重度冷應激下民豬背最長肌應答分子調控網絡分析
2022-09-23 04:06:26馬守正李忠秋王文濤張冬杰
中國農業大學學報 2022年9期
馬守正 汪 亮 李忠秋 王文濤 劉 娣, 張冬杰*
(1.東北農業大學 動物科學技術學院,哈爾濱 150038; 2.黑龍江省農業科學院 畜牧研究所,哈爾濱 150086)
豬是對環境溫度較為敏感的一種大型家養動物,尤其新生仔豬。現代化集約飼養模式的興起,雖然提供了一個穩定的環境,但也導致豬對溫度的敏感性提高。冷應激會對機體的神經、心血管和免疫系統造成損害,因此突降的災害性天氣會導致巨大的經濟損失。在人和小鼠上的研究顯示,骨骼肌和棕色脂肪組織是機體遭受冷應激后的主要產熱部位。骨骼肌在室溫環境下,它的產熱占比在20%左右,但在寒冷環境下,它的產熱占比可升到40%。它同時以戰栗性產熱和非戰栗性產熱兩種方式為機體提供熱量,這對棕色脂肪較少的大型哺乳動物或沒有棕色脂肪的鳥類和家豬格外重要。骨骼肌的戰栗性產熱主要是通過骨骼肌不隨意的節律性收縮,增加代謝率和產熱量。骨骼肌的非戰栗性產熱獨立于戰栗性產熱,目前已知線粒體質子泄漏和肌漿網鈣循環是其主要產熱機理。
1954年,Sellers等首次提出在寒冷環境下,小鼠的骨骼肌會通過電活動(Electrical acticity)產熱維持體溫。Davis在對狗的骨骼肌研究中發現,低溫環境會導致骨骼肌的耗氧量增加,切除肌肉內的神經不會影響肌肉產熱,提出肌肉的非戰栗性產熱可能是體液介導的。1997年,de Mei等研究發現,肌質網膜上的Ca-ATP酶(Sarcoplasmic reticulum Ca-ATPase,SERCA
)可介導Ca由細胞基質中泵入肌質網中儲存起來,使肌質網內Ca濃度比胞質中高出許多。當機體受到神經沖動或環境變化刺激后,Ca由濃度高的肌質網腔中釋放入胞質中,在這一過程中,部分Ca與ATP的合成相耦合,還有一部分Ca泄露,偏離ATP合成,并導致熱量散失,被認為是肌肉產熱的主要來源。其中起關鍵作用的SERCA酶受小分子蛋白受磷蛋白(Phospholamban,PLB)和肌質蛋白(Sarcolipin,SLN)調控,尤其是SLN
。2017年Bal等發現在輕度和重度冷適應期間,肌肉的非戰栗性產熱與棕色脂肪的代謝產熱同時被啟動,如果一條途徑被阻斷,另一條途徑會增加產熱以維持核心溫度,2個途徑彼此間存在相互作用。此外,肌質網上的線粒體串擾、線粒體生物合成的增加、解偶聯蛋白3(Uncoupling protein 3,UCP3)誘導產熱以及糖異生的關鍵酶-果糖-1,6-二磷酸酶(Fructose 1,6-bisphosphatase 2,FBP2)的變化等都會改變骨骼肌的非戰栗性產熱。由此可知,低溫誘導下的骨骼肌非戰栗性產熱機理非常復雜。但受實驗條件所限,目前對大型家養動物的相關研究報道很少。豬既是一種大型的模式動物,同時也是一種重要的經濟動物。研究低溫環境對豬骨骼肌產熱的影響,一方面可為人類的相關研究提供借鑒,另一方面也可為生豬健康養殖提供理論幫助。本研究所選擇的民豬是東北地區唯一一個受到國家級保護的地方豬種,因長期生活在較為寒冷的環境中,使其具有明顯的耐寒特性。在-30 ℃的低溫環境中,也不會出現任何凍傷。本研究通過對遭受重度冷應激后民豬背最長肌進行轉錄組測序,從全基因組水平上篩選和分析了受低溫誘導的基因及lncRNA的表達情況,旨在為豬的骨骼肌非戰栗性產熱研究提供理論依據。
1 材料與方法
1.1 試驗動物
本研究在黑龍江省農業科學院畜牧研究所民豬養殖場進行。2021年1月3日,將6頭6月齡體重相近的雌性民豬隨機分成2組,每組3頭豬。試驗開始前,6頭民豬全部在室內有供暖的豬舍內飼養,溫度控制在(18±2) ℃,試驗開始后,一組置于室外的半敞篷舍內飼養,溫度隨外界環境溫度變化而變化,白天氣溫平均-17 ℃,夜間氣溫平均-26 ℃,一組仍留在溫暖的舍內飼養,兩組均保證自由采食和飲水,共處理3 d。試驗結束后,屠宰6頭民豬,取100 mg背最長肌置于液氮中保存。
1.2 RNA的提取
采用酚/氯仿法提取RNA,使用NanoDrop 2000&8000微量分光光度計對所提取的RNA進行純度檢測,使用安捷倫2100 Bioanalyzer進行質量濃度與完整性檢測。
1.3 文庫構建和上機測序
每個樣品取1 μg總RNA作為起始量構建lncRNA文庫。首先使用Ribo-off rRNA Depletion試劑盒去除rRNA,向反應體系中加入 Fragmentation Buffer使RNA片段化成為短片段,再以片段后的RNA為模板,利用隨機引物進行逆轉錄,實現cDNA第一鏈合成,并加入2nd Strand Marking Buffer、2nd Strand/End Repair Enzyme Mix合成cDNA第二鏈,后經末端修復、加堿基A,加測序接頭,通過磁珠篩選回收目的片段,加UNG酶消化cDNA二鏈,并進行PCR擴增,最后通過磁珠純化回收目的片段,從而完成整個文庫制備工作。使用NovaSeq 6000 S4對構建好的文庫進行測序。
1.4 原始數據的處理
測序得到的原始下機序列,含有測序接頭序列以及低質量序列,為了保證信息分析數據的質量,需要對原始下機數據序列進行過濾,包括去除接頭污染的Reads,低質量的Reads,含N比例大于5%的Reads,以及與核糖體RNA匹配的Reads,得到高質量的Clean reads后再進行后續分析。
1.5 lncRNA的鑒定
lncRNA的篩選條件如下:轉錄本長度≥200 bp,外顯子個數≥2;計算每條轉錄本的reads覆蓋度,篩去所有樣本中均小于5的轉錄本;利用gffcompare(http:∥ccb.jhu.edu/software/stringtie/gff.shtml)同豬的注釋文件進行比較,篩除豬中已知的mRNA及其他非編碼RNA(rRNA,tRNA,snoRNA,snRNA等);根據比較結果中的class_code信息(“u”,“i”,“x”)篩選潛在的lincRNA、intronic lncRNA和anti-sense lncRNA。將篩選所得的lncRNA作為最終的候選lncRNA進行后續分析。綜合4種分析軟件進行是否具有編碼潛能的篩選,主要有CNCI分析、CPC分析、PFAM蛋白結構域分析和CPAT分析。這4種分析方法均判別為non-coding的轉錄本為最終的lncRNA數據集。
1.6 民豬背最長肌受冷應激誘導的基因表達量分析
使用FPKM定量估計基因表達值,采用DEseq2進行試驗組和對照組間差異表達基因分析,滿足|logFC|≥1和q
<0.05的差異基因被認為達到顯著水平。根據兩組間上下調基因情況繪制差異表達基因的火山圖。1.7 民豬背最長肌受冷應激誘導的基因功能分析
GO富集分析。針對GO數據庫中的二級條目,計算每個條目的基因數目,應用超幾何檢驗,找出與整個基因組背景相比,差異表達基因顯著富集的GO條目。對KEGG中每個Pathway應用超幾何檢驗進行富集分析,找出差異表達基因顯著性富集的Pathway。
1.8 民豬背最長肌受冷應激誘導的lncRNA與mRNA的聯合分析
對于差異表達的lncRNA的靶基因,分別進行Cis和Trans靶標分析,通過靶基因間接預測其功能。Cis作用靶基因預測主要是通過lncRNA的功能與其在基因組座位上臨近的蛋白編碼基因的相關性進行的。將lncRNA相鄰位置(上下游50 kb)的蛋白編碼基因篩選出來作為靶基因。Trans靶基因預測主要是通過lncRNA的功能不依賴于和編碼基因的位置關系,而是與共表達的基因相關性進行的。根據lncRNA同mRNA的表達量相關性系數(斯皮爾曼相關系數,corr≥0.9)進行篩選。篩選后獲得的靶基因開展同mRNA相似的功能分析。根據識別出的差異表達lncRNA基因與mRNA基因及lncRNA的順式、反式靶標預測的基因的關系,繪制差異lncRNA與靶基因的調控網絡分析。
1.9 民豬背最長肌差異表達基因的Real-time PCR驗證
利用Roche實時熒光定量PCR儀LightCycler480Ⅱ對處理組存在顯著差異的高表達8個基因和2個lncRNA進行SYBR Green實時定量檢測,引物信息見表1。反應體系為:cDNA樣品0.5 μL,2×SYBR Green PCR Mixture 10 μL,特異性引物上下游各0.5 μL,滅菌水補充至20 μL。反應程序為:95 ℃ 10 min;95 ℃ 15 s,60 ℃ 30 s,72 ℃ 30 s,40個循環。檢測結果根據2法計算各模板中目的基因相對于內參基因β
-actin
的表達量。2 結果與分析
2.1 民豬遭受冷應激后背最長肌轉錄組測序結果
原始下機數據經過濾處理后,每個樣品平均獲得98 M的Clean reads,與基因組的比對效率為96.12%,其中,38.94%的序列比對到外顯子區域,30.50%的序列比對到內含子區域,30.56%的序列比對到基因間區, 篩選出的lncRNA經CNCI分析、CPC分析、PFAM蛋白結構域分析和CPAT分析后,取四者間的交集,共獲得10 220個lncRNA。
表1 qRT-PCR檢測用引物信息
Table 1 Primer information for qRT-PCR
基因或長鏈非編碼RNAGene or lncRNA引物序列(5'-3')Primer sequence (5'-3')SRXN11F: TCCACTCGGGTTGTATCGC1R: TGTCCACGAGGCTCTGCACSF12F: TTGCCTTTGAGTTTGTAGACC2R: TTGGCATTGGGAGTGTTGGPRC5A3F: GTGGATTGCCCTGCTCTT3R: ATGGTGTTCCGTTGCTTGIL1R14F: CGGGTCCACCTCTAACTC4R: GTAGTCGTCCCTGCCACACD2095F: TCTTCGTCTCATTGGGTTTC5R: TGGGTCTCCTGCTGGTCTARID5B7F: GGGCAACCCAGGTATCAT7R: TCATCCCTCGCAATCAGTMMP198F: CCCCAAGAGGCTGAATAG8R: GGTAGCTGCTGAAGTCCATITGA59F: CTCGCCAGGCTAGTTCCA9R: AAGGATGGTGACATAACCGTAGlncRNA13646010F: AGGAGGGAACAGATGGATTG10R: GTGAGGGCTGGCTTGTATTTlncRNA7596311F: TAGACAGAGGCTCCCTTCAT11R: CTGGGTTGCACAACTTCCβ-actin12F: CGGGACCTGACCGACTACCT12R: GGGCCGTGA TCTCCTTCTG
2.2 民豬遭受冷應激后背最長肌差異表達基因的篩選與分析
按照|logFC|≥1和q
<0.05的條件篩選后,民豬在遭受急性重度冷應激后,背最長肌共有88個基因表達發生了顯著變化,其中53個基因表達發生了顯著上調,35個基因表達發生了顯著下調。91個lncRNA表達發生了顯著變化,其中53個lncRNA表達發生了顯著上調,38個lncRNA表達發生了顯著下調。由于豬的lncRNA數據庫還不完善,因此這91個lncRNA均顯示為novel lncRNA。表達發生顯著變化的前10個基因列表見表2。上調變化倍數最大的基因是肝細胞生長因子激活因子(Hepatocyte growth factor activator,HGFAC
),上調了9.55倍;下調變化倍數最大的基因是多亮氨酸重復區和跨膜域1(Leucine rich rRepeats and transmembrane domains 1,LRTM1
),下調了7.04倍。2.3 民豬遭受冷應激后背最長肌差異表達基因的功能分析
對差異表達的基因進行GO功能注釋,發現在分子功能(Molecular function)中不存在顯著富集的條目,在生物過程(Biological process)中,有21個條目存在顯著富集,其中富集程度最高的是膠原分解代謝過程(Collagen catabolic process),在細胞組分(Cellular component)中,有5個條目存在顯著富集,其中4個是位于細胞外區間(圖1)。
表2 重度冷應激后民豬背最長肌表達發生顯著變化的前10個基因
Table 2 The top 10 genes with significant changes in the longissimus dorsi muscle of Min pigs after acute cold stress
基因Genelog2差異倍數log2 Fold change變化Change基因Genelog2差異倍數log2 Fold change變化ChangeHGFAC9.55UpLRTM1-7.04DownTREH5.59UpKCTD4-3.06DownFOSL14.87UpPCSK1N-2.82DownAREG4.53UpTMEM139-2.80DownMMP254.52UpCSRNP3-2.71DownHAS14.26UpLRRN3-2.63DownSLC11A13.72UpNCBP2-AS2-2.49DownTRIB33.67UpCOL8A2-2.41DownSLCO4A13.47UpENHO-2.25DownPRSS363.44UpNPPC-1.91Down

每個點的顏色表示該GO條目的富集程度。每個點的大小表示富集到該GO條目的基因的個數。用于表示富集程度的rich ratio,其計算公式為:(該通路的差異基因/所有的差異基因)/(注釋到該通路的基因/所有能被注釋到的基因)。 The abscissa represents the degree of enrichment, and the ordinate represents the GO term. The color of each dot indicates the degree of enrichment of that GO term. The size of each point represents the number of genes enriched for that GO term. The rich ratio used to express the degree of enrichment is calculated as (differential genes of this pathway/all differential genes)/(genes annotated to this pathway/all genes that can be annotated).圖1 重度冷應激下民豬背最長肌差異表達基因GO條目FDR值富集圖Fig.1 Enrichment map of Go terms FDR value of differentially expressed genes in longissimus dorsi of Min pig under acute cold stress
對差異表達的基因進行pathway富集分析,發現僅絲裂原活化蛋白激酶(MAPK)信號通路存在顯著富集,該通路中7個基因白細胞介素1受體1型(Interleukin 1 receptor type 1,IL1R1
)、雙調蛋白(Amphiregulin,AREG
)、絲裂原活化蛋白激酶8(Mitogen-activated protein Kinase 8,MAP3K8
)、70 ku熱休克蛋白1L(Heat Shock Protein Family A (Hsp70) Member 1 like,HSPA1L
)、集落刺激因子1(Colony stimulating factor 1,CSF1
)、RELB原癌基因NF-κβ亞基(RELB proto-oncogene, NF-kB subunit,RELB
)的表達發生了顯著上調,2個基因成纖維細胞生長因子受體4(Fibroblast growth factor receptor 4,FGFR4
)和p21活化激酶1(p21- activated Kinase 1,PAK1
)的表達發生了顯著下調。2.4 民豬遭受冷應激后背最長肌差異表達lncRNA的靶基因預測
lncRNA不編碼蛋白,主要通過順式(Cis)或反式(Trans)方式作用于蛋白編碼基因來實現。本研究中表達發生顯著變化的91個lncRNA順式調控129個靶基因,反式調控750個靶基因。通過構建lncRNA與mRNA之間的相互作用網絡(僅列出了發生顯著變化的前10個),發現兩者間的調控關系錯綜復雜(圖2)。比如lncRNA122198同時調控39個基因,使它們的轉錄發生了向上或向下的變化,而基因C11orf52
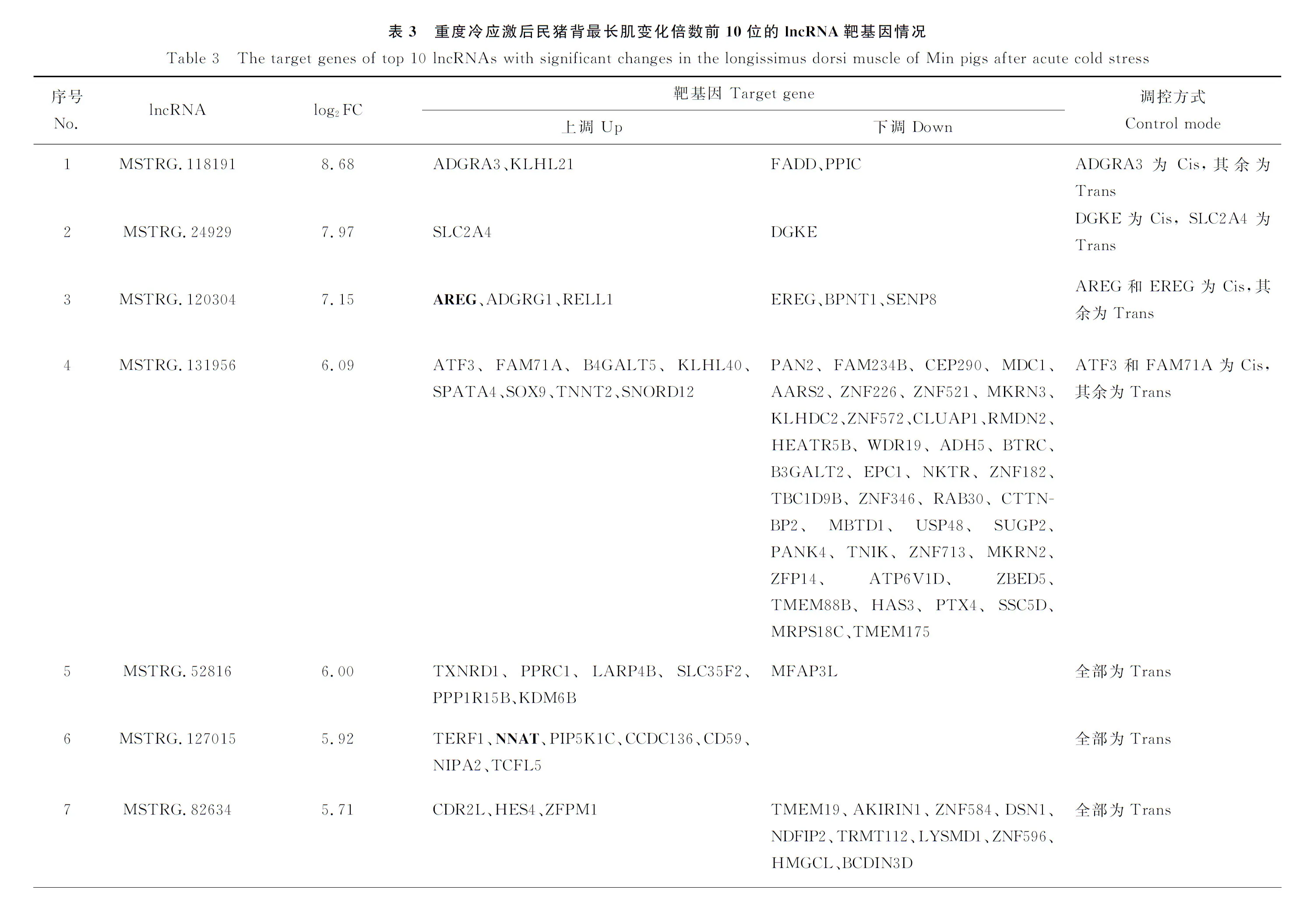
則同時受到9個lncRNA的調控。表3列出了表達發生顯著變化的前10個lncRNA及其靶基因表達變化和調控模式。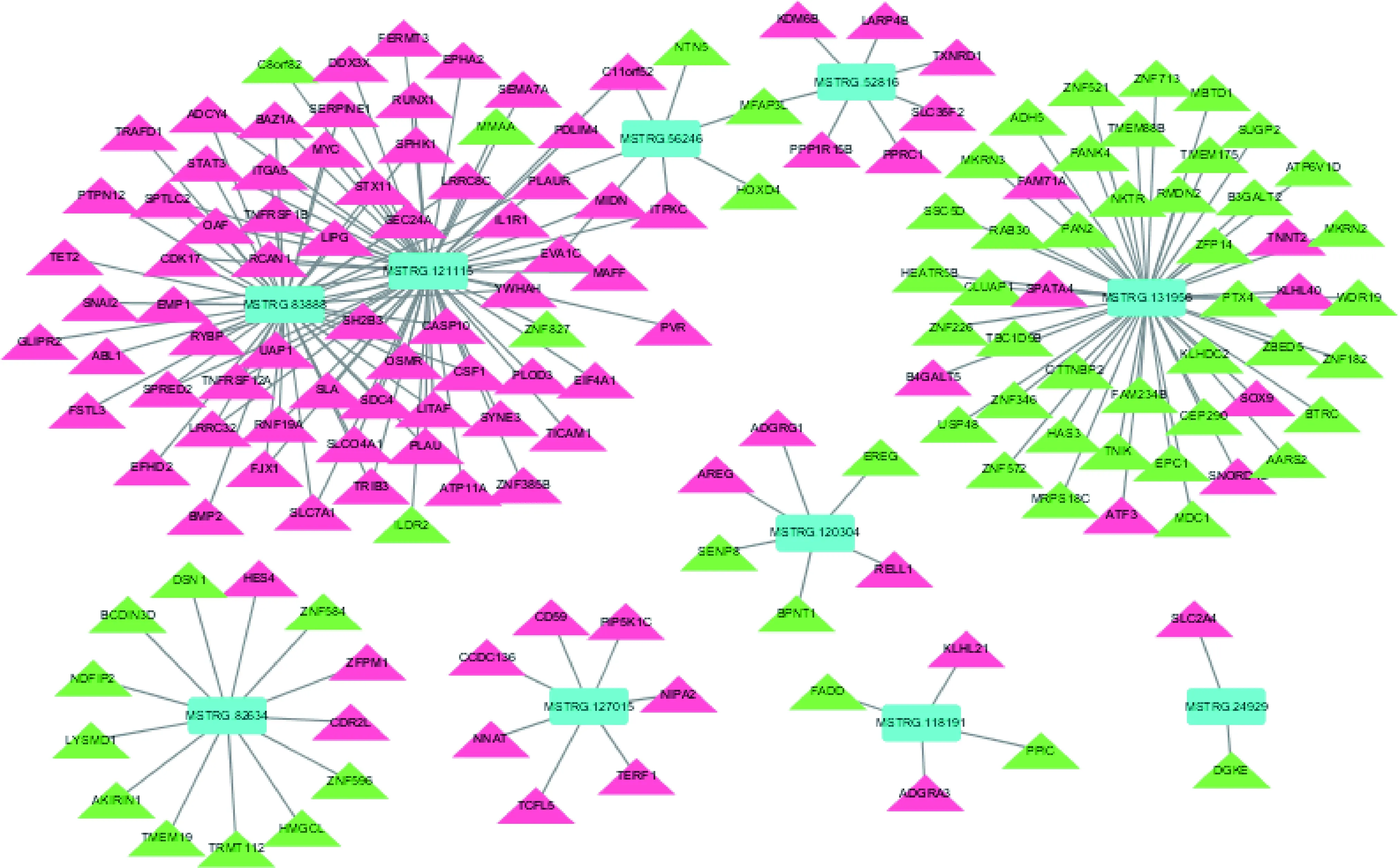
藍色代表lncRNA,紅色代表上調表達的基因,綠色代表下調表達的基因。 Blue represents lncRNA, red represents up-regulated genes, and green represents down-regulated genes.圖2 重度冷應激下民豬背最長肌差異表達lncRNAs和mRNAs之間的調控網絡圖Fig.2 Regulatory network between differentially expressed lncRNAs and mRNAs in the longissimus dorsi of Min pigs under acute cold stress
2.5 民豬遭受冷應激后背最長肌差異表達lncRNA的靶基因功能分析
對差異表達的lncRNA靶基因進行GO功能注釋,發現在分子功能中有19個條目存在顯著富集,其中磷酸轉移酶活性(Phosphotransferase activity)富集程度最高;在生物過程中,有232個條目存在顯著富集,其中富集程度最高的是單核細胞活化(Monocyte activation),在細胞組分中,有20個條目存在顯著富集,其中富集程度最高的在核質(Nucleoplasm)和核部分(Nuclear part),每個本體中富集程度前10的條目情況見圖3。

對差異表達的lncRNA靶基因進行pathway富集分析,發現MAPK信號通路(MAPK signaling pathway)和TNF信號通路(TNF signaling pathway)存在顯著富集,MAPK通路中有21個基因的表達發生了顯著上調,6個基因的表達發生了顯著下調。TNF通路中,有11個基因的表達發生了顯著上調,3個基因的表達發生了顯著下調。
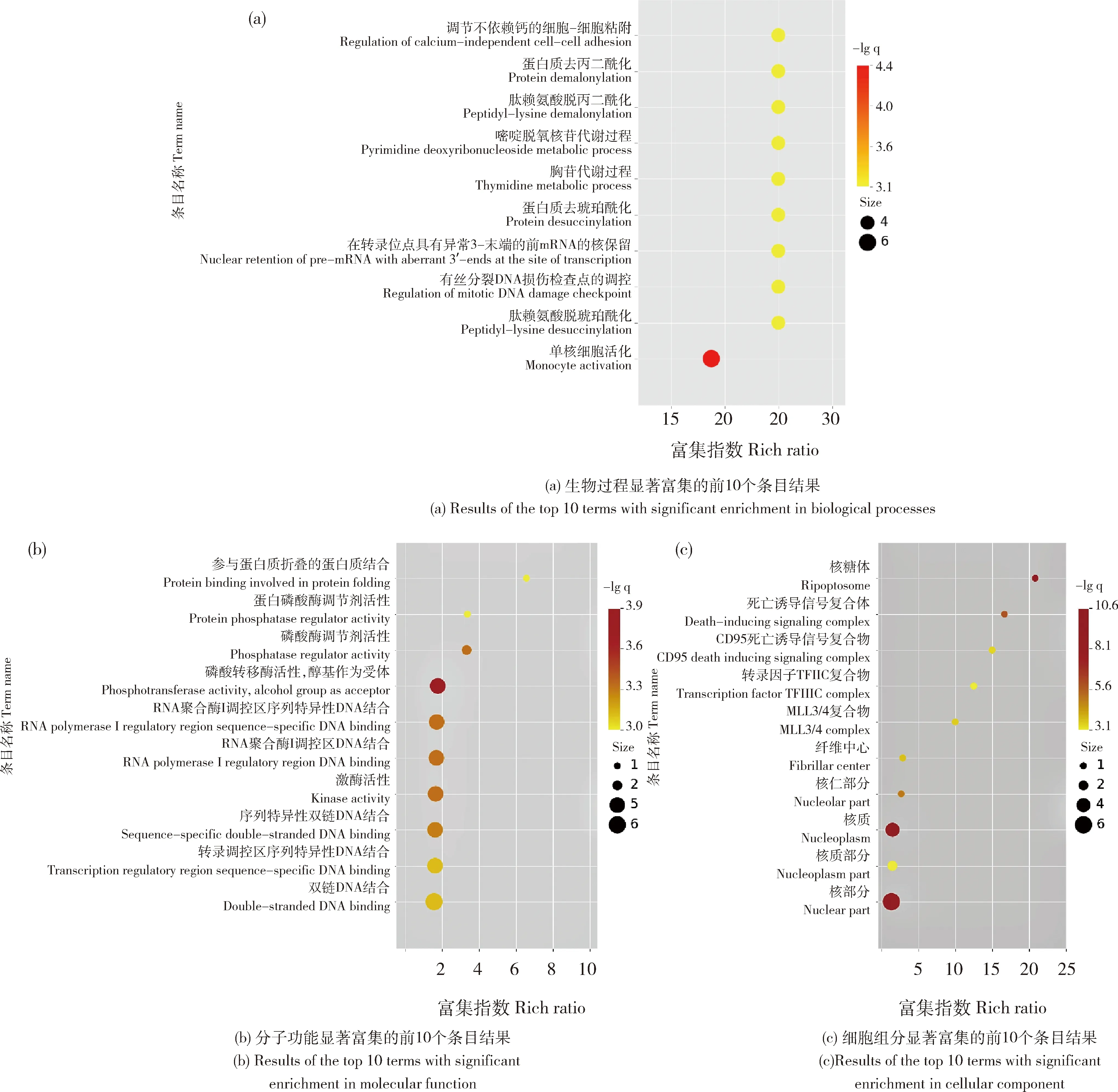
每個點的顏色表示該GO條目的富集程度。每個點的大小表示富集到該GO條目的基因的個數。用于表示富集程度的rich ratio,其計算公式為:(該通路的差異基因/所有的差異基因)/(注釋到該通路的基因/所有能被注釋到的基因)。 The abscissa represents the degree of enrichment, and the ordinate represents the GO term. The color of each dot indicates the degree of enrichment of that GO term. The size of each point represents the number of genes enriched for that GO term. The rich ratio used to express the degree of enrichment is calculated as (differential genes of this pathway/all differential genes)/(genes annotated to this pathway/all genes that can be annotated).圖3 重度冷應激下民豬背最長肌差異表達lncRNA的靶基因GO條目FDR值富集圖Fig.3 Enrichment map of GO terms FDR value of target genes of differentially expressed lncRNAs in the longissimus dorsi of Min pig under acute cold stress
2.6 測序結果的實時定量PCR驗證
為了驗證RNA測序結果的準確性,選擇了8個顯著上調表達的mRNA和2個顯著上調表達的lncRNA進行了 qRT-PCR驗證。結果顯示,qRT-PCR定量檢測值與測序值相比,變化趨勢基本一致(圖4),表明測序結果可信。

圖4 測序結果的qRT-PCR驗證Fig.4 qRT-PCR validation of sequencing results
3 討 論
戰栗是肌肉收縮的一個重復過程,是產生熱量的一個重要機制,是急性寒冷暴露的第一道防線。各種轉基因小鼠模型的研究表明,單靠戰栗不足以維持機體核心溫度(Core body temperature,Tc),必須激活非戰栗機制來維持熱量生產。對新生哺乳動物和成年嚙齒動物的研究表明,棕色脂肪組織(Brown adipose tissue,BAT)是非戰栗性產熱(Nonshivering thermogenesis,NST)的重要部位。因此,近二十年來對非戰栗性產熱的研究大多集中在棕色脂肪上。雖然BAT在許多哺乳動物中發揮重要作用,但它通常僅限于新生兒階段,在成年大型非冬眠哺乳動物(包括人類)中作用減弱。此外,還有一些恒溫動物,如鳥類、有袋類動物和豬,它們可以在沒有BAT的情況下依舊保持恒定的Tc,并在寒冷的氣候中正常生長。因此,有人提出骨骼肌是非戰栗性產熱的另一個重要部位。
二代測序技術的出現,使得研究人員可以在全轉錄組水平利用測序技術同時進行定量與定性分析,提高了分析的效率和準確性。本研究采用該技術對遭受重度冷應激后的民豬背最長肌進行分析,發現88個基因受到顯著影響(P
<0.05),其中HGFAC
上調倍數最大,HGFAC是活化單鏈前體肝細胞生長因子(HGF)成為有生物學功能的HGF的關鍵酶,在受損組織器官的修復和再生中發揮重要作用,HGFAC-HGF-PPARγ信號通路可參與調控全身的碳水化合物、葡萄糖和脂質代謝。它的上調表達,推測是由于肌肉組織受低溫影響發生了損傷。海藻糖酶基因(Trehalase,TREH
)在低溫刺激下也發生了5.59倍的上調表達。海藻糖酶是一種催化海藻糖水解的酶,在哺乳動物中鮮有報道,但在昆蟲和植物中已經明確其可受低溫脅迫影響,該酶在能量代謝、生長和逆境恢復中起著重要作用。其他的,如雙調蛋白基因(Amphiregulin,AREG
)、透明質酸合成酶1基因(Hyaluronan synthase 1,HAS1
)、Tribbles同源蛋白3基因(Tribbles pseudokinase 3,TRIB3
)等都發生了顯著的上調。AREG是一種細胞因子,參與抵抗由蠕蟲引起的感染免疫、免疫調節和傷口修復,是炎癥環境下肌成纖維細胞分化的重要驅動因素。HAS1是負責透明質酸合成的三種同工酶之一, 它的表達和活性由白細胞介素和細胞因子等促炎因子誘導,在許多炎癥反應中如骨關節炎、感染性肺病中,HAS1的表達都增高,表明它參與了炎癥反應。TRIB3屬于Tribbles信號調節蛋白家族,是關鍵的“壓力調節開關”,參與內質網應激反應、細胞生長分化等生物過程。此外,與先天性免疫相關的MMP25
和天然抗性相關的SLC11A1
也發生了顯著上調。被認為在非戰栗性產熱中起重要作用的SERCA1a
并未發生顯著變化,但其調節因子SLN
的表達水平下降了一半。通過對發生顯著上調的基因分析發現,急性重度冷應激下民豬的背最長肌發生了應激和炎癥反應。本課題組在前期研究中發現急性重度冷應激會造成民豬皮下脂肪與免疫、呼吸、氧化應激、血液循環和脂類代謝等相關基因的表達水平顯著上調,而與神經系統相關基因的表達水平發生顯著下調。小鼠骨骼肌中的催產素受體(Oxytocin receptor,Oxtr
)、瞬時感受器電位香草酸受體1 (Transient receptor potential vanilloid 1,TRPV1
)、冷誘導RNA結合蛋白(Cold-induced RNA-binding protein,CIRP
)、鋅α2糖蛋白(Zinc-α2-glycoprotein,ZAG
)等基因也會受到冷誘導而上調表達。本研究中篩選出的差異表達基因與前人的研究結果存在較大差異,推測這是由基因表達特性導致的。基因的表達具有顯著的時間特異性和空間特異性,受外界環境因素影響。因此種屬、日齡、組織、環境溫度以及作用時間等因素都會導致篩選出的基因存在差異。通過對差異表達基因的GO功能注釋分析和pathway富集分析后發現,這些差異表達基因所涉及的生物過程主要集中在膠原代謝、傷口愈合表皮細胞擴散和肌肉肥大調節過程等。這與人們在大鼠比目魚肌上發現的冰敷可促進膠原沉積增加、新生雛雞的急性冷暴露會導致心肌肥大等現象是一致的。差異表達基因在細胞組分(CC)的分析中發現,它們多位于細胞外區間,推測這可能與急性冷暴露下Ca、K等離子的轉運變化相關。重度冷刺激下,民豬肌肉組織中僅有1條生物學通路(MAPK信號通路)受到影響,說明低溫對民豬肌肉組織的正常生理功能影響較小。MAPK信號通路有三級的信號傳遞過程,可調節細胞的生長、分化、應激、炎癥反應等多種重要的生理/病理效應,是將環境刺激轉化為細胞反應的重要信號模塊。目前,在植物中已明確其對冷應激的應答反應機制,但在哺乳動物中還未見其對冷應激應答的報道。
lncRNA是長度大于 200 個核苷酸的非編碼 RNA,在劑量補償效應、表觀遺傳調控、細胞周期調控和細胞分化調控等眾多生命活動中發揮重要作用。民豬背最長肌遭受重度冷應激后,91個lncRNA發生了顯著變化,因豬的lncRNA數據注釋有限,所以這些lncRNA均顯示為novel lncRNA,它們主要以反式調控的方式作用于靶基因。在對靶基因的功能注釋中發現,許多基因與單核細胞激活作用、鈣非依賴性細胞間粘附的調節、蛋白質去甲基化、嘧啶脫氧核糖核苷代謝等生物過程相關。分子功能主要集中在醇基作為受體的磷酸轉移酶活性和RNA聚合酶Ⅱ調節區特異性 DNA 結合序列等。靶基因所富集的生物學通路,除了差異表達基因所富集的MAPK通路外,還有腫瘤壞死因子(TNF)信號通路。TNF是哺乳動物免疫反應中的一個關鍵傳遞者和調節者,它的主要生化反應是激活并調控NF-KB和AMPK途徑,在某些病理狀態下, 也能誘發細胞死亡。該通路的激活,也再次證明了冷應激下,骨骼肌內發生了炎癥反應。通過以上分析可知,冷應激影響了民豬背最長肌部分基因的表達,同時也影響了調控基因轉錄的部分lncRNA的表達。
本研究僅從全基因組水平探討了重度冷應激對民豬骨骼肌基因和lncRNA表達模式的影響,未來應從分子、細胞和個體水平深入研究冷應激對家豬能量代謝及其他方面的影響,通過培育耐寒品種、改善飼料營養等多種方式提高家豬抗寒性。
4 結 論
民豬在遭受3 d的重度冷應激后,背最長肌的88個基因受到顯著影響。與炎癥反應、低溫脅迫、應激反應和營養物質代謝相關的基因發生了顯著上調。差異表達基因所涉及的生物過程主要集中在膠原代謝、傷口愈合表皮細胞擴散和肌肉肥大調節過程等,且多位于細胞外區間。顯著富集的生物學通路僅MAPK通路1個。91個lncRNA的表達發生了顯著變化,這些lncRNA調控879個基因,且以反式調控為主。lncRNA調控的靶基因顯著富集到MAPK和TNF通路。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
電子制作(2018年18期)2018-11-14 01:48:24
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
山東工業技術(2016年15期)2016-12-01 05:31:22
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
山東女子學院學報(2014年6期)2014-03-01 02:24:55