哈茨木霉對鎘脅迫下草地早熟禾的促生和增抗效應及其生理機制
2022-09-08 01:31:08董斯琳姚桐桐謝琳淼孫廣玉敖紅
浙江農業科學 2022年9期
關鍵詞:植物
董斯琳,姚桐桐,謝琳淼,孫廣玉,敖紅
(東北林業大學 生命科學學院,黑龍江 哈爾濱 150040)
重金屬鎘(Cd)的生物毒性很強,是植物生長發育的非必需元素[1]。近年來,工業和農業生產中將鎘不同程度地排放到土壤環境中,使鎘污染成為危害最大、分布最廣的農業污染之一[2]。據統計,中國已有2 000萬hm2的土地面積被鎘污染,并且污染面積正逐步擴大[3]。過量鎘會對動植物產生極大危害,植物體內的鎘過量時會產生毒害癥狀,主要表現為抑制根部增長,破壞某些生理生化反應,進而遏制根部從土壤吸收水分和營養物質[4]。植物的生長發育過程同樣會受到鎘的抑制,其結果會導致植物的生長遲緩,植株萎蔫,甚至導致死亡[5]。更重要的是,鎘可以通過食物鏈進入人體,對人體造成巨大損傷[6]。對重金屬土壤的修復有多種方式,其中植物修復受到人們的關注。而草坪草因其生命力旺盛、覆蓋力和適應性強、分布廣、抗逆性強、易收割等優點在植物修復重金屬污染土壤中具有較好的應用價值[7]。
木霉菌是一類被廣泛應用的植物生防菌,能抵御多種植物病原菌[8]。哈茨木霉菌可以防治及緩解黃瓜的根腐病和線蟲病[9]、馬鈴薯黑痣病[10]、煙草青枯病[11]等植物病害。木霉菌還具有促進植物的生長發育,改善植物生理代謝等作用[12-15]。近年來發現,利用木霉菌與植物的互作關系可以增強植物對微生物脅迫的抗性[16]。李舒依等[17]研究發現,使用哈茨木霉菌可以提高鹽脅迫下黃瓜種子的發芽率和胚芽長;付健等[18]研究發現,施用哈茨木霉菌可以提高鹽堿土壤中玉米葉片光合色素含量,增強光合作用、抗氧化酶活性,從而緩解玉米鹽堿抑制。關于哈茨木霉菌對植物重金屬抗性作用相關研究較少,待進一步探索。
草地早熟禾(Poapratensis)是一種多年生草坪草,其生命力十分頑強,能適應各種不同的生長環境,無論潮濕還是干旱;氣溫偏高或是寒冷地區,只要水分供應充足就能生長[19]。因此,草地早熟禾常作為草坪草廣泛運用在園林綠化中。本試驗使用草地早熟禾作為材料,研究哈茨木霉菌對不同濃度鎘處理下草地早熟禾種子萌發、生長狀況及生理效應的影響,探討哈茨木霉菌提高植物抗鎘能力的生理機制,并為植物修復鎘污染土地的應用提供理論支撐。
1 材料和方法
1.1 材料
草地早熟禾(Poapratensis),購于哈爾濱市和平綠化有限公司。哈茨木霉菌(T.harzianum)Th62菌株由東北林業大學園林學院張榮沭博士提供。Cd脅迫處理使用氯化鎘試劑。
1.2 方法
1.2.1 種子萌發試驗
哈茨木霉濃度篩選。選取顆粒飽滿、大小均勻的草地早熟禾種子先用95%乙醇浸泡2 min,再用20%次氯酸鈉溶液消毒20 min,蒸餾水沖洗3~4遍后浸種2 h。取5個小燒杯放入足夠種子,將濃度分別為0(CK)、102、104、106和108mL-1的哈茨木霉菌懸浮液2 mL依次加入燒杯,浸種3 h。隨后將種子轉移到具有雙層濾紙發芽床的直徑9 cm的培養皿中,每皿50粒種子,加蒸餾水3 mL,設置3個重復。光照14 h、25 ℃,黑暗10 h、15 ℃。28 d為一個發芽期,每天記錄種子萌發數量。本試驗發芽結束時間以5 d內沒有新種子發芽為準。結果顯示,106mL-1處理后種子萌發率最高(表1),因此,選該濃度作為后續種子萌發試驗的哈茨木霉處理濃度。

表1 不同濃度哈茨木霉菌處理對草地早熟禾的種子萌發率的影響
用106mL-1哈茨木霉菌懸浮液(T處理)和蒸餾水(C)分別浸種,處理方法同上。處理后的種子移入具雙層濾紙發芽床的培養皿,皿中分別加入濃度為0(CK)、100、200、300和400 μmol·L-1的CdCl2溶液,每個處理3次重復,每個重復50粒種子。每日記錄發芽及生長情況,10 d計算發芽勢、28 d計算發芽率,并測量胚芽和胚根長度,計算出發芽指數及活力指數。
發芽勢=(10 d內發芽種子數/供試種子數)×100%;
發芽率=(24 d內發芽種子數/供試種子數)×100%;
發芽指數=Σ(在不同時間的發芽數/發芽日數);
活力指數=發芽指數×胚芽長度。
1.2.2 幼苗生長、鎘積累及生理特性試驗
幼苗盆栽試驗:將配置好的濃度為108mL-1(根據預試驗結果確定)的哈茨木霉提取液混入無菌土中,每500 g無菌土加入200 mL哈茨木霉菌提取液,將草地早熟禾種子播種于土中(T處理),同時設置土壤中加200 mL蒸餾水為對照組(C)。28 d后補加一次哈茨木霉菌提取液,再培養28 d后進行Cd2+處理。Cd2+濃度為0(CK)、10、40、80和100 mg·L-1,每處理3個重復,每盆100 mL Cd溶液。處理7和14 d后分別取樣,測定草地早熟禾幼苗的株高、生物量、各項生理指標和鎘含量。脯氨酸含量參照高俊鳳的測定方法;可溶性蛋白含量的測定參照G250法;丙二醛含量的測定參照硫代巴比妥酸法;超氧化物歧化酶(SOD)的測定采用氮藍四唑法;過氧化物酶(POD)的測定采用愈創木酚法;過氧化氫(CAT)的測定采用過氧化氫法;鎘含量的測定采用原子吸收分光光度計進行測定。
1.3 數據處理與分析
用Excel 2019整理試驗數據、繪制圖表。
2 結果與分析
2.1 哈茨木霉對鎘脅迫下草地早熟禾種子萌發和幼苗生長的影響
由表2可知,隨著Cd2+濃度增大,種子發芽率、發芽勢降低,且發芽勢受到抑制程度大于發芽率。哈茨木霉菌浸種(T)組相對于未浸種(C)組,發芽率和發芽勢、發芽指數和活力指數在各濃度Cd2+處理下均升高。
由圖1可知,隨著Cd2+濃度的增加,草地早熟禾種子的胚芽長、胚根長均顯著降低,且T組相對于C組,種子的胚芽長在各Cd2+處理下均升高,胚根長度除400 μmol·L-1濃度差異不明顯外,其他組同樣升高。
2.2 哈茨木霉對鎘處理下草地早熟禾植株生長的影響
由表3可知,隨著Cd2+濃度的升高,幼苗的株高呈降低趨勢,且T組相對于C組,株高在不同濃度Cd2+處理下均升高。Cd2+處理7 d,各Cd2+濃度處理下T組株高比C組分別增長了5.29、7.87、9.22、8.14和8.41 cm;Cd2+處理14 d,T組比C組分別增長了5.84、8.28、8.79、8.75和7.32 cm。
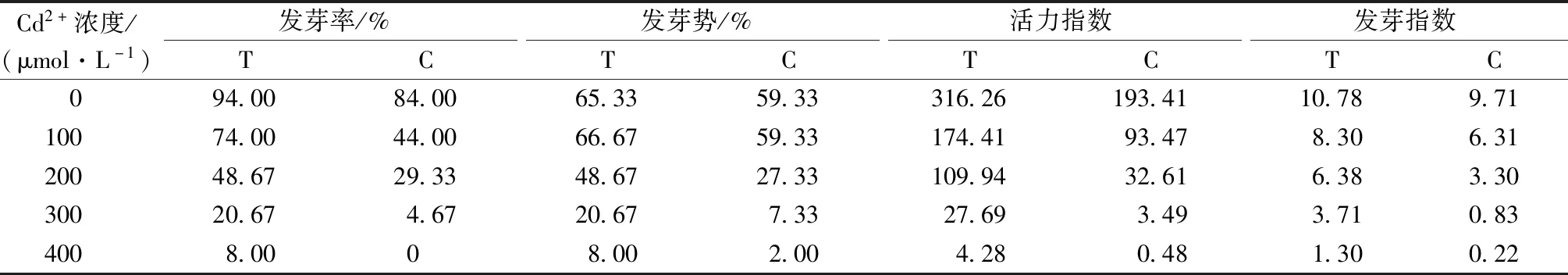
表2 哈茨木霉對Cd2+脅迫下草地早熟禾種子萌發的影響
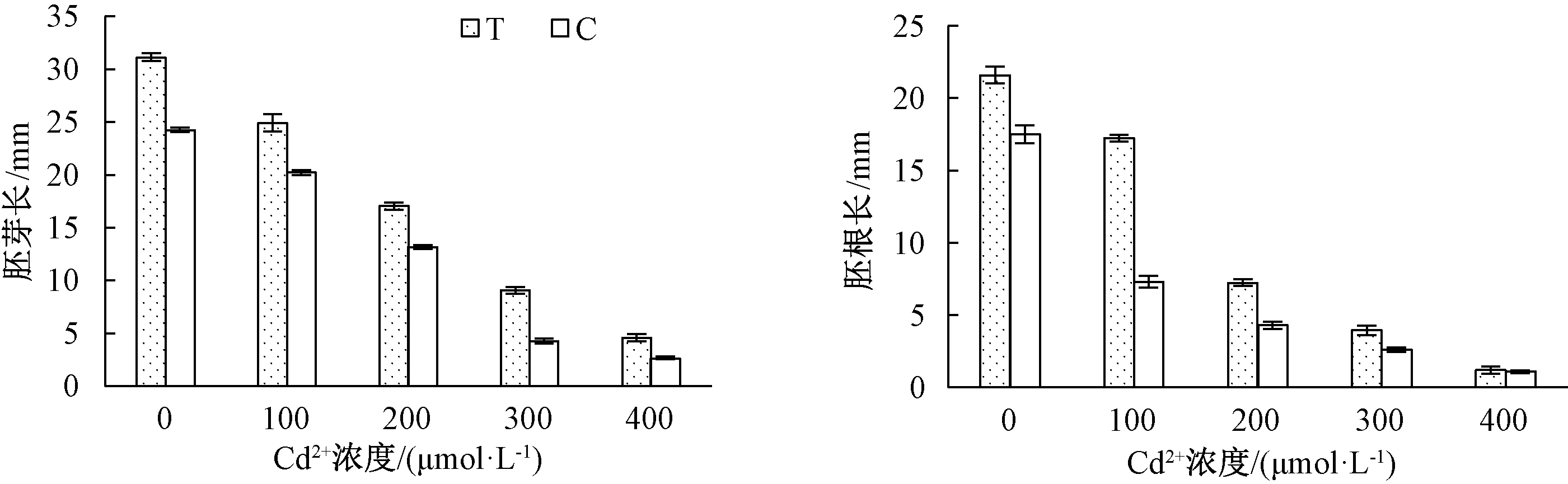
柱上無相同小寫字母表示組間差異顯著(P<0.05)。圖2~4同。圖1 哈茨木霉對鎘脅迫草地早熟禾胚芽、胚根長度的影響
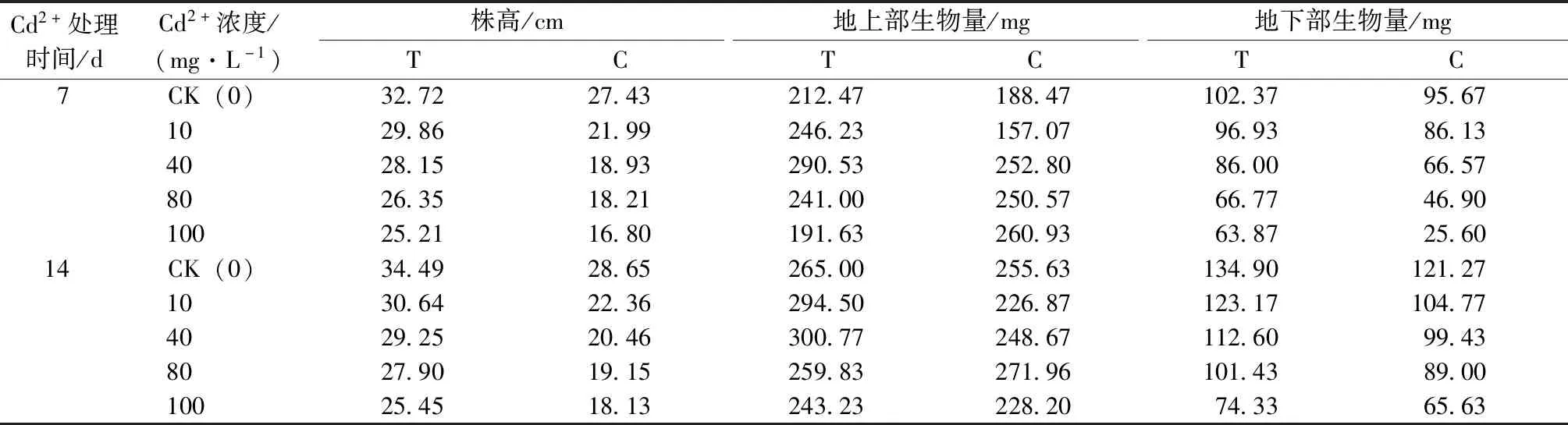
表3 哈茨木霉菌對Cd2+脅迫下草地早熟禾株高及生長量的影響
幼苗地上部分生物量的變化規律總體看,一定濃度的Cd2+對其有促進作用,且在低濃度下,T組數值高于C組。地下部分生物量隨Cd2+濃度升高而降低。各Cd2+處理下T組比C組分別增長了6.7、10.8、19.4、19.9和38.3 mg;處理14 d時,各Cd2+處理下T組比C組分別增長了13.6、18.4、13.2、12.4和8.7 mg。
2.3 哈茨木霉菌對鎘處理下草地早熟禾生理特性的影響
由表4可知,隨著Cd2+濃度的升高,葉綠素a、葉綠素b和總葉綠素含量均呈降低趨勢。Cd2+處理7 d,各Cd2+濃度處理下T組葉綠素a比C組分別增加了0.04、0.07、0.11、0.19和0.20 mg·g-1;葉綠素b含量增加了0.11、0.31、0.34、0.32和0.33 mg·g-1;葉綠素總含量分別增加了0.15、0.39、0.44、0.50和0.53 mg·g-1。Cd2+處理14 d,各濃度Cd2+處理下,T組相較于C組葉綠素a含量增加,但葉綠素b和總葉綠素含量減少。
脯氨酸和可溶性蛋白是植物在逆境下產生的主要滲透調節物質。由圖2可知,隨著Cd2+濃度的升高,脯氨酸含量增加,且T組脯氨酸含量高于C組。Cd2+處理7 d,各Cd2+濃度處理下T組脯氨酸含量比C組分別增加了79.91、104.37、119.95、115.71和119.87 μg·g-1;Cd2+處理 14 d,T組較C組分別增加了47.15、39.00、41.39、89.02和92.54 μg·g-1。
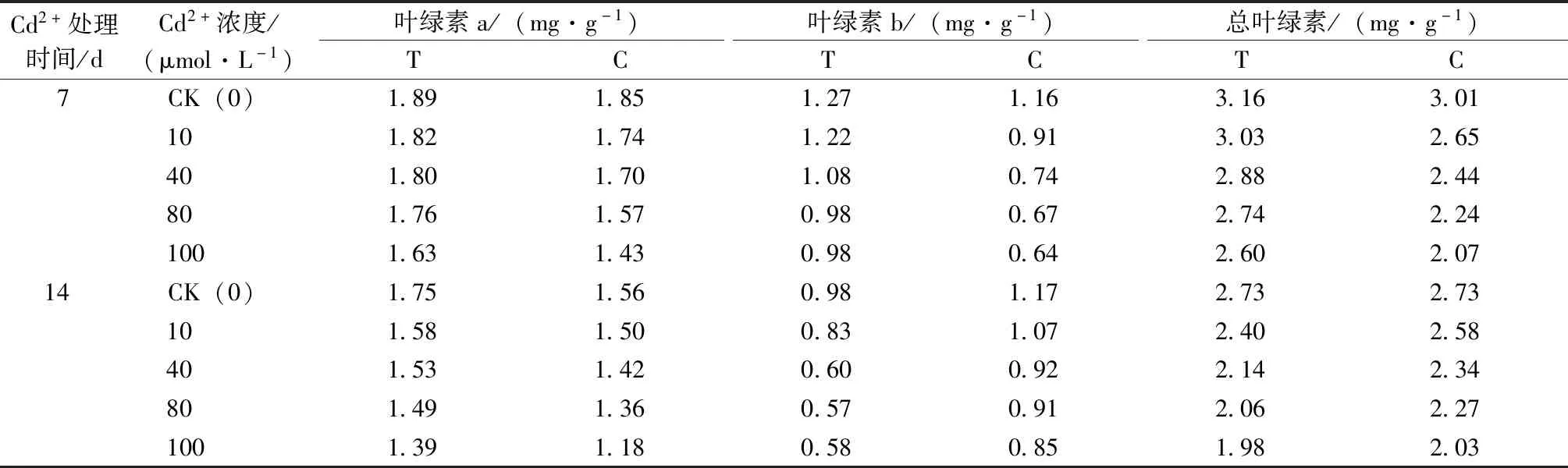
表4 哈茨木霉菌對鎘脅迫下草地早熟禾葉綠素含量的影響
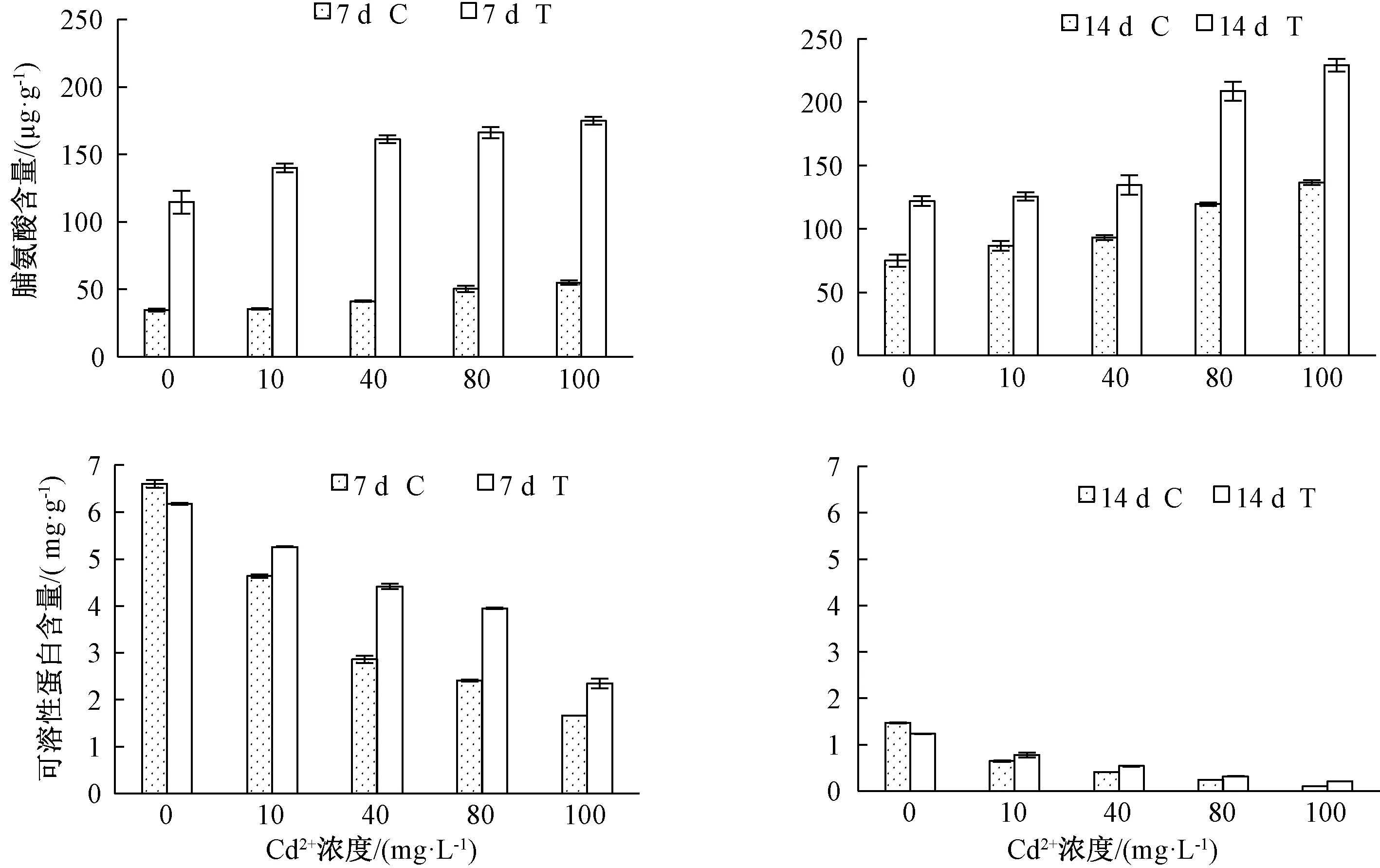
圖2 哈茨木霉對鎘脅迫下草地早熟禾脯氨酸、可溶性蛋白含量的影響
隨著Cd2+濃度的升高,可溶性蛋白含量減少。在無Cd2+處理時,T組低于C組,在各組Cd2+處理時,T組高于C組。Cd2+處理7 d,各Cd2+濃度處理下T組可溶性蛋白含量比C組分別增加了0.62、1.56、1.54和0.69 mg·g-1;Cd2+處理14 d,T組較C組分別增加了0.13、0.12、0.08和0.11 mg·g-1。
丙二醛(MDA)為膜脂過氧化產生的有毒物質。圖3結果顯示,Cd2+濃度越大,葉片MDA含量越大,且T組均低于C組。Cd2+處理7 d,各Cd2+濃度處理下T組MDA含量比C組分別減少了0.45、0.11、0.43、0.65和0.83 mg·g-1;Cd2+處理14 d,T組較C組分別減少了0.81、0.90、0.64、1.58和1.43 mg·g-1。
在逆境下,植物體內可產生大量的活性氧,而保護酶系統可以起到清除活性氧的作用。由圖4可知,隨著Cd2+濃度的增加,SOD和CAT活性先升高后降低,POD活性呈逐漸升高。在Cd2+處理7 d后,除Cd2+濃度為80 mg·L-1外,其他各Cd2+濃度下T組SOD活性較C組升高;Cd2+處理14 d后,比較T與C處理組SOD活性在低Cd2+濃度下升高,高Cd2+濃度下差異不明顯。Cd2+處理7 d和14 d后,在各Cd2+濃度下,T組POD活性較C組POD活性均降低。Cd2+處理7 d后,40 mg·L-1Cd2+處理下CAT活性最大,且Cd2+濃度為80和100 mg·L-1時,T組較C組分別減少1.67、3.93 U·g-1;Cd2+處理14 d,T組較C組CAT活性增加。
2.4 哈茨木霉菌對鎘處理下草地早熟禾鎘轉運的影響
由表5可知,草地早熟禾地上部所吸收的Cd含量隨著Cd2+處理濃度的增加和處理時間的延長而增加,且T組含量均高于C組。Cd2+處理7 d,各Cd2+濃度處理下T組Cd含量比C組分別增加了1.24、0.78、2.71、2.10和4.26 mg·kg-1;Cd2+處理14 d,除40 mg·L-1Cd2+處理T組與C組無明顯差異外,其他組處理下T組比C組分別增加了0.32、1.62、1.37和2.46 mg·kg-1。
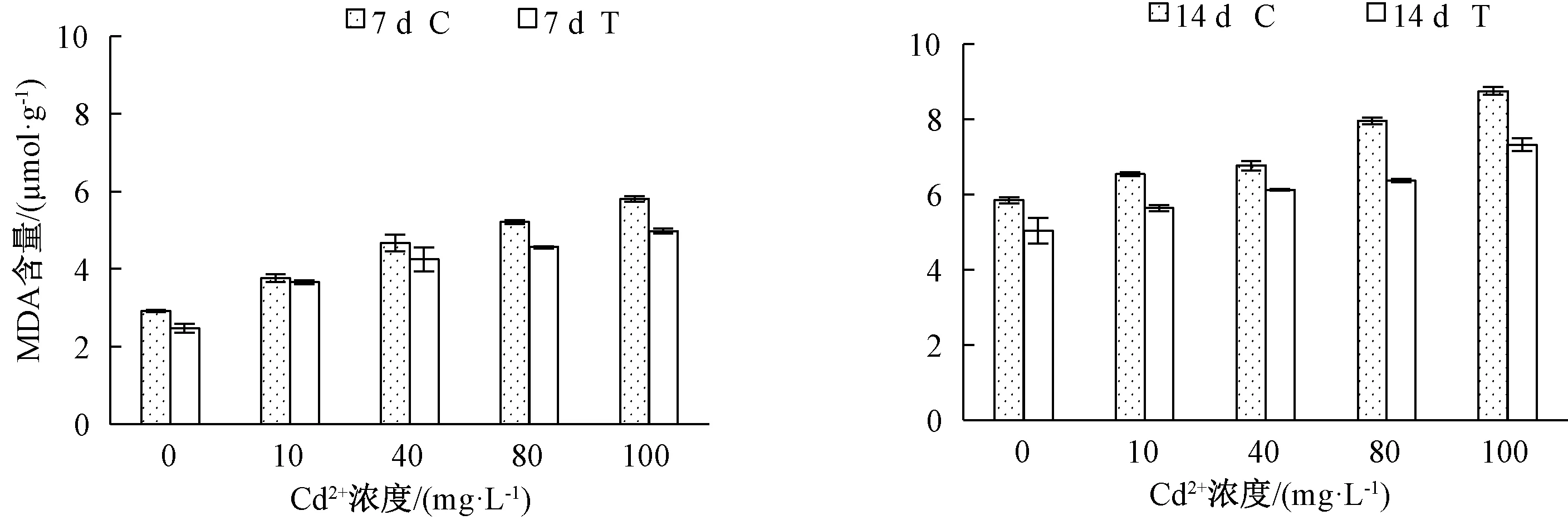
圖3 哈茨木霉菌對鎘脅迫下草地早熟禾MDA含量的影響
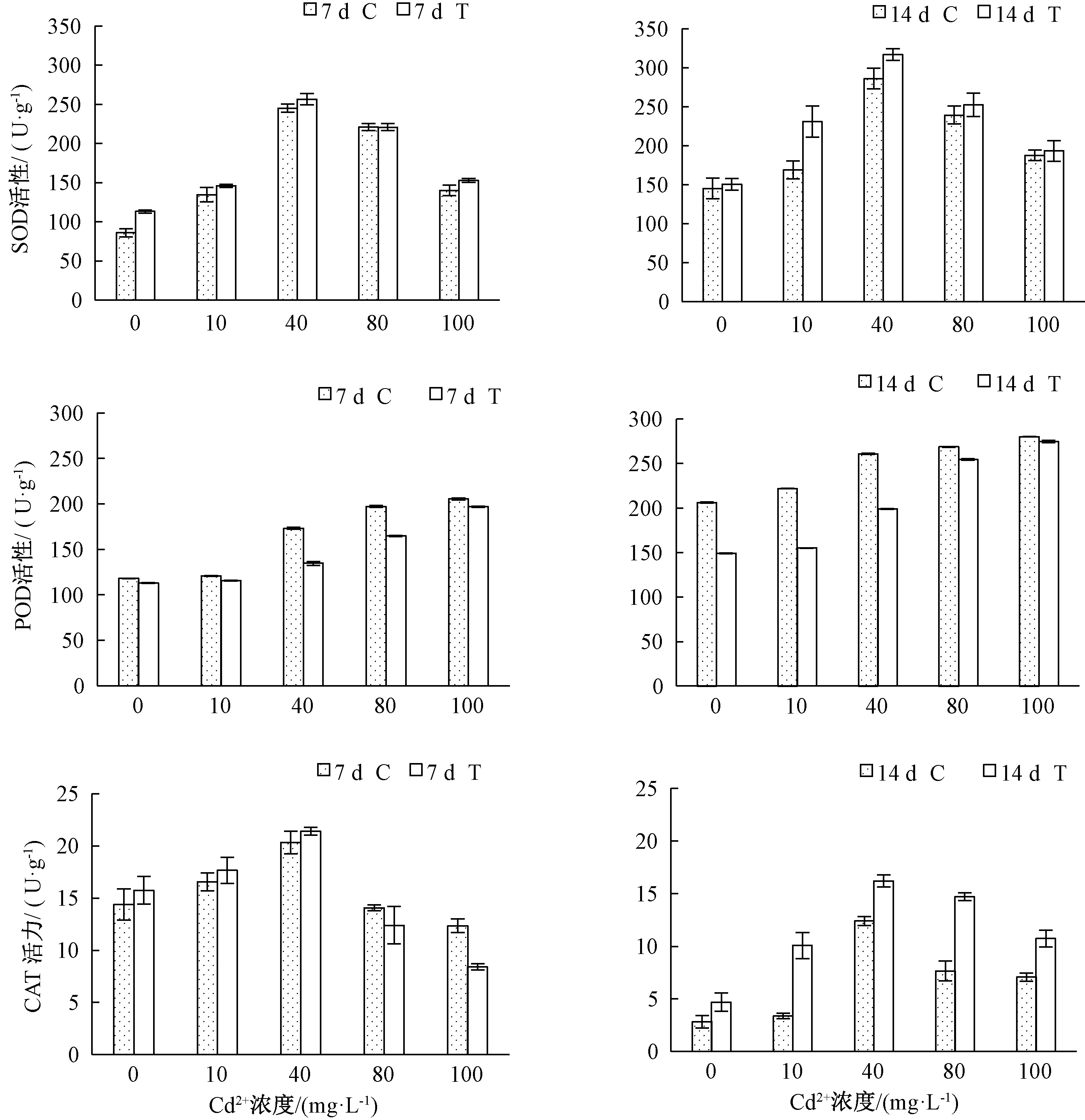
圖4 哈茨木霉菌對鎘脅迫下草地早熟禾抗氧化系統活性的影響
C組草地早熟禾地下部所吸收的Cd含量隨著Cd2+處理濃度的增加和處理時間的延長而增加;T組地下部所吸收的Cd含量隨著Cd2+處理濃度的增加先增加后減少;隨處理時間的延長而增加。100 mg·L-1Cd2+處理時,T組地下部Cd含量低于C組。鎘脅迫會降低早熟禾Cd轉移系數,施加哈茨木霉菌會增加早熟禾Cd轉移系數。處理7 d時,0、10、40、80和100 mg·L-1Cd2+處理時T組比C組增加了0.25、0.04、0.09、0.02和0.09;處理14 d時,各組Cd2+處理時T組比C組增加了0.19、0.05、0.01、0.01和0.05。
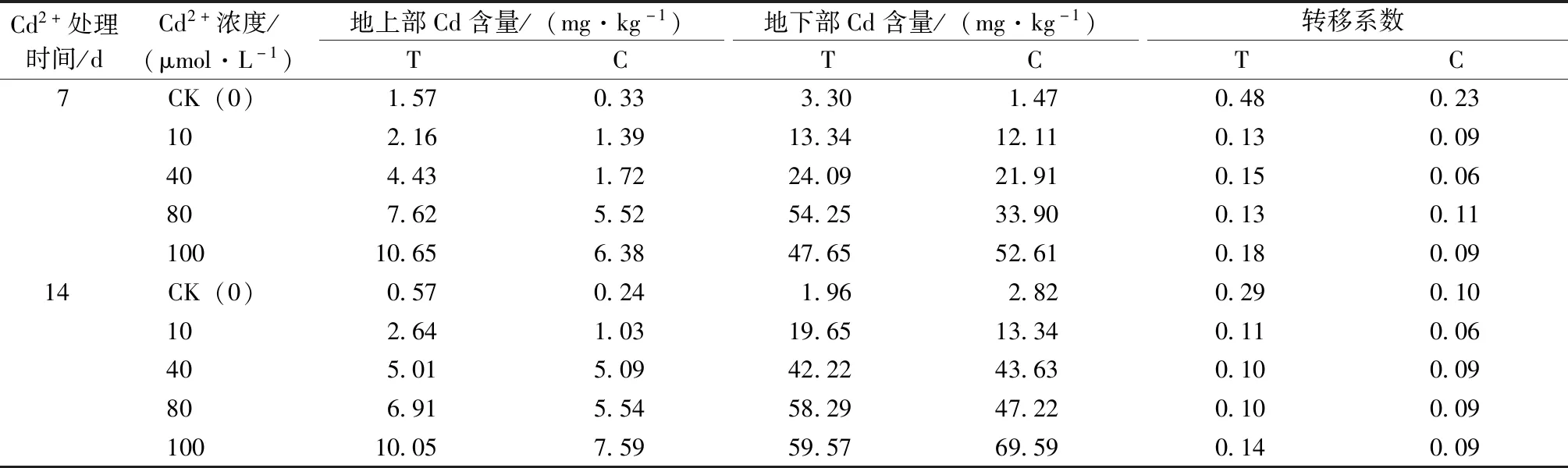
表5 哈茨木霉對鎘脅迫下草地早熟禾Cd含量的影響
3 小結與討論
哈茨木霉作為一種重要的生防菌,對多種植物具有促生效果[20]。眾多研究結果顯示,經低濃度哈茨木霉處理后種子的發芽率有不同程度的提高,濃度升高時促進作用變弱[21-22]。木霉菌可以產生一些生物調節類物質,如赤霉素、生長激素等,赤霉素可以增加種子萌發過程中可溶性糖和可溶性蛋白的含量,從而促進生長[23]。有研究還發現,使用康寧霉素浸種可減少種子內部營養物質外滲,保留種子內部營養充足,供種子萌發過程使用[24]。本試驗結果表明,濃度為106mL-1的哈茨木霉菌對促進草地早熟禾種子的萌發和生長最為有效,濃度超過106mL-1對發芽率的促進作用減弱,這與前人研究結果一致。在鎘脅迫下,隨著Cd2+濃度的升高,草地早熟禾種子的發芽率呈顯著降低的趨勢,且T組較C組的發芽率、發芽勢、發芽指數、活力指數均增加,說明哈茨木霉菌浸種可以在一定程度上減輕Cd抑制的種子萌發過程。試驗還表明,草地早熟禾種子的胚芽、胚根受鎘脅迫的抑制,隨著Cd2+濃度的升高抑制作用增強,且對胚根的抑制作用大于胚芽。而哈茨木霉菌可以起到緩解鎘脅迫對草地早熟禾的毒害作用。
鎘對植物形態和生理代謝具有嚴重的毒害作用[25]。諸多研究表明,鎘對植物的影響呈“低促高抑”的現象,低濃度可以促進植物的生長,但當鎘含量過高時,會抑制植物生長,嚴重時還會出現植株矮小萎黃、根系死亡、生物量減少等癥狀[26-27]。本研究結果顯示,隨著鎘脅迫強度及脅迫時間增加,草地早熟禾植株株高呈下降趨勢,相較于C組,T組可以緩解鎘脅迫的抑制作用,使株高增加。鎘脅迫對草地早熟禾植株地上生物量、地下生物量的影響有差異,T組在低濃度Cd2+處理時地上生物量高于C組。地下部分生物量隨著Cd2+濃度增加而降低,且T組較C組增高。從上述結果可以看出,哈茨木霉菌對鎘脅迫下草地早熟禾的生長有一定的促進作用。
光合作用是植物轉化光能為化學能的基礎生理活動,葉綠素具有捕捉和傳遞光能的作用[28],是光合作用的關鍵色素,其含量多少在一定程度上標志著植物生長能力的強弱[29]。本研究結果顯示,處理7 d后,隨著Cd2+濃度升高草地早熟禾植株的葉綠素各指標降低,T組含量高于C組;隨著處理時間增長到14 d時,T組的葉綠素b含量降低并低于C組,說明哈茨木霉菌可以緩解鎘脅迫短時間內造成的草地早熟禾植株的光合色素的下降。
細胞滲透調節物質是一類具有維持細胞平衡作用的物質,主要包括游離脯氨酸、可溶性蛋白、可溶性糖等[30]。逆境條件會使植物中脯氨酸含量顯著增加,當逆境持續加重時,會致使植株內脯氨酸含量減少,嚴重時還會致使植株死亡,所以通過植物體內游離脯氨酸含量的變化情況觀察植物的抗逆性強弱[31]。可溶性蛋白的積累可以防止植物細胞水分的流失,進而增強植物抗逆性;可溶性蛋白還可以充當各種生理代謝過程所需的酶類,植物體在逆境條件下,也會消耗大量的可溶性蛋白來激活抗氧化系統,使植物抵御傷害[32]。本試驗顯示,草地早熟禾植株脯氨酸含量隨著Cd2+濃度的增加和脅迫時間的增長而增加,且T組高于C組;可溶性蛋白含量則隨Cd2+濃度增加和脅迫時間增長而減少,且T組同樣高于C組。說明哈茨木霉菌可以增強鎘脅迫下草地早熟禾的滲透調節能力。
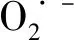
植物對鎘的吸收能力是有限度的,當外界鎘濃度超過植物本身能承受的濃度時,植物將減少對鎘的吸收[37]。卜和申等[38]發現,鎘污染土壤中分離的哈茨木霉菌可以通過細胞壁和細胞質吸附Cd2+,產生IAA類物質,促進植物生長,并可提高植物吸收Cd2+的能力,使Cd2+向地上部分轉移,從而降低土壤中的鎘含量。本試驗結果顯示,草地早熟禾地上部分Cd2+含量隨著Cd2+濃度的增加而增加,且處理時間越長積累的越多,T組積累量比C組增多。地下部分Cd2+含量隨著Cd2+濃度增加大致呈增加趨勢,且0~80 mg·L-1時,T組Cd含量較C組增高。轉移系數可以反映植物根部從土壤中吸收重金屬后從地下向上轉運能力的強弱,轉移系數越大,轉運能力越強[39]。施加哈茨木霉菌可以增加Cd轉移系數,處理7 d時,Cd2+濃度為40 mg·L-1時轉移系數增長最多,處理14 d時,Cd2+濃度為10 mg·L-1時轉移系數增長最多。這說明哈茨木霉在較低Cd2+濃度范圍內可以增強草地早熟禾對鎘的積累和轉運,并且通過多種生理效應保證植株正常生長,這對于增強鎘污染土壤的植物修復效果具有重要的意義。
本試驗結果表明,哈茨木霉浸種能夠減輕鎘脅迫對草地早熟禾種子萌發及幼苗生長的抑制作用;哈茨木霉也可以通過提高Cd轉運能力,減少Cd對植株根系的直接傷害,緩解鎘脅迫對草地早熟禾生長的抑制作用,并可以通過降低葉綠素破壞、減少MDA積累,增強抗氧化酶(SOD、POD、CAT)活性,增加滲透調節物質來提高草地早熟禾對Cd的抗性。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
