兩類紫云英苗菜生長過程中酚類物質及其抗氧化活性的變化
2022-09-13 10:33:18趙齊燕唐寧賈鑫程永強
食品工業科技 2022年18期
趙齊燕,唐寧,賈鑫,程永強
(植物源功能食品北京市重點實驗室,中國農業大學食品科學與營養工程學院,北京 100083)
紫云英(L.)是豆科黃耆屬二年生草本植物,紫云英多作為綠肥、蜜源植物和牧草栽培使用,但種植效益不高。紫云英較其他綠肥含有更多的羧酸和類蛋白質成分,其花蜜總抗氧化性高于金合歡等花源蜂蜜,嫩梢富含抗壞血酸,且有毒有害物質含量均符合國家標準。可見,紫云英在安全、營養兩方面都具備成為綠色蔬菜品種的潛力。2014 年,何春梅等開發了菜肥兼用的紫云英高效種植施肥技術。2020 年初,浙江省嘗試綜合開發紫云英的綠肥、飼料、菜用、景觀、蜜用功能,富陽、臨安、奉化等地部分農戶把紫云英作菜用種植,經濟效益非常可觀。在此基礎上,季衛英等篩選出了適合種植在浙江地區的菜用紫云英品種。由此可見,菜用紫云英的開發利用、發掘紫云英的經濟效益是大勢所趨。
芽苗菜,也稱活體蔬菜,生育期短、品質脆嫩、營養豐富。大量研究顯示,豆科種子發芽后,營養水平和抗氧化水平都有顯著提升:綠豆發芽3 d 后抗壞血酸含量增長2.7 倍,咖啡酸、兒茶素、阿魏酸、沒食子酸和蘆丁等酚類化合物逐漸增加;大豆發芽3 d 后異黃酮含量高達4.68 mg/g 干樣,豆芽中染料木黃酮、大豆苷元、總糖苷配基分別增加了3、2.4 和2.5 倍;豇豆發芽5 d 后總酚含量增加了1.7倍,抗氧化活性增加了2.8 倍,葉酸含量也有所增加。且萌發后豆類中的脂肪、碳水化合物、蛋白質等大分子物質會水解成氨基酸、寡糖等小分子物質,更易被人消化吸收,抗炎、抗菌、抗氧化性增強,營養價值提高。抗營養因子如棉子糖、水蘇糖含量在發芽后也會顯著降低,避免脹氣。水培活體蔬菜能夠提升活性物質含量與抗氧化能力,且清潔、健康、快速,符合當代生活節奏與綠色消費需求,是開發功能食品的理想方法。
紫云英產量高,營養豐富,易于生長,因此非常適合做芽苗菜開發。但到目前為止,關于其成分的研究較少,且紫云英的菜用價值還未被開發利用。本文在對比分析紫云英湖南種子與安徽種子的基本營養成分的基礎上,水培開發苗菜,對比分析兩產地苗菜總酚、總黃酮含量以及酚類物質組成并進行DPPH、ABTS 抗氧化評價,獲得苗菜發芽過程中的物質變化規律并優化芽菜培育工藝,以期拓寬菜用紫云英的資源化利用渠道,為提高紫云英經濟效益提供新思路。
1 材料與方法
1.1 材料與儀器
湘紫1 號 資陽區晟昱紫云英種植專業合作社;皖紫1 號和紫云英土培成熟莖葉(4 月25 日至5 月10 日播種,8 月25 日采摘)蕪湖青弋江種業有限公司;原兒茶酸(≥98%)、香草酸(≥98%)、綠原酸(≥98%)、對香豆酸(≥98%)、沒食子酸(≥98%)、槲皮素(≥98%)、阿魏酸(≥98%)、蘆丁(≥98%)、兒茶素(≥98%)、芥子酸(≥98%)、肉桂酸(≥98%)、表兒茶素(≥98%)索萊寶;芹菜素(≥98%)、山奈酚(≥98%)、丁香酸(≥98%)、水楊酸(≥98%)ANPEL Laboratory Technologies Inc.(Shanghai,China);對羥基苯甲酸(99.5%)、木犀草素(≥98%)ZZSTANDARD 甄準生物;ABTS(Ultra-Pure)AMRESCO;DPPH(>97%)東京化成工業株式會社;其他實驗試劑 均為國產分析純。
SMP500-15275-SWXN 型酶標儀 Molecular Devices;T6 NG 型分光光度計 北京普析通用儀器有限責任公司;KDY-9820 型半自動凱氏定氮儀 北京市通潤源機電技術有限責任公司;PRx-450C 型智能培養箱 寧波賽福實驗儀器有限公司。
1.2 實驗方法
1.2.1 紫云英種子基本成分的測定 參考國標GB 5009.3-2016 中的直接干燥法、GB 5009.5-2016 中的凱氏定氮法、GB 5009.9-2016 中的酶水解法、GB 5009.6-2016 中的索氏抽提法、GB/T 5009.10-2003 中的酸堿消煮法、GB/T 5009.4-2016 中的高溫灰化法測量湘紫1 號、皖紫1 號種子水分、蛋白質、淀粉、粗脂肪、粗纖維、總灰分含量。
1.2.2 紫云英苗菜的開發 浸種方法參考Mastropasqua 等的方法。用超過紫云英種子體積1 倍的1.05%~1.10%(體積分數)的鹽水室溫避光浸種2 h,清水洗凈后用適量0.1%(體積分數)的次氯酸鈉浸泡種子半小時,用蒸餾水洗至中性。將處理過的紫云英種子均勻播種于發芽盤中,溫度25 ℃、濕度80%黑暗培養3 d 后于光照80%的培養箱中發芽育苗,于發芽8、10、14 d 取樣測定酚類組成、總酚和總黃酮含量以及抗氧化能力,以土培成熟莖葉為對照。
1.2.3 酚類物質的提取 參考鐘少杰等的方法并稍作修改。用蒸餾水沖洗苗菜,50 ℃、12 h 烘干,粉碎密封備用。稱取0.5 g 該干燥粉末于150 mL 錐形瓶中,用體積分數31%乙醇以1:41 的料液比于73 ℃超聲提取106 min,抽濾取上清液,定容在25 mL 容量瓶中,用0.22 μm 濾膜過濾后儲存在小棕瓶中,?20 ℃備用。
1.2.4 總酚、總黃酮含量測定 總酚測定參考Folin-Ciocalteu 法,取醇提液100 μL,加入500 μL 福臨酚溶液,搖勻,再加400 μL 7.5% NaCO溶液,放入37 ℃培養箱中1 h 后,在765 nm 下測定吸光值,以沒食子酸標準品繪制標準曲線,結果以每克樣品中沒食子酸當量(gallic acid equivalents,GAE)表示測定樣品(干物質)中的總酚含量。標準曲線方程為y=4.9012x+0.1193,=0.9932。
總黃酮測定參考黃酮沉淀法,取醇提液500 μL,加入500 μL 的20%鹽酸溶液和250 μL 37%甲醛溶液,搖勻,放在暗處靜置24 h。之后取出1 mL 于離心管中離心(12000 r/min,10 min),除去白色沉淀,上清備用。每個再分別加入500 μL 福臨酚溶液,搖勻,再加400 μL 7.5% NaCO溶液,放入37 ℃培養箱中1 h 后,在765 nm 下測定吸光值,以沒食子酸標準品繪制標準曲線計算濾液中總酚含量,總酚含量減去濾液總酚含量即為總黃酮含量,結果以每克樣品中沒食子酸當量(gallic acid equivalents,GAE)表示測定樣品(干物質)中的總黃酮含量。
1.2.5 酚類成分測定 色譜柱:ACQUITY UPLC HSS T3(2.1 mm×100 mm,1.8 μm);流動相:流動相A 為乙腈,流動相B 為0.05%甲酸水溶液,梯度洗脫程序見表1;流速為0.3 mL/min;柱溫:30 ℃;進樣量為5 μL;自動進樣器溫度為20 ℃;負離子模式下,毛細管電壓2.90 kV,離子源溫度120 ℃,脫溶劑氣溫度400 ℃,錐孔氣(N)流速50 L/h,脫溶劑氣(N)流速600 L/h,碰撞氣(Ar)流速0.07 mL/min。正離子模式下,毛細管電壓3.10 kV,其余條件與負離子模式相同。結果以mg/g DW 表示。
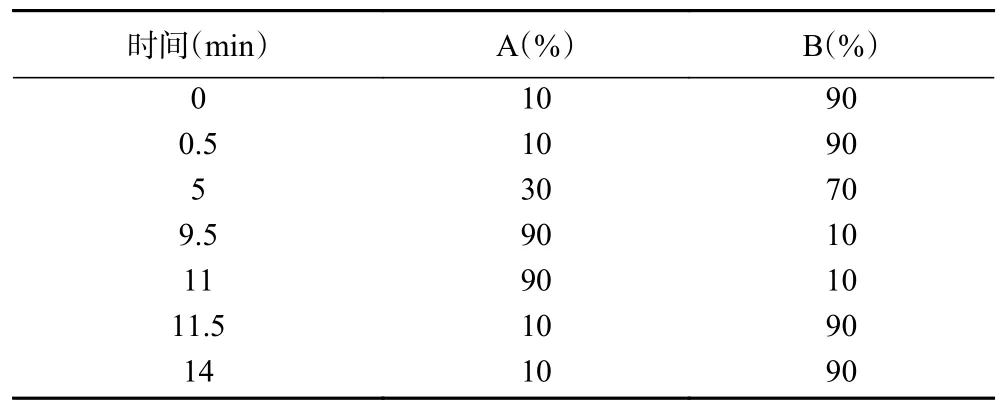
表1 UPLC 洗脫程序Table 1 Elution program of UPLC
1.2.6 DPPH 自由基清除率的測定 參考Koodkaew的方法。用乙醇配制0.1 mmol/L DPPH 溶液,避光保存備用。取96 孔板,每孔加入180 μL DPPH 溶液和紫云英醇提液溶液20 μL,振蕩混勻后,室溫避光放置15 min,于517 nm 波長處測定吸光度。以Trolox 溶液作為陽性對照,結果以Trolox 當量(mmol TE/L)和IC(mg/mL)表示。DPPH 自由基清除率按式(1)計算。
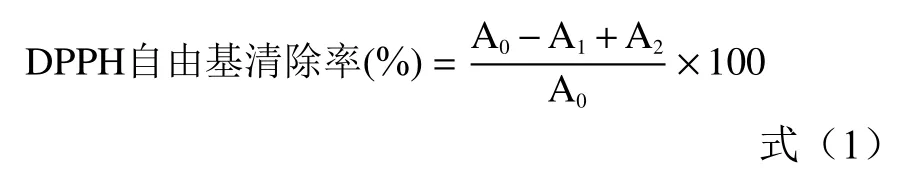
式中:A為樣品吸光度;A為用無水甲醇代替DPPH 同法測定吸光度;A為以樣品溶劑溶劑即31%乙醇代替樣品同法測定吸光度。
1.2.7 ABTS自由基清除率的測定 參考Islam 等的方法。取7 mmol/L 的ABTS 與1.4 mmol/L 的過硫酸鉀溶液等體積混合均勻,避光室溫放置16 h,臨用前用95%乙醇溶液以1:40 體積比將混合液在734 nm 下的吸光值調整為0.7±0.02,得ABTS 工作液。取工作液5 mL、紫云英苗菜醇提液0.5 mL,混合均勻,反應6 min 后用分光光度計于734 nm 波長處測定吸光度。以Trolox 溶液為陽性對照,結果以Trolox 當量(mmol TE/L)和IC(mg/mL)表示。ABTS自由基清除率按式(2)計算。
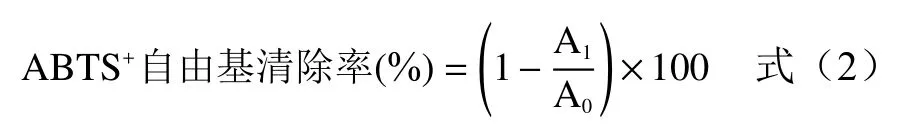
式中:A為樣品吸光度;A為以溶劑即31%乙醇代替樣品同法測得的吸光度。
1.3 數據處理
運用Excel 2013 整理各指標的測定結果,采用SPSS 25.0 進行差異性分析和相關性分析,每個處理重復3 次,用Origin 2021 作圖。
2 結果與分析
2.1 紫云英種子基本成分
蛋白質是紫云英種子的優勢成分,湘紫1 號和皖紫1 號種子蛋白質含量分別為33.22%±0.62%和38.66%±0.41%。豆科植物蛋白質含量豐富,但不同豆類的蛋白質含量差異較大。大豆蛋白含量相對較高,約為33.1%~49.2%,綠豆中含蛋白質14.6%~33.0%,鷹嘴豆為17%~22%,赤小豆為23.17%~25.57%。由此對比,紫云英種子蛋白質含量與大豆相當,十分豐富。紫云英種子的脂肪含量(表2)與鷹嘴豆(6.48%)和赤小豆(1.92%~3.42%)相似,低于大豆(14.13%~22.19%)脂肪含量,淀粉含量也低于綠豆淀粉含量(約50%)。綜上所述,紫云英種子具有高蛋白、低淀粉、低脂肪的特點,且皖紫1 號種子蛋白質含量可與大豆媲美,是一種良好的蛋白質補充劑。
由表2 可知,皖紫1 號種子蛋白質明顯高于湘紫1 號,粗脂肪含量為湘紫1 號的1.3 倍,灰分含量更低,淀粉、粗纖維、水分與湘紫1 號無明顯差異。這說明從基本營養成分角度考慮,皖紫1 號種子營養價值高于湘紫1 號。
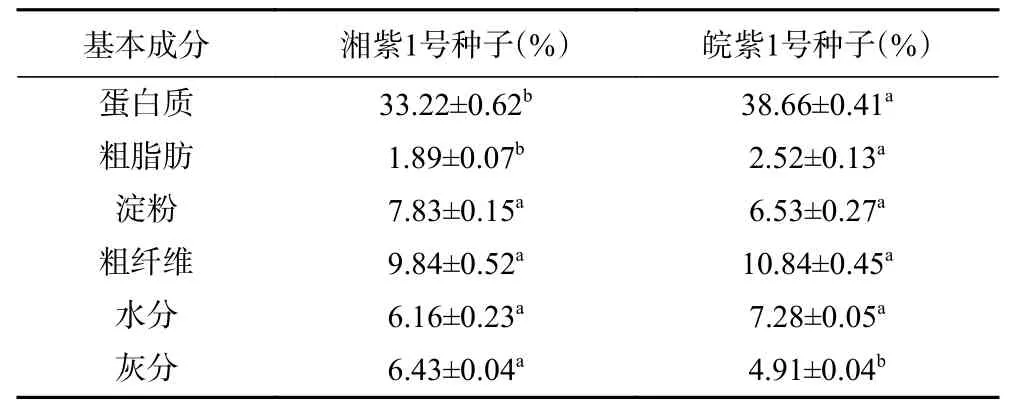
表2 紫云英種子基本成分Table 2 Basic components of Xiang seeds and Wan seeds
2.2 紫云英苗菜生長過程
圖1 為紫云英苗菜第1~14 d 的生長情況。第3 d(圖1C)形成胚根,第4 d(圖1D)長出兩片子葉,下胚軸和胚根伸長,葉片長大。第5~10 d(圖1E~圖1J),胚根由0.75 cm 生長到3.45 cm,子葉葉片更大更密,第5 d 長出真葉,之后下胚軸長度基本不變,上胚軸長度變長,真葉葉片變大,10 d 后外觀形態趨于穩定,莖高在6 cm 左右。

圖1 湘紫1 號苗菜生長過程Fig.1 Vegetation process of Xiang sprouts
2.3 紫云英苗菜總酚、總黃酮含量的變化
如圖2 和圖3 所示,湘紫1 號、皖紫1 號苗菜萌發過程中總酚、總黃酮含量呈上升趨勢,均在第14 d 時達到最大。這與小麥、綠豆萌發后次級產物積累趨勢類似。這是因為豆類種子萌發過程中,莽草酸途徑、苯丙烷代謝途徑和類黃酮代謝途徑關鍵酶、限速酶如苯丙氨酸解氨酶(PAL)、肉桂酸-4-羥化酶(C4H)、4-香豆酰輔酶A 連接酶(4CL)、査耳酮合成酶(CHS)等酶活上升,促進糖酵解的中間產物磷酸烯醇式丙酮酸(EMP)和赤蘚糖-4-磷酸(PPP)合成植物多酚、黃酮等次級代謝產物。
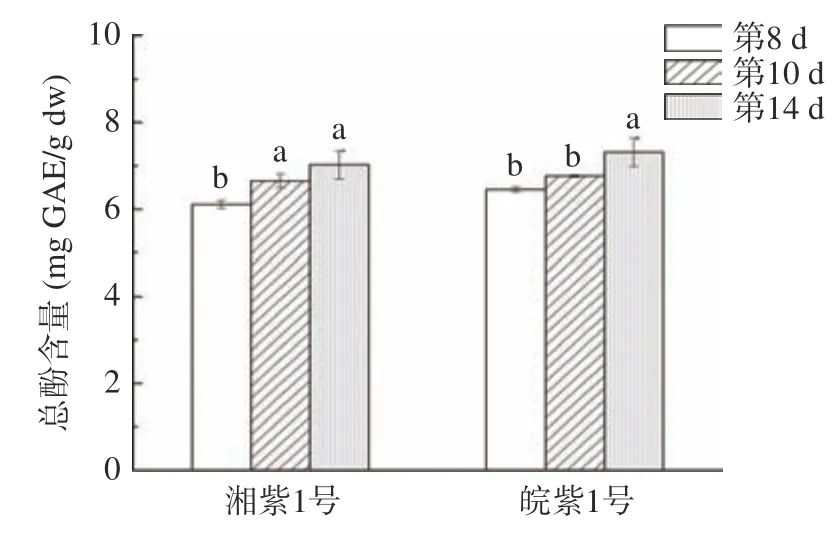
圖2 苗菜總酚含量Fig.2 Total phenolic content in CMV sprouts
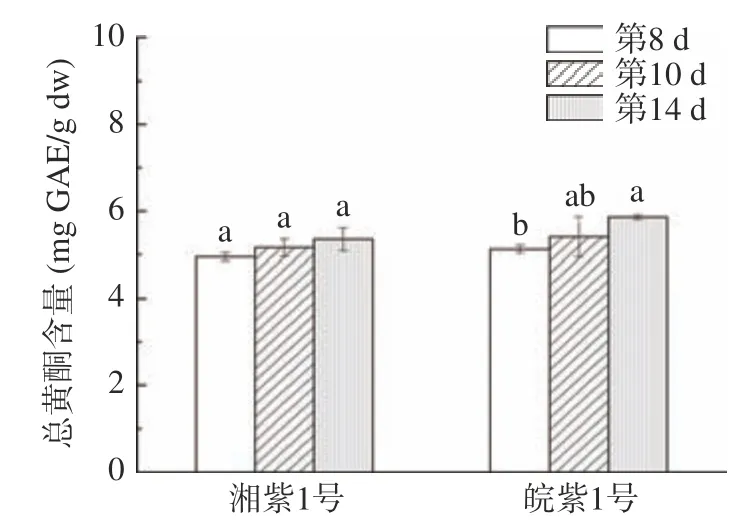
圖3 苗菜總黃酮含量Fig.3 Total flavonoids content in CMV sprouts
皖紫1 號苗菜總酚、總黃酮含量普遍高于湘紫1 號,發芽14 d 的皖紫1 號總酚、總黃酮在水培苗菜中最高。萌發8~14 d 中,皖紫1 號苗菜總酚含量由6.45 mg/g 上升至7.31 mg/g,增長13.33%,總黃酮含量由5.12 mg/g 上升至5.86 mg/g,含量上升14.45%,分別達到生長3 個月土培成熟莖葉總酚含量(12.96±0.23 mg/g)、總黃酮含量(10.55±0.13 mg/g)的50%和45%,可見在總酚、總黃酮方面,發芽14 d 的皖紫1 號單位時間投入產出比更高。
2.4 紫云英酚類成分分析
如表3,在紫云英苗菜、土培成熟莖葉中共檢測出14 種酚類化合物,其中7 種酚酸,5 種黃酮類化合物,2 種多酚化合物。在紫云英苗菜、土培成熟莖葉中均未檢測出原兒茶酸、對羥基苯甲酸、表兒茶素和肉桂酸。

表3 湘紫1 號酚類物質含量Table 3 Basic components of Xiang seeds and Wan seeds
在湘紫1 號苗菜和皖紫1 號苗菜發芽8~14 d的過程中,酚類化合物總量隨發芽時間升高,與總酚、總黃酮含量變化趨勢一致。皖紫1 號苗菜酚類化合物含量顯著高于湘紫1 號苗菜,發芽8、10、14 d 時皖紫1 號苗菜酚類物質總量是湘紫1 號苗菜的1.28、1.21、1.42 倍,在發芽14 d 時達到41.28±1.85 mg/kg。
酚類物質具有抗氧化、抗腫瘤、調節成骨細胞分化、神經免疫調節和神經保護作用等多種藥理學活性。發芽14 d 的皖紫1 號苗菜酚類化合物種類豐富,其中共檢測出12 種酚類化合物,比土培成熟莖葉多檢測出沒食子酸、阿魏酸、香草酸、水楊酸、丁香酸5 種酚類物質。發芽14 d 的皖紫1 號苗菜優勢成分為山奈酚、芹菜素、槲皮素、對香豆酸、沒食子酸、木犀草素和阿魏酸,其中,山奈酚、對香豆酸、沒食子酸含量分別為發芽14 d 湘紫1 號苗菜的1.67、1.69 和13 倍,山奈酚含量(12.86±0.34 mg/kg)是豌豆尖山奈酚含量的2~15 倍,槲皮素含量與豌豆尖相當,對香豆酸含量為4.59±0.13 mg/kg,約為土培成熟莖葉的33 倍。
發芽14 d 的皖紫1 號苗菜酚類含量豐富,酚類物質的單位時間投入產出比優于湘紫1 號和土培成熟莖葉,有進一步開發利用潛力。短時開發酚類物質豐富的紫云英菜品,可選擇皖紫1 號水培14 d。
2.5 紫云英苗菜抗氧化評價
2.5.1 DPPH 自由基清除能力評價 如圖4A 所示,紫云英苗菜在8~14 d 對DPPH 自由基的IC與發芽天數呈負相關,發芽14 d 湘紫1 號苗菜IC為15.18 mg/mL,是其發芽8 d IC(21.77 mg/mL)的70%;發芽14 d 皖紫1 號苗菜IC為9.53 mg/mL,是其發芽8 d IC(19.44 mg/mL)的49%。可見,隨發芽天數的增加,湘紫1 號和皖紫1 號苗菜對DPPH自由基的IC顯著下降,清除能力逐漸增強。同時,皖紫1 號苗菜IC值明顯低于湘紫,發芽14 d 皖紫1 號苗菜IC是同期湘紫的63%,是生長三個月土培成熟莖葉(4.76 mg/mL)的2.00 倍,可以得出,皖紫1 號苗菜對DPPH 自由基的清除能力優于湘紫1 號苗菜,發芽14 d 皖紫1 號苗菜DPPH 自由基清除能力在水培苗菜中最強,但弱于土培成熟莖葉。
如圖4B 和圖4C 所示,苗菜對DPPH 自由基的清除能力在0.5~15 mg/mL 的范圍內呈濃度依賴性。隨濃度的增加,發芽14 d 的皖紫1 號苗菜對DPPH 自由基的清除能力由13.97 mmol TE/L 升至22.64 mmol TE/L,成熟莖葉對DPPH 自由基清除能力由16.92 mmol TE/L 升至37.29 mmol TE/L。因此,在0.5~15 mg/mL 范圍內,可以通過提高水培苗菜提取液濃度,提高DPPH 清除能力,從而通過14 d水培得到DPPH 自由基清除能力較強的產品。
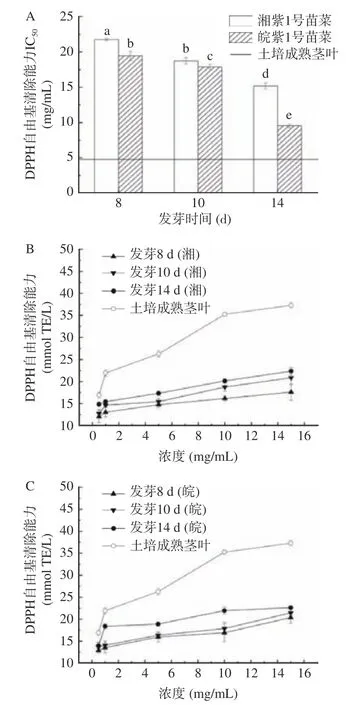
圖4 紫云英苗菜DPPH+自由基清除能力Fig.4 DPPH+ scavenging activity of CMV sprouts
2.5.2 苗菜ABTS自由基清除能力評價 如圖5A所示,發芽8~14 d 的湘紫和皖紫苗菜對ABTS自由基的IC與發芽天數呈負相關,隨發芽天數的增加,湘紫苗菜對ABTS自由基的IC值由8.18 mg/mL下降至6.08 mg/mL,皖紫對ABTS自由基的IC值由7.19 mg/mL 顯著下降至4.08 mg/mL,湘紫和皖紫苗菜在發芽第8~14 d 內對ABTS自由基的清除能力隨發芽天數的增加而增強。發芽8~14 d,湘紫苗菜發芽對ABTS自由基的IC值均明顯高于相同天數的皖紫苗菜,即皖紫苗菜對ABTS自由基的清除能力優于湘紫苗菜。
如圖5B 和圖5C 所示,發芽14 d 的湘紫苗菜清除能力由2.43 mmol TE/L 上升至20.96 mmol TE/L,發芽14 d 的皖紫苗菜清除能力由2.20 mmol TE/L 上升至20.8 mmol TE/L,而土培成熟莖葉由4.57 mmol TE/L 上升至21.19 mmol TE/L,即苗菜對ABTS自由基的清除能力在0.5~15 mg/mL 的范圍內與濃度成正相關,隨濃度的增加,苗菜對ABTS自由基的清除能力逐漸上升。
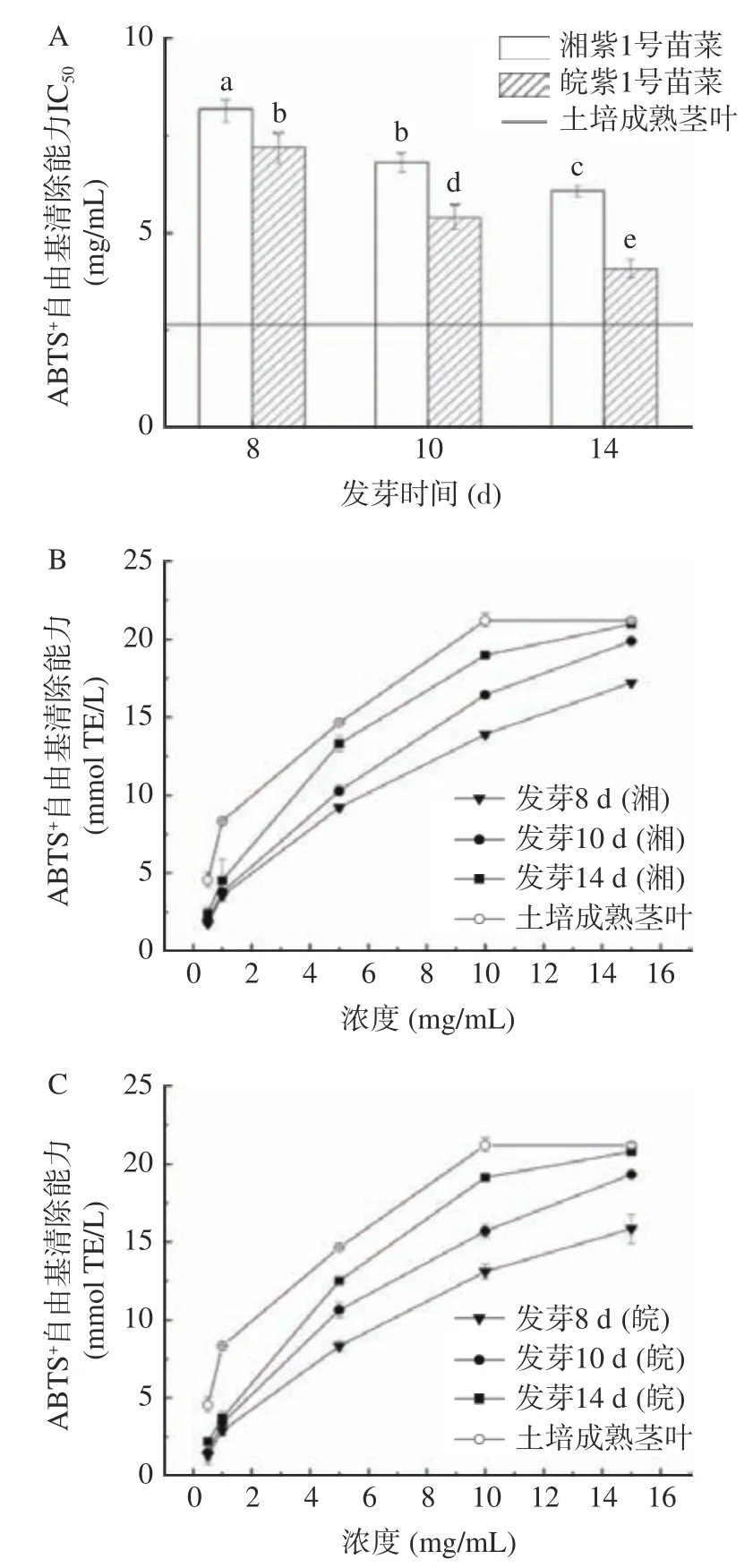
圖5 紫云英苗菜ABTS+自由基清除能力Fig.5 ABTS+ scavenging activity of CMV sprouts
與土培成熟莖葉相比,兩類苗菜對ABTS自由基的清除能力較弱,但當濃度到達15 mg/mL 時,發芽14 d 的湘紫苗菜和皖紫苗菜對ABTS自由基的清除能力與土培成熟莖葉相當,這說明發芽14 d 后,紫云英苗菜具有可觀的抗氧化性,而且水培的紫云英苗菜耗時短,成本低,與土培相比更有潛力獲得經濟回報。
2.5.3 苗菜酚類物質含量與抗氧化性的相關性分析
如表4,湘紫1 號苗菜和皖紫1 號苗菜的總酚、總黃酮含量與DPPH、ABTS自由基清除能力顯著相關,研究人員在水稻、綠豆芽苗菜、桃子中也觀察到了這一現象。山奈酚、槲皮素、阿魏酸在兩種紫云英苗菜中與DPPH、ABTS自由基清除能力顯著相關,表示這三種酚類化合物對2 種紫云英水培苗菜抗氧化能力的貢獻率較大。皖紫1 號苗菜中木犀草素、對香豆酸、沒食子酸與DPPH 和ABTS自由基清除能力顯著相關,而這三種酚類化合物在湘紫1 號中沒有表現出較高相關性,從而解釋了皖紫1 號苗菜抗氧化性強于湘紫1 號的原因。
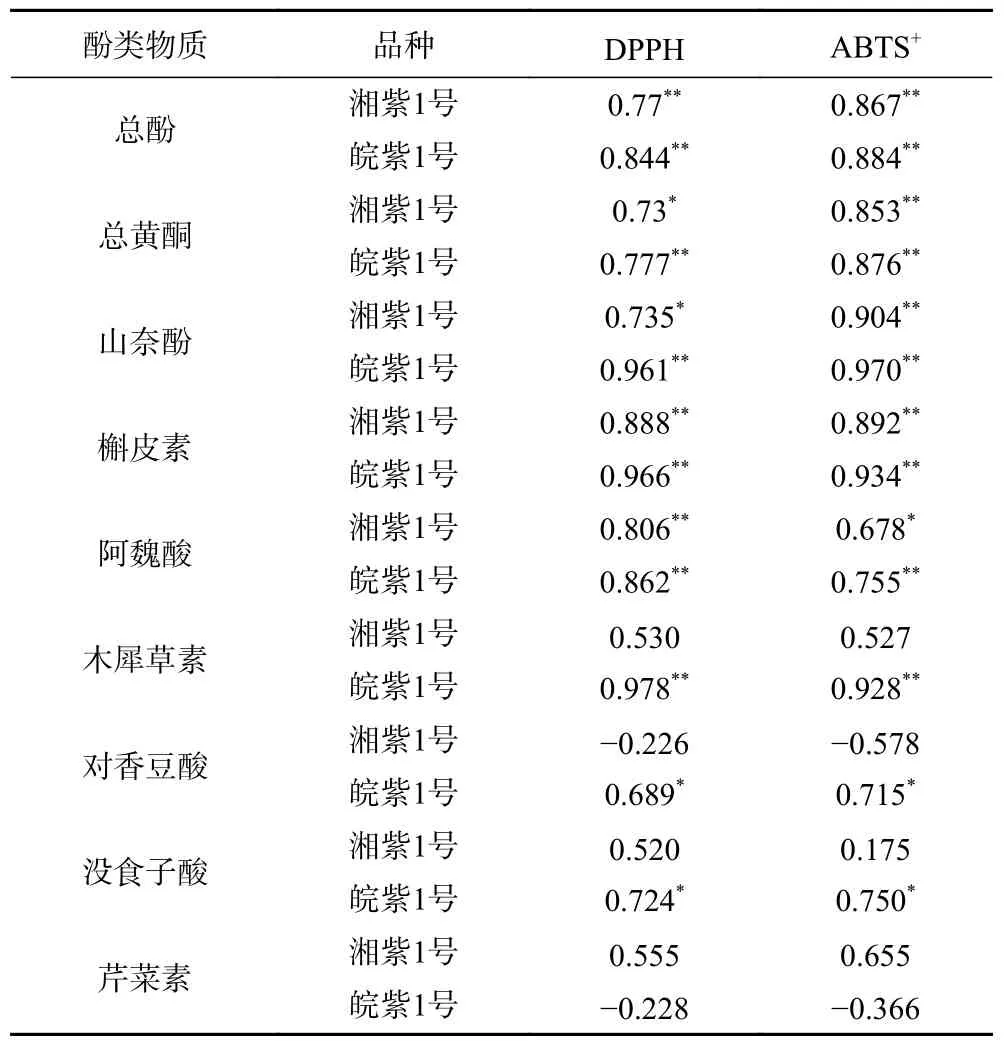
表4 苗菜酚類含量與抗氧化性之間的相關性Table 4 Correlation of phenolics and antioxidant activities
植物體展示出抗氧化性是一個復雜體系相互作用的結果。伴隨種子萌發,植物體中有非酶與酶促兩個抗氧化系統共同維持著細胞內的氧化還原平衡。非酶方面,游離氨基酸、抗壞血酸、酚類化合物都具有直接清除自由基的作用,皖紫1 號種子蛋白含量比湘紫1 號種子更豐富,萌發過程中皖紫1 號中更豐富的蛋白質可能會分解產生更豐富的游離氨基酸,從而表現出更強的抗氧化作用;酶促抗氧化系統方面,隨著萌發過程中細胞內活性氧水平的上升,過氧化氫、丙二醛(MDA)等氧化代謝產物逐漸積累,超氧化物歧化酶(SOD)、過氧化物酶(POD)、過氧化氫酶(CAT)、抗壞血酸過氧化物酶(APX)的酶活和相關基因表達量也可能影響苗菜的抗氧化性。本文從酚類化合物角度揭示了皖紫1 號苗菜抗氧化性較強的原因,為紫云英苗菜選種、育苗提供了初步理論支撐,下一步可以從游離氨基酸和酶促角度進一步深挖紫云英水培苗菜發芽過程中抗氧化動態機制,得到營養物質含量更豐富、功能性更強的產品。
3 結論
本文探究了湘紫1 號、皖紫1 號兩種紫云英種子水培開發過程中酚類物質和抗氧化性的動變化,從酚類化合物角度揭示了皖紫1 號苗菜抗氧化性較強的原因。結果表明,兩類紫云英苗菜生長過程中總酚、總黃酮含量以及抗氧化能力均呈上升趨勢,15 mg/mL 濃度下,發芽14 d 兩類苗菜對ABTS自由基的清除能力可媲美土培成熟莖葉;水培14 d 的皖紫1 號苗菜酚類含量豐富,總酚、總黃酮含量達到7.31 和5.86 mg/g,共檢測出12 種酚酸化合物,比土培成熟莖葉多檢測出5 種酚類物質,對香豆酸含量達到土培成熟莖葉的33 倍;皖紫苗菜培育周期短,成本較低,酚類物質和抗氧化能力較強,單位時間投入產出比更高,因此更有潛力獲得經濟回報。下一步可以從游離氨基酸和酶促角度進一步深挖紫云英水培苗菜發芽過程中抗氧化動態機制,為紫云英水培苗菜的開發提供更為全面的科學依據。
猜你喜歡
發明與創新(2022年30期)2022-10-03 08:40:56
四川蠶業(2021年3期)2021-02-12 02:38:46
動漫星空(興趣百科)(2020年12期)2020-12-12 05:31:40
中成藥(2018年11期)2018-11-24 02:57:00
人大建設(2018年6期)2018-08-16 07:23:10
文理導航·科普童話(2017年5期)2018-02-10 19:42:14
中成藥(2017年8期)2017-11-22 03:19:40
中成藥(2017年10期)2017-11-16 00:50:13
無人機(2017年10期)2017-07-06 03:04:36
中成藥(2017年4期)2017-05-17 06:09:50
