外源添加L-蘋(píng)果酸降低秀麗隱桿線蟲(chóng)體內(nèi)脂肪含量
2022-09-16 07:22:06王潤(rùn)圓張夢(mèng)媛
中國(guó)食物與營(yíng)養(yǎng) 2022年8期
機(jī)體中,L-蘋(píng)果酸參與蘋(píng)果酸天冬氨酸轉(zhuǎn)運(yùn)過(guò)程,具有轉(zhuǎn)移還原型煙酰胺腺嘌呤二核苷酸、還原型輔酶I(NADH)的作用
。此外,L-蘋(píng)果酸對(duì)機(jī)體有較多有益的生理功能,如運(yùn)動(dòng)過(guò)程中降低血清肌酸激酶水平,減少骼肌損傷
;減少脂質(zhì)過(guò)氧化增強(qiáng)機(jī)體抗氧化能力
;預(yù)防老年性癡呆等
。L-蘋(píng)果酸作為低熱量的酸味食品添加劑廣泛應(yīng)用于各類食品等的生產(chǎn)加工,可保持食品的口感和色澤
。秀麗隱桿線蟲(chóng)作為一種實(shí)驗(yàn)?zāi)J缴铮子谂囵B(yǎng)、遺傳操作簡(jiǎn)單,通體透明便于觀察。更重要的是,哺乳動(dòng)物中發(fā)現(xiàn)的許多核心代謝通路在線蟲(chóng)中相當(dāng)保守,包括脂肪酸合成、延伸、去飽和和β-氧化,以及神經(jīng)肽、5 -羥色胺和胰島素信號(hào)通路等。本研究通過(guò)外源添加L-蘋(píng)果酸探索其在秀麗隱桿線蟲(chóng)脂代謝過(guò)程中的作用和機(jī)制。
1 材料與方法
1.1 材料與試劑
L-蘋(píng)果酸,上海生工生物工程有限公司;RNA提取試劑盒、c-DNA合成試劑盒,天根生化科技(北京)有限公司;qPCR引物,由上海生工生物工程有限公司合成;SYBR Green,北京聚合美生物科技有限公司;尼羅紅、油紅染料,美國(guó)Sigma-Aldrich公司;
-6:GFP和
-7:GFP線蟲(chóng),由云南生物資源保護(hù)與利用國(guó)家重點(diǎn)實(shí)驗(yàn)室饋贈(zèng)。
1.2 儀器與設(shè)備
倒置熒光顯微鏡,日本Nikon公司;超聲破碎儀,寧波新芝生物科技公司;酶標(biāo)儀,美國(guó)BioTek儀器有限公司;PCR儀,德國(guó)Biometra公司;電泳儀、凝膠成像系統(tǒng),美國(guó)Bio-Rad公司;實(shí)時(shí)熒光定量PCR儀,德國(guó)Eppendorf公司;低溫高速離心機(jī),美國(guó)貝克曼公司;震蕩儀,海門市其林貝爾儀器制造有限公司;旋渦混合器,北京金紫光科技發(fā)展有限公司。
由表8可知隨著染色溫度的增加,棉條表觀深度K/S值提高,但溫度超過(guò)70℃以后,K/S值減小。這是由于溫度升高,染料加速擴(kuò)散,上染速率提高,同時(shí)染料水解速率提高,但在一定的溫度范圍內(nèi),溫度提高,染料上染速率大于水解速率,但是當(dāng)超過(guò)合適溫度時(shí),染料的水解速率大于上染速率,所以上染率降低,棉條得色淺,因此選擇染色溫度70℃為宜。
1.3 方法
1.3.1 秀麗隱桿線蟲(chóng)培養(yǎng) 使用大腸桿菌OP50菌株作為食物源,將其均勻涂于線蟲(chóng)生長(zhǎng)的培養(yǎng)基(NGM)上。將同步化后的800~1 000條左右的L1的線蟲(chóng)接種于涂布了OP50的NGM平板上,20 ℃培養(yǎng)2 d至產(chǎn)1~3顆卵的成蟲(chóng)期備用。
1.3.7 qPCR分析 收集L4期線蟲(chóng),按寶生物工程(大連)有限公司的RNA提取試劑盒的步驟提取線蟲(chóng)的總RNA,并測(cè)定其含量。按天根生化科技有限公司的逆轉(zhuǎn)錄試劑盒步驟來(lái)合成cDNA,反應(yīng)體系:SYBR green mix 10 μL、上游引物0.5 μL、下游引物0.5 μL、cDNA 2 μL、ddH
O 10 μL,使用羅氏熒光定量PCR儀進(jìn)行定量。我們以
-1為內(nèi)參基因,用2
計(jì)算基因轉(zhuǎn)錄水平差異,引物序列見(jiàn)附表。
1.3.3 油紅O(ORO)染色 500 mg ORO粉溶于100 mL異丙醇,制成母液避光密封保存?zhèn)溆谩J褂们耙匀ルx子水按3∶2比例稀釋異丙醇油紅O母液,震蕩2 h使其充分溶解,用0.2 μm微孔濾膜過(guò)濾雜質(zhì)制備為工作液,錫箔紙包裹避光冰箱儲(chǔ)存?zhèn)溆谩R訮BST(含0.01% Triton)將攜帶1~3個(gè)蟲(chóng)卵的成蟲(chóng)輕洗于1.5 mL離心管中,反復(fù)清洗3次至澄清狀態(tài)無(wú)菌。以560 g離心1 min去掉上清,留100 μL重懸,加入600 μL 40%異丙醇在室溫下震蕩孵育3 min,560 g離心1 min,去上清,留100 μL重懸。加入600 μL ORO工作液混勻,將樣品在室溫下避光旋轉(zhuǎn)孵育2 h,560 g 離心1 min,去上清留100 μL。加入600 μL PBST重懸,在避光條件下旋轉(zhuǎn)孵育30 min去除多余ORO染色劑,560 g離心1 min,留50 μL上清液。取15 μL線蟲(chóng)懸浮液置于載玻片上,相同曝光時(shí)間進(jìn)行白光拍照。實(shí)驗(yàn)重復(fù)3次,每個(gè)組拍攝不少于20條。
3.2.5 避免生理鹽水沖洗氣管插管 生理鹽水滴注到氣管插管造成管壁生物膜沖入肺內(nèi),易形成感染。研究認(rèn)為,生理鹽水沖洗氣管插管沒(méi)有起到稀釋痰液的作用,反而使肺的氧合下降,血壓及心率增快,顱內(nèi)壓增高及發(fā)生VAP風(fēng)險(xiǎn)增大[47]。
ECDSA是數(shù)字簽名算法(DSA)的其中一個(gè)例子。和非對(duì)稱加密算法(RSA)進(jìn)行對(duì)比,在相同的安全強(qiáng)度下,ECDSA可以使用的密鑰更短,從而節(jié)省網(wǎng)絡(luò)和存儲(chǔ)空間,具有較高的研究?jī)r(jià)值[5]。
秀麗隱桿線蟲(chóng)體脂主要儲(chǔ)存于腸上皮細(xì)胞和腸道中,ORO和NR染色法能客觀地展示秀麗隱桿線蟲(chóng)體內(nèi)脂肪
。采用ORO染色、NR染色和TG含量測(cè)定3種方法對(duì)線蟲(chóng)體內(nèi)脂肪進(jìn)行分析。將同步化后500條左右L1線蟲(chóng)分別加至含L-蘋(píng)果酸0、200、500、1 000 μg/mL的NGM平板,培養(yǎng)至攜帶1~3個(gè)蟲(chóng)卵的成蟲(chóng),進(jìn)行ORO染色和NR染色。其中NR染色表明,與對(duì)照組(0 μg/mL)相比,200、500、1 000 μg/mL L-蘋(píng)果酸喂養(yǎng)線蟲(chóng)脂肪顆粒數(shù)量減少(圖1A);脂肪顆粒體積減小(圖1A),由1.66 μm分別減小至1.43、1.17、1.05 μm(圖1C)(
<0.05)。ORO染色表明,與對(duì)照組(0 μg/mL)相比,200、500、1 000 μg/mL L-蘋(píng)果酸喂養(yǎng)線蟲(chóng)脂肪含量分別降至83.33%、87.67%、76.67%(圖1B、D)(
<0.05)。同時(shí),根據(jù)TG含量的測(cè)量結(jié)果,與對(duì)照組(0 μg/mL)相比,200 、500 、1 000 μg/mL L-蘋(píng)果酸喂養(yǎng)線蟲(chóng)TG含量分別降至0.49、0.61、0.82倍(圖1E)(
<0.05)。綜上所述,外源添加200、500、1 000 μg/mL L-蘋(píng)果酸喂養(yǎng)線蟲(chóng),脂肪顆粒的數(shù)量減少、體積減小,降低體內(nèi)脂肪含量。
1.3.2 脂肪酸測(cè)定 培養(yǎng)線蟲(chóng)至產(chǎn)1~3顆卵的成蟲(chóng)期,用單蒸水洗脫5~10個(gè)培養(yǎng)皿的線蟲(chóng)至8 mL GC專用玻璃管中,置于冰上沉淀。單蒸水清洗2次,待蟲(chóng)體沉淀后去除上清。液氮速凍保存于-80 ℃冰箱。加入1 mL酯化液,70 ℃水浴鍋酯化1~2 h,冷卻后加入1.5 mL雙蒸水和200 μL正己烷,劇烈震蕩。4 000 r/min離心2 min后分2層,上層有機(jī)相為酯化脂肪酸。取上層有機(jī)相80 μL置于GC玻璃瓶中(加入襯管),上機(jī)檢測(cè)
。

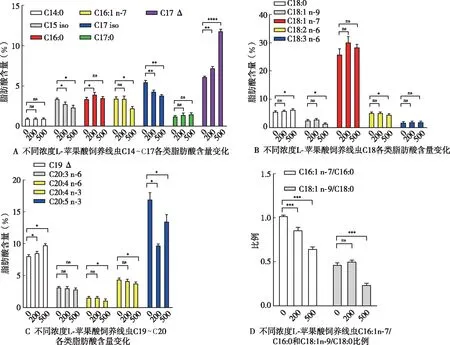
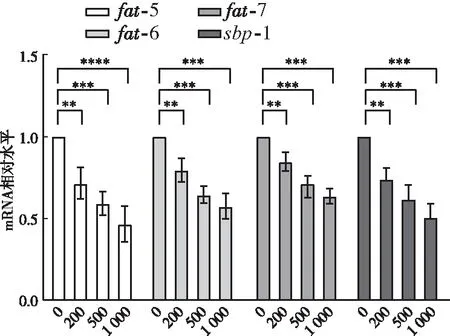
外源添加L-蘋(píng)果酸降低了C16:1n-7/C16:0和C18:1n-9/C18:0的比值,硬脂酰輔酶a去飽和酶(SCD)是將棕櫚酸C16:0和硬脂酸C18:0分別轉(zhuǎn)化為棕櫚油酸C16:1n-7和油酸C18:1n-9的關(guān)鍵酶
,接下來(lái)檢測(cè)L-蘋(píng)果酸對(duì)線蟲(chóng)3個(gè)SCD基因
-5、
-6、
-7的影響。qPCR結(jié)果顯示,與對(duì)照組(0 μg/mL)相比,200、500、1 000 μg/mL L-蘋(píng)果酸飼養(yǎng)線蟲(chóng),
-5基因mRNA相對(duì)水平分別下調(diào)為0.71、0.59、0.46倍(
<0.05)(圖3);
-6基因mRNA相對(duì)水平分別下調(diào)為0.79、0.64、0.57倍(
<0.05)(圖3);
-7基因mRNA相對(duì)水平分別下調(diào)為0.84、0.69、0.63倍(
<0.05)(圖3)。此外,檢測(cè)了L-蘋(píng)果酸對(duì)SCD基因轉(zhuǎn)錄因子
-1的影響,與對(duì)照組(0 μg/mL)相比,200 、500、1 000 μg/mL L-蘋(píng)果酸飼養(yǎng)線蟲(chóng)
-1 基因mRNA相對(duì)水平分別下調(diào)為0.74、0.62、0.50倍(
<0.05)(圖3),表明L-蘋(píng)果酸抑制轉(zhuǎn)錄因子
-1的表達(dá),從而下調(diào)
-5、
-6和
-7基因的轉(zhuǎn)錄。
2 結(jié)果與討論
2.1 外源添加L-蘋(píng)果酸可減少秀麗隱桿線蟲(chóng)體內(nèi)脂肪含量
1.3.6 FAT-6:GFP和FAT-7:GFP熒光強(qiáng)度觀察 將同步化后孵化的
-6:GFP、
-7:GFP L1線蟲(chóng)轉(zhuǎn)移至含L-蘋(píng)果酸0、200、500、1 000 μg/mL 的NGM培養(yǎng)皿中,在標(biāo)準(zhǔn)條件下培養(yǎng)至攜帶1~3個(gè)蟲(chóng)卵的成蟲(chóng)。M9洗下后吸取15 μL制片,在熒光顯微鏡下使用FITC/GFP通道觀察。

2.2 外源添加L-蘋(píng)果酸對(duì)脂肪酸含量的影響
將同步化后500條左右L1線蟲(chóng)分別加至含L-蘋(píng)果酸0、200、500 μg/mL的培養(yǎng)基中培養(yǎng)線蟲(chóng)至攜帶1-3個(gè)蟲(chóng)卵的成蟲(chóng),分別收集5-10個(gè)培養(yǎng)皿的線蟲(chóng)測(cè)定各類脂肪酸含量。與對(duì)照組(0 μg/mL)相比,200 μg/mL L-蘋(píng)果酸飼養(yǎng)線蟲(chóng)后,脂肪酸含量C15 iso由3.42%降至2.8%、C17 iso由5.5%降至4.29%、C20:5n-3由17.00%降至9.77%(
<0.05),脂肪酸含量C16:0由3.4%增至4%、C17 Δ由6.13%增至7.23%、C19 Δ含量由8.10%增至8.59%(
<0.05),脂肪酸含量C14:0由0.95%變?yōu)?.92%、C16:1n-7由3.47%變?yōu)?.44%、C17:0由1.24%變?yōu)?.45%、C18:0由5.56%變?yōu)?.94%、C18:1n-9由2.66%變?yōu)?.00%、C18:1n-7由25.96%變?yōu)?0.23%、C18:2n-6由5.34%變?yōu)?.35%、C18:3n-6由1.88%變?yōu)?.02%、C20:3n-6由3.17%變?yōu)?.12%、C20:4n-6由1.64%變?yōu)?.62%、C20:4n-3由4.44%變?yōu)?.20%,含量無(wú)明顯變化(
>0.05)(圖2)。500 μg/mL L-蘋(píng)果酸喂養(yǎng)線蟲(chóng)后,C15 iso由3.42%降至2.40%、C16:1n-7由3.48%降至2.28%、C17 iso由5.50%降至3.83%、C18:2n-6由5.35%降至4.62%、C18:1n-9由2.66%降至1.53%、C20:4n-6由1.62%降至1.15%、C20:4n-3由4.44%降至3.81%、C20:5n-3由17.00%降至13.55%(
<0.05),脂肪酸C17 Δ由6.13%增至11.85%、C18:0由5.67%增至6.37%、C19 Δ由8.10%增至9.83%(
<0.05),脂肪酸C14:0由0.95%變?yōu)?.94%、C16:0由3.40%變?yōu)?.50%、C17:0由1.24%變?yōu)?.51%、C18:1n-7由25.96%變?yōu)?8.51%、C18:3n-6由1.88%變?yōu)?.05%、C20:3n-6由3.17%變?yōu)?.88%,含量無(wú)明顯變化(
>0.05)(圖2A-C)。此外,與對(duì)照組(0 μg/mL)相比,200、500 μg/mL L-蘋(píng)果酸喂養(yǎng)線蟲(chóng)后C16:1n-7/C16:0由1.02降至0.86和0.65(
<0.05)(圖2D);C18:1n-9/C18:0的比例由0.47變?yōu)?.50(
>0.05)和0.24(
<0.05),C16:1n-7/C16:0和C18:1n-9/C18:0比例降低。脂肪中的單不飽和脂肪酸棕櫚油酸C16:1n-7和油酸C18:1n-9是生物合成甘油三酯、磷脂、膽固醇酯等的重要底物。C16:1n-7/C16:0和C18:1n-9/C18:0比例降低,表明L-蘋(píng)果酸抑制了硬脂酸C18:0向油酸C18:1n-9的轉(zhuǎn)化,同時(shí)抑制了棕櫚酸C16:0向棕櫚油酸C16:1n-9的轉(zhuǎn)化,導(dǎo)致線蟲(chóng)脂肪儲(chǔ)存減少。
(1)徹底性與相當(dāng)性相結(jié)合的原則。非法集資犯罪涉案財(cái)物的處置首先應(yīng)遵循徹底性原則,使被追繳的涉案財(cái)物數(shù)量最大化。同時(shí),應(yīng)綜合考慮涉案財(cái)物的利用方式、使用頻度、價(jià)值大小、與非法集資行為的關(guān)聯(lián)程度等因素,兼顧集資人、被集資人、利害關(guān)系人、國(guó)家等各方利益,防止涉案財(cái)產(chǎn)處置的范圍被不當(dāng)擴(kuò)大。
2.3 外源添加L-蘋(píng)果酸對(duì)線蟲(chóng)脂代謝通路基因表達(dá)的影響
1.3.8 數(shù)據(jù)處理及分析 使用GraphPad 8.4.0版本軟件對(duì)數(shù)據(jù)進(jìn)行作圖和統(tǒng)計(jì)學(xué)分析,
檢驗(yàn)進(jìn)行兩兩比較,單因素方差分析進(jìn)行組間比較,以
<0.05為差異有統(tǒng)計(jì)學(xué)意義。
1.3.5 甘油三酯檢測(cè) 用PBS緩沖液將攜帶1~3個(gè)蟲(chóng)卵的成蟲(chóng)從生長(zhǎng)板上洗下,收集于無(wú)菌的1.5 mL EP管中,清洗至肉眼無(wú)菌。超聲細(xì)胞破碎儀對(duì)線蟲(chóng)進(jìn)行破碎勻漿處理(冰水浴、超聲功率150 W、時(shí)間3 min),破碎勻漿后以2 000 r/min離心10 min,將上清液轉(zhuǎn)移至新的1.5 mL的EP管中待測(cè)。按照甘油三酯檢測(cè)試劑盒說(shuō)明,使用96孔板進(jìn)行檢測(cè),在空白孔里加入10 μL蒸餾水和1 000 μL的工作液,標(biāo)準(zhǔn)孔里加入校準(zhǔn)品10 μL和1 000 μL工作液,各個(gè)樣本孔中加入樣本10 μL和1 000 μL工作液,混勻后在37 ℃避光孵育10 min,酶標(biāo)儀,波長(zhǎng)510 nm,測(cè)定各孔OD值。每個(gè)組平行測(cè)定3次,取OD值得均值計(jì)算甘油三酯的含量。同時(shí)以BCA法測(cè)定各組樣本蛋白的濃度。
1.3.4 尼羅紅(NR)染色 100 mg NR粉溶于200 mL丙酮,避光攪拌2 h,-20 ℃冰箱密封儲(chǔ)存?zhèn)溆谩J褂们埃? μL儲(chǔ)備液溶于1 mL 40%異丙醇中制備為工作液。PBST(含0.01% Triton)將攜帶1~3個(gè)蟲(chóng)卵的成蟲(chóng)輕洗于1.5 mL離心管中,反復(fù)清洗3次至肉眼澄清無(wú)菌,以560 g離心1 min去除上清液。加入100 μL 40%異丙醇,并在室溫下震蕩孵育3 min,再以560 g離心1 min,去上清。加入600 μL的尼羅紅工作液混勻,室溫下避光旋轉(zhuǎn)孵育2 h。560 g離心1 min去上清,加入600 μL PBST進(jìn)行重懸,避光旋轉(zhuǎn)孵育30 min以去除多余NR染色劑,560 g離心1 min,留50 μL上清液重懸。取15 μL線蟲(chóng)懸浮液置于載玻片上,使用FITC/GFP通道,相同曝光時(shí)間進(jìn)行成像。實(shí)驗(yàn)重復(fù)3次,每個(gè)組拍攝不少于20條。
世界腕表不僅得到精英階層們爭(zhēng)相追捧,也深受各國(guó)政要們的喜愛(ài)。日內(nèi)瓦人民更是以世界時(shí)表作為禮物,饋贈(zèng)予那些在第二次世界大戰(zhàn)中為和平而戰(zhàn)的軍事領(lǐng)袖。這些藝術(shù)與科學(xué)融合的作品,得到了領(lǐng)導(dǎo)人們極高的贊譽(yù)。
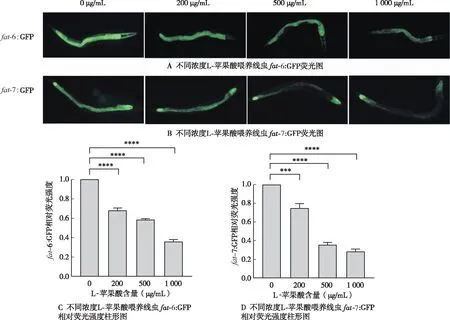
2.4 外源添加L-蘋(píng)果酸fat-6:GFP、fat-7:GFP線蟲(chóng)熒光強(qiáng)度的影響
確定L-蘋(píng)果酸降低
-5、
-6、
-7、
-1 mRNAs后,進(jìn)一步檢測(cè)
-6:GFP、
-7:GFP線蟲(chóng)熒光強(qiáng)度變化。與對(duì)照組(0 μg/mL)相比,200、500、1 000 μg/mL L-蘋(píng)果酸喂養(yǎng)線蟲(chóng),
-6:GFP、
-7:GFP熒光強(qiáng)度減弱(圖4A、4B),
-6:GFP熒光強(qiáng)度分別下降為對(duì)照組的0.68、0.58和0.36倍(
<0.05)(圖4C),
-7:GFP熒光強(qiáng)度分別下降為對(duì)照組的0.75、0.38和0.28倍(
<0.05)(圖4D)。進(jìn)一步證實(shí)L-蘋(píng)果酸抑制轉(zhuǎn)錄因子
-1表達(dá),進(jìn)而降低fat-5、fat-6、fat-7蛋白表達(dá),抑制硬脂酸C18:0向油酸C18:1n-9的轉(zhuǎn)化,同時(shí)抑制了棕櫚酸C16:0向棕櫚油酸C16:1n-7的轉(zhuǎn)化,導(dǎo)致線蟲(chóng)脂肪儲(chǔ)存的減少。
3 結(jié)論
線蟲(chóng)基因組共包含3個(gè)SCD基因
-5、
-6和
-7
,其中
-5主要將棕櫚酸(C16:0)轉(zhuǎn)化為棕櫚油酸(C16:1n-7),而
-6和
-7均將硬脂酸(C18:0)轉(zhuǎn)化為油酸(C18:1n-9)
。
-1編碼了秀麗隱桿線蟲(chóng)的哺乳動(dòng)物SREBP同源基因,SREBP是脂肪酸、膽固醇和其他脂質(zhì)代謝的關(guān)鍵調(diào)控因子
。在線蟲(chóng)中,
-49與PPAR
和
-1(SREBP的同源基因)序列相似調(diào)控SCD基因
-5、
-6和
-7的表達(dá)。
-1的突變導(dǎo)致油酸(C18:1n-9)與硬脂酸(C18:0)的比例下降
。研究發(fā)現(xiàn),L-蘋(píng)果酸降低秀麗隱桿線蟲(chóng)體內(nèi)脂肪和甘油三酯,使得脂肪酸含量發(fā)生變化,C16:1n-7/C16:0和C18:1n-9/C18:0比例降低。其機(jī)制為L(zhǎng)-蘋(píng)果酸抑制轉(zhuǎn)錄因子
-1表達(dá),進(jìn)而降低
-5、
-6、
-7基因表達(dá),抑制硬脂酸、棕櫚酸向油酸和棕櫚油酸的轉(zhuǎn)化,導(dǎo)致線蟲(chóng)脂肪儲(chǔ)存的減少。
[1]梁靜,張文舉,王博.L-蘋(píng)果酸的生理功能及其在反芻動(dòng)物生產(chǎn)應(yīng)用中的研究進(jìn)展[J].中國(guó)畜牧獸醫(yī),2016,43(7):1916-1921.
[2]Farney Tyler M,Bliss Matthew V,Hearon Christopher M,et al.The effect of citrulline malate supplementation on muscle fatigue among healthy participants[J].Journal of Strength and Conditioning Research,2019,33(9):2464-2470.
[3]Wu J L,Wu Q P,Huang J M,et al.Effects of L-malate on physical stamina and activities of enzymes related to the malate-aspartate shuttle in liver of mice[J].Physiological Research,2007,56(2):213-220.
[4]Guo Hai-peng,Chen Hou-ming,Hong Chun-tao,et al. Exogenous malic acid alleviates cadmium toxicity in
through enhancing photosynthetic capacity and restraining ROS accumulation[J].Ecotoxicology and Environmental Safety,2017(141):119-128.
[5]Commisso M,Bianconiu M,Poletti S,et al.Metabolomic profiling and antioxidant activity of fruits representing diverse apple and pear cultivars[J].Biology,2021,10(5):380-380.
[6]Alavez S,Vantipalli M C,Zucker D,et al.Amyloid-binding compounds maintain protein homeostasis during ageing and extend lifespan[J].Nature,2011,472(7342):226-229.
[7]Chen Xiu-lai,Wang Yuan-cai,Dong Xiao-xiang,et al. Engineering rTCA pathway and C4-dicarboxylate transporter for L-malic acid production[J].Applied Microbiology and Biotechnology,2017,101(10):4041-4052.
[8]Shi X,Li J,Zou X,et al.Regulation of lipid droplet size and phospholipid composition by stearoyl-CoA desaturase[J].Journal of Lipid Research,2013,54(9):2504-2514.
[9]Jones K T,Ashrafi K.Caenorhabditis elegans as an emerging model for studying the basic biology of obesity[J].Disease Models and Mechanisms,2009,2(5-6):224-229.
[10]Eyleen J O’Rourke,Soukas AA,Carr CE,et al.C.elegans Major fats are stored in vesicles distinct from lysosome-related organelles[J].Cell Metabolism,2009,10(5):430-435.
[11]He Bao-shen,Zhang Jing-jing,Wang Yan-li,et al.
Identification of cytochrome b5 CYTB-5.1 and CYTB-5.2 in
;evidence for differential regulation of SCD[J].Biochimica et Biophysica Acta(BBA)- Molecular and Cell Biology of Lipids,2018,1863(3):235-246.
[12]Brock T J,Browse J,Watts J L.Fatty acid desaturation and the regulation of adiposity in
[J].Genetics,2007,176(2):865-875.
[13]Zhu Y,Bai J,Zhou Y,et al.Water-soluble and alkali-soluble polysaccharides from bitter melon inhibited lipid accumulation in HepG2 cells and
[J].International Journal of Biological Macromolecules,2021,166:155-165.
[14]Chun-han C,Xian-qi L H,Fu-Lan H,et al.N-γ-(l-Glutamyl)-l-Selenomethionine inhibits fat storage via the Stearoyl-CoA desaturases FAT-6 and FAT-7 and the selenoprotein TRXR-1 in
[J].Molecular Nutrition & Food Research,2019,63(4):e1800784.
[15]Kuan Y C,Hashidume T,Shibata T,et al.Heat shock protein 90 modulates lipid homeostasis by regulating the stability and function of sterol regulatory element-binding protein(SREBP)and SREBP cleavage-activating protein[J].The Journal of biological chemistry,2017,292(7):3016-3028.
[16]Machado M L,Arantes L P,Gubert P,et al.Ilex paraguariensis modulates fat metabolism in
through purinergic system(ADOR-1)and nuclear hormone receptor(NHR-49)pathways[J].PLOS ONE,2018,13(9):e0204023.
[17]Bin Liang,Kim F,Lisa K,et al.The role of nuclear receptor NHR-64 in fat storage regulation in
[J].PLOS ONE,2010,5(3):e9869.
[18]Yang F, Vought B,Satterlee J,et al.An ARC/Mediator subunit required for SREBP control of cholesterol and lipid homeostasis[J].Nature,2006,442(7103):700-704.
