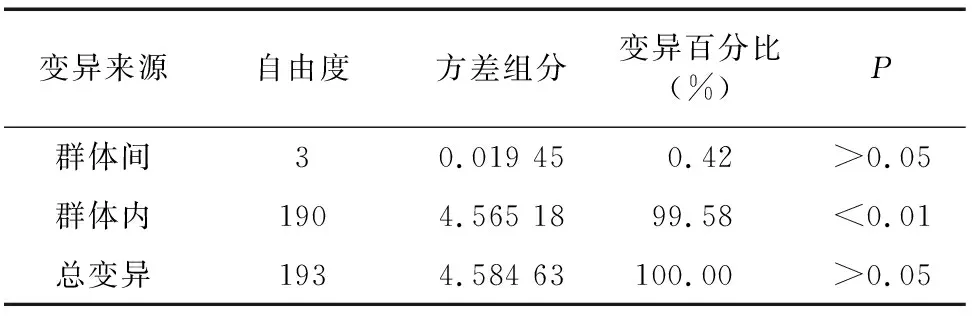
表4 中華絨螯蟹4個養殖群體的分子方差分析(AMOVA)
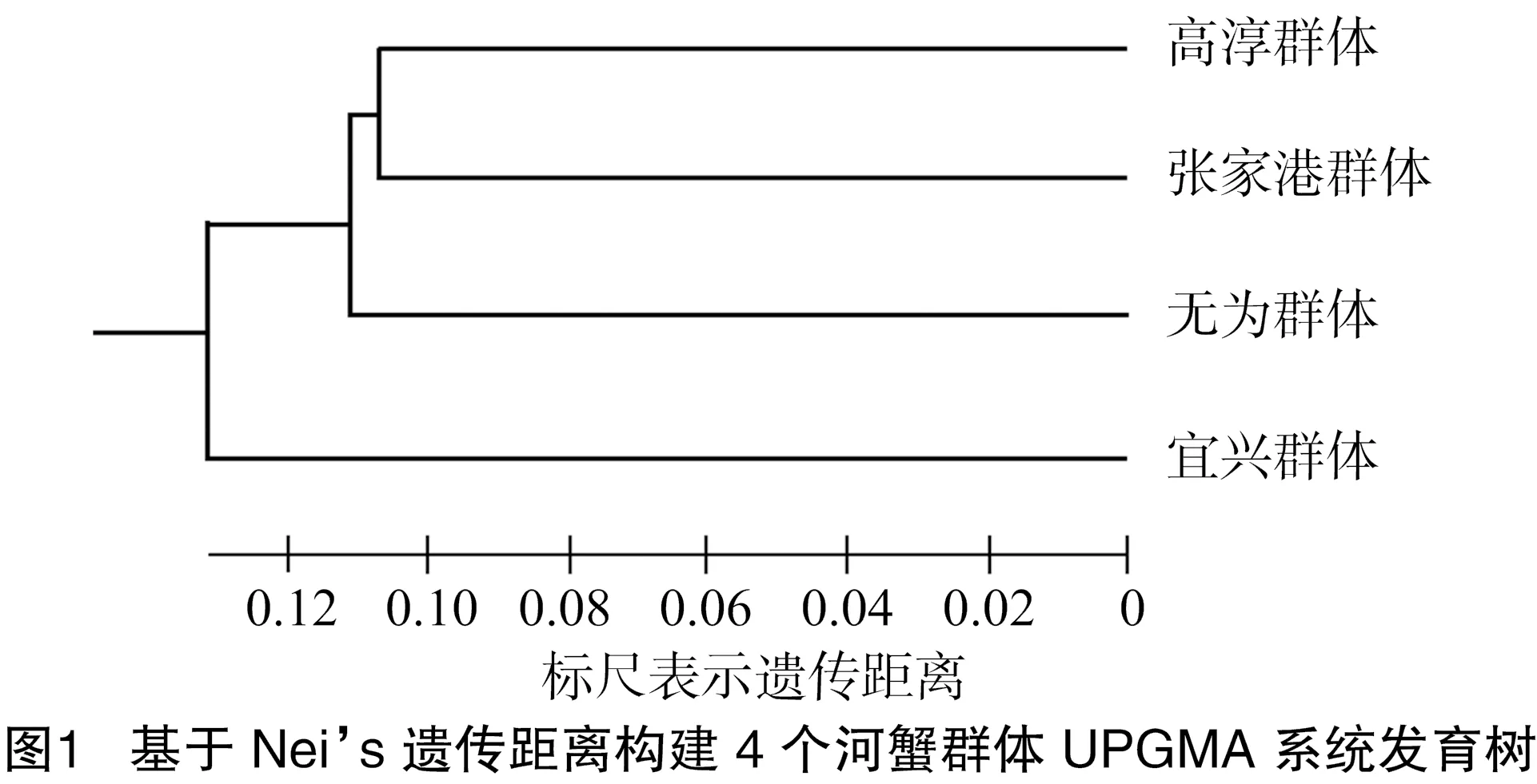
基于Nei’s遺傳距離用UPGMA法構建的4個群體的系統發育樹表明,4個養殖群體具有共同的祖先型,高淳和張家港群體最先聚類為一支,互為姊妹群,然后與無為群體聚類,最后與宜興群體聚類,表明高淳和張家港群體親緣關系最近、宜興群體與其他群體間親緣關系較遠(圖1)。
2.3 瓶頸效應分析
中華絨螯蟹4個養殖群體的突變-漂移平衡分析結果見表5。IAM假設下,Sign test顯示高淳和無為2個群體偏離了突變-漂移平衡,表現出雜合顯著過剩;Wilcoxon sign-rank顯示所有群體均偏離了突變-漂移平衡。SMM假設下,所有群體均符合突變-漂移平衡。TPM假設下,Sign test顯示所有群體均符合突變-漂移平衡;Wilcoxon sign-rank顯示僅無為群體偏離突變-漂移平衡,表現為雜合過剩。
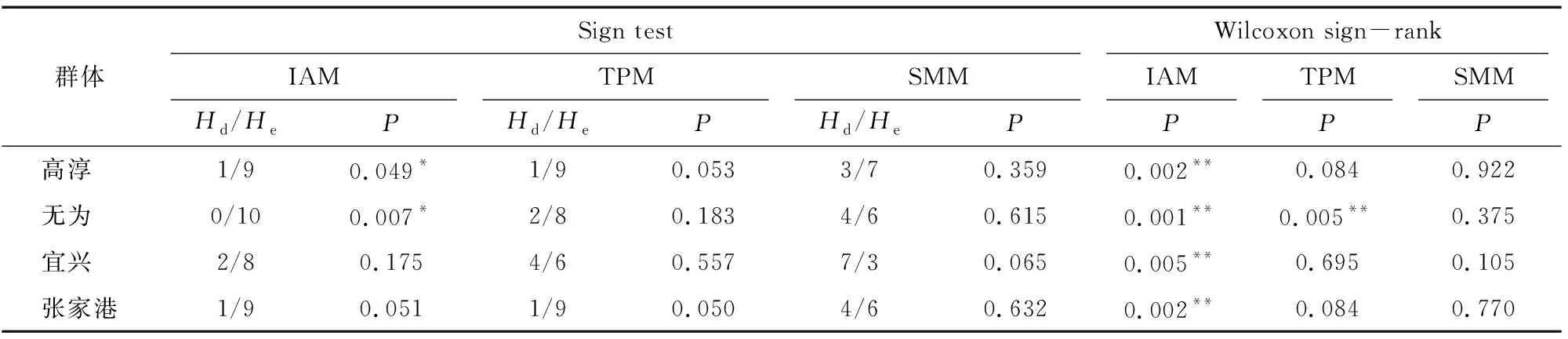
表5 中華絨螯蟹4個群體的突變-漂移平衡分析
2.4 群體遺傳結構分析
設置預設聚類組數為2~5,=4時為最佳聚類分組。遺傳結構圖見圖2,每個養殖群體均包含4個遺傳聚類組,說明4個養殖群體存在相似的遺傳組成,無明顯的遺傳結構差異。
3 討論與結論
3.1 中華絨螯蟹的遺傳多樣性
遺傳多樣性是種質鑒定的重要部分,一般認為遺傳多樣性的高低與物種或群體生存和進化潛力呈呈正相關,遺傳多樣性越高,物種或群體適應環境變化的能力越強。而有效等位基因數、預期雜合度和多態信息含量,均是衡量遺傳多樣性高低的重要參數。中華絨螯蟹4個養殖群體的為9.810~11.660,為0.904~0.918,為0.874~0.891,分別高于4.0、0.5、0.5的高度多樣性標準,均顯示出較高的遺傳多樣性水平。如,高于奇數年中華絨螯蟹3個人工選育群體(“長江1號”“光合1號”和七里海群體)和1個海河自然群體的遺傳多樣性(為4.79~5.87,為0.720~0.745,為0.687~0.716);高于偶數年長江水系中華絨螯蟹3個育種基礎群(崇明團結沙、揚中江段、靖江與六合、江浦江段)(:3.90~3.95,:0.696~0.705,:0.746~0.792);但與偶數年(2012年)遼河、黃河及長江3個水系的野生和養殖6個群體的遺傳多樣性結果(:13.08~14.10,:0.870~0.881,均值0.864)比較,較小、略小、值相近;大小還與長江水系野生和人工繁殖大眼幼體群體的遺傳多樣性水平(0.898、0.893)相近。另外,基于線粒體等其他分子標記的中華絨螯蟹研究也顯示多數野生和養殖群體均具有較高的遺傳多樣性。綜上所述,中華絨螯蟹具有高遺傳多樣性應該是普遍現象;本研究的中華絨螯蟹4個養殖群體遺傳多樣性處于高水平,具有較大的育種潛力。

3.2 中華絨螯蟹的遺傳結構
群體遺傳距離和遺傳分化指數分別是用于衡量群體間親緣關系、遺傳分化程度的重要指標。本研究中,中華絨螯蟹成對群體間的遺傳距離范圍為0.121 0~0.189 0、遠低于0.3的物種標準,表明這些群體親緣關系較近。成對群體間值為0.001 5~0.008 2,均遠低于0.05的遺傳分化程度中等以上標準。進一步的分子變異分析(AMOVA)結果顯示,群體間遺傳變異占比極小(0.42%)。Structure遺傳聚類分析結構顯示,所有個體可分為4個遺傳組,但每個養殖群體中均包含這4個遺傳組,這些一致表明4個中華絨螯蟹養殖群體間遺傳組成相似,群體間無顯著遺傳分化。
近交系數大小常用于評估群體內個體間的近交程度。本研究中,10個微衛星位點在每個群體及所有個體中的雖然均為正值(0.093~0.183、0.143)但數值較低,且40個群體-位點組合中的9個值為負值,這些結果表明,中華絨螯蟹4個養殖群體不僅均存在一定程度的近交,而且還存在少量的遠緣繁殖。考慮到這些群體高度的遺傳多樣性水平,分析可能其苗種或親本群體來源較為復雜。事實上,已有不少研究報道,中華絨螯蟹在我國不同地區相互引種,導致不同水系的中華絨螯蟹種質相互混雜,致使養殖群體遺傳多樣性偏高。
基于Nei’s遺傳距離構建的4個河蟹群體UPGMA系統發育樹表明,4個養殖群體具有共同的祖先型,高淳和張家港群體親緣關系最近,其次是無為群體,最后是宜興群體,這表明宜興群體與其他群體間遺傳距離較遠。
瓶頸效應分析結果顯示,3種突變模型(IAM、TPM、SMM)假設下的4個中華絨螯蟹養殖群體的突變-漂移平衡評估結果不同。由于現有研究認為微衛星數據更符合TPM模型,Wilcoxon符號秩次檢驗比符號檢驗的統計效率相對較高,并可用于4個位點以上任意樣本數群體的分析。因此,在TPM假設下,符號秩次檢驗結果顯示僅無為群體偏離了突變-漂移平衡,且表現為雜合顯著過剩,這與其遺傳多樣性最高的結論相吻合。因此,雖然無為群體經歷了瓶頸效應,但由于有遠緣繁殖的引種效果,其遺傳多樣性依然很高。
綜上所述,中華絨螯蟹4個養殖群體的遺傳多樣性水平高,群體間無顯著遺傳分化,群體間可能存在少量的種質混雜。從開發利用的角度來說,在今后的選育中應該結合采用分子遺傳學、形態學等多種方法,將傳統選育技術與分子輔助育種技術相結合,對長江水系中華絨螯蟹進行提純復壯,培育長江水系中華絨螯蟹良種。
