辣椒果實螺旋性狀的組織學與轉錄組學分析
2022-09-23 04:32:50曹曉萌王怡心李子雄沈火林
中國農業大學學報 2022年9期
曹曉萌 王怡心 李子雄 沈火林 孫 亮
(中國農業大學 園藝學院/設施蔬菜生長發育調控北京市重點實驗室,北京 100193)
辣椒(Capsicum
annuum
L.)原產于南美洲,是一種重要的世界性蔬菜。我國是辣椒生產與消費大國,2019年辣椒播種面積達226萬hm,占全國蔬菜總播種面積的8%~10%,年產值約2 500億元。與許多園藝作物類似,辣椒果實的相關性狀,尤其是果形的變異十分豐富。不同地區消費者對不同果形的辣椒喜愛程度不同,例如,東北與華北地區消費者偏愛果形順直的牛角與羊角椒,華中、云、貴和川等地區的消費者更喜愛指形的朝天椒與細長形的線椒,而西北地區及越來越多的東部地區消費者則對螺絲椒青睞有加。螺絲椒因形似“螺絲”而得名,其果實呈長羊角形,基部有褶皺,整體呈右螺旋生長。與一般長羊角椒相比,螺絲椒的辣味更濃,皮更薄且肉更脆,因此深受消費者喜愛。螺絲椒起初在甘肅、青海、新疆和陜西等西北地區大規模種植,其在新疆鮮食辣椒種植面積中占比超85%;而后螺絲椒在我國東南部地區也逐漸發展起來,近年來在全國各地內均有大面積種植。雖然螺絲椒在辣椒產業中的地位逐年提高,然而有關其形成機理的研究卻相對較少。在自然界中,螺旋狀生長是一種常見發育模式,多見于植物的根、莖、纏繞的卷須、葉片以及呈螺旋狀排列的花瓣中。植物器官的螺旋生長對增強植物卷須的攀援能力、根系的侵徹力、種子的散播能力以及植物器官的機械強度等均有重要作用。在機理方面,有學者用植物力學模型與線性彈性理論來解釋多細胞器官徑向的扭曲,并認為這種螺旋生長主要源于幾何學與力學的約束。然而更多的研究發現,植物的螺旋生長與生理、亞細胞、細胞和分子層面的改變有關。在生理學方面,有研究發現植物器官不同的旋轉方向(左旋或右旋)是由南北半球的引力、磁場與植物激素共同作用的結果,北半球的植物左側生長素分布較多,生長較快,右側生長素分布較少,生長較慢,使植物向左彎曲;而南半球的情況恰好相反。在亞細胞與細胞層面,植物器官的螺旋生長被認為與細胞壁中的纖維素微纖絲以及細胞骨架的排布有直接關系,細胞的伸長和擴張受到細胞壁中纖維素微纖絲沉積方向的調控,通常來說纖維素微纖絲的排列方向與細胞伸長的方向垂直。然而,在螺旋生長的植物中,微纖絲往往呈斜螺旋狀排列,并且有研究表明細胞的縱向擴張與螺旋節距的增加有關。因此,在不增加纖維素微纖絲長度或減少細胞直徑的情況下,為了適應細胞長度的增加,細胞必然會扭曲。在植物細胞中,纖維素微纖絲的排列受細胞骨架尤其是微管排布的調控。纖維素微纖絲由纖維素合成酶A六聚體復合物(Hexameric CesA rosette)合成,而這一結構在纖維素合成互作蛋白(CELLULOSE SYNTHASE INTERACTING,CSI)與纖維素合成伴侶蛋白(COMPANION OF CELLULOSE SYNTHASE,CC)的輔助下沿微管移動。很多研究表明,微管排列方向的改變會直接影響纖維素微纖絲的排布方向。
很多植物的螺旋生長突變體也都與細胞骨架相關蛋白的突變有關。水稻中tid
1-1突變體的葉片和莖呈現右螺旋生長,TID
基因編碼α
-微管蛋白,tid1-1
突變是α
-微管蛋白第56個氨基酸由蘇氨酸突變為異亮氨酸所導致。擬南芥中與TID
高度同源的LEFTY
基因突變也會導致根系的螺旋生長,但其旋轉方向與水稻tid1-1
突變體相反,為左旋生長。lefty1
和lefty2
突變分別由微管蛋白α-tubulin 6和α-tubulin 4顯性突變所導致。同時,擬南芥右螺旋生長突變體tortifolia2
也是由α
-微管蛋白突變所導致的。除微管結構蛋白的突變外,一些微管相關蛋白(MAP)的突變也會導致植物的螺旋生長,例如擬南芥中CSI1、SPIRAL1、SPIRAL2和WAVE-DAMPENED蛋白的突變均會導致器官的右旋生長。其中,SPIRAL1是定位于微管的蛋白,過表達其編碼基因會增強微管對微管解聚藥物的抗性并促進下胚軸的伸長;SPIRAL2主要參與微管負端的穩定;WAVE-DAMPENED屬于TPX2-like家族,該家族成員均可與微管結合并參與細胞分裂、下胚軸伸長和維管束發育等過程。此外,番茄果形位點sun
也會導致器官的旋轉。sun
基因編碼IQ67家族蛋白-IQD12,在番茄中的過表達不但會引起果實的過度伸長,而且還會導致子葉與莖等器官的旋轉。擬南芥中研究發現很多IQ67家族成員均定位于微管并且與微管相關蛋白ROP以及KINESIN LIGHT CHAIN-RELATED互作。除上述MAP外,植物激素與重力等因素也會影響植物的螺旋生長。有研究表明激素可以通過改變周質微管的排列,促進或抑制微管的解聚,控制纖維素微纖絲在細胞壁上的排列方向和調控細胞的生長方式,引起植物的螺旋生長。在一些生長素相關基因的突變體中也發現了螺旋生長現象,例如aux1
突變體的根呈左螺旋生長,而rcn1
突變體的根則呈右螺旋卷曲。乙烯也被發現可以通過影響纖維素的定向沉積來改變微管的排列,并引起植物器官的螺旋生長。超重力也會造成植物的周質微管從橫向排列轉變為縱向排列并最終導致器官的旋轉生長。辣椒是為數不多的能在果實上觀察到螺旋生長的園藝作物,這種性狀與辣椒的經濟價值相關,然而目前人們對辣椒這一性狀形成機理的了解卻不夠深入。本研究以螺絲椒與牛角椒自交系為試材,利用徒手切片、石蠟切片和轉錄組測序等技術,旨在初步揭示辣椒螺旋果形的形成機理,為全面揭示植物器官形態建成理論奠定基礎。
1 材料與方法
1.1 試驗材料
試驗以本課題組選育的螺絲椒自交系19C705與牛角椒自交系19C961為試材。所有試材均于2021年春茬種植在中國農業大學上莊試驗站塑料大棚。
1.2 果實發育調查
于花后0、3、5、8和12 d觀察果實的旋轉情況,于破色期(花后38 d)用直尺測量果實的最大縱橫徑。以果實果肩至遠軸端最大垂直距離作為果實縱徑,以平行于果肩的果實最寬處作為果實橫徑。果形指數為果實縱徑除以橫徑所得值。每個試材選取至少5個植株,每個植株至少調查3個果實。
1.3 花期子房和破色期果實徒手切片制作與組織學調查
分別從19C705和19C961植株上選擇發育良好的破色期果實,切取果實中段1/3部分,分別做橫切、縱切與剖切,每個試驗材料至少調查5個果實,每個果實每個方向至少制作3個切片。取19C705和19C961的花期子房,去除萼片后分別制作橫切和縱切石蠟切片,每個試驗材料至少取8個子房,每個方向至少制作4張石蠟切片;組織薄片用甲苯胺藍染色后置于OLYMPUS DP72體式顯微鏡下觀察并拍照。使用Image J軟件測量橫向、縱向細胞的平均大小、細胞數目以及縱剖面細胞排列方向上的細胞形狀指數、細胞排列方向與果實縱軸的夾角。
1.4 統計學分析
上述試驗數據使用Microsoft Excel 2019與R軟件進行統計學分析。
1.5 轉錄組測序與分析
在19C705和19C961開花當天剝取子房并用液氮速凍,而后利用Invitrogen TRIzol試劑盒(Invitrogen,美國)提取子房RNA。轉錄組測序文庫構建與測序均在百邁客公司(青島)完成。轉錄組數據分析由百邁客云平臺(www.biocloud.net)完成,其中差異顯著基因利用DEseq2軟件包分析,錯誤率設置為FDR=0.05,差異倍數閾值設定為FC=2,平均表達量≥2 FPKM。差異基因富集分析(包括GO、COG、KEGG和KOG富集)和注釋分析(包括Pfam、Swiss prot、NR與eggNOG注釋)均在百邁客云平臺完成。篩選出重點基因的表達量使用R語言中Z
-score方法進行標準化,熱圖使用MeV4.9.0軟件進行繪制。1.6 實時定量熒光PCR
使用Prime Script RT Reagent Kit(寶生物工程(大連)有限公司)進行mRNA反轉錄,而后利用TB GreenPremix ExTaq
II(寶生物工程(大連)有限公司)試劑盒進行實時定量PCR反應,PCR儀為ABI PRISM 7500 Real-Time PCR System(Applied Biosystems,美國)。使用Primer 5軟件設計引物,詳細信息參見表1。選擇UBI-3
(Capana06
g002873
)基因作為內參基因,使用2計算相對表達量。2 結果與分析
2.1 破色期果實表型及組織學調查
19C705與19C961 2種試材果實大小與形狀差異見圖1(a)。19C705為典型的螺絲椒類型,果實較長、表面褶皺且整體呈右手螺旋;19C961為長牛角形,果面光滑。通過破色期果實徒手切片發現,在縱剖面方向上19C705的細胞呈傾斜排列且細胞形狀細長,而19C961的細胞整體呈垂直排列且細胞形狀較圓(圖1(c)和(d))。在橫切面方向上,2種材料的細胞均呈圓形且排列均垂直于表皮方向(圖1(e)和(f))。在縱切面方向上,19C705的細胞呈波浪狀排列,細胞大且細胞較多;而19C961細胞整體上呈縱向排列(圖1(g)和(h))。
2種試材花期子房的橫切與縱切切片見圖2。對2種材料花期子房進行測量與組織學分析發現,19C705的花期子房長度和寬度均顯著大于19C961,但二者子房的形狀指數差異不顯著(圖2和圖3(a));2種材料果肉縱、橫向細胞數目差異不顯著(圖3(b)),但19C705縱向細胞面積顯著大于19C961(圖3(c))。對2種材料破色期果實而言,19C705的果實長度顯著大于19C961,但果實寬度卻顯著小于后者,這也最終導致了19C705的果形指數顯著大于19C961(圖4(a));在細胞數目方面,19C705的縱向與橫向細胞數目均顯著多于19C961(圖4(b));在細胞大小方面,19C705的縱向與橫向切面細胞面積均顯著小于19C961,但是前者的縱剖面細胞面積卻顯著大于后者(圖4(c))。為更加準確地衡量果實縱剖面細胞的排列方式,本研究測量了縱剖面細胞排列方向與縱軸的夾角,結果發現19C705的夾角顯著大于19C961(圖4(d))。在縱剖面上,19C705的細胞形狀指數(細胞長度/寬度)也明顯高于19C961(圖4(d))。
表1 實時定量引物
Table 1 Primers for real-time qPCR
基因IDGene ID引物序列(5'-3')Primer sequence (5'-3')產物長度/bpProduct length退火溫度/℃TmCapana09 g000310_OFP8AGAGAATTGCAACTTCAACACCATGACCCCGAGAGAAGAATATTCCGATGAATC22064.865.1Capana01 g002642_KIN12ATCACCATCTCAGCTATCTCATCTGCTCCAGAGATTTATTTGATGCAGAATGGCAG18065.466.8Capana11 g001424_CycB1CATCATCTTCCCAACCAACAAGTATCTCAGGTAGCGGTCAATAATGTTGATTCG17965.765.1Capana03 g000798_ExpA1CGAGTTTCAAATGTTGTTGTCTTGGCTGACATAGGCATCCAGCCACTTCTAG19966.366.0Capana08 g000965_PGAGTAGCTGAGAACGTGTCAATGGCCCAGCTAGCAACTTGCAAGGTGAAGG18565.966.7Capana03 g002435_GA20ox1TGCATAGAGCAGTGGTAAACAACAAGACCTGAGTGAATTCATGGAGGGTAGGC16465.465.3Capana03 g003744_ZOG1AGCTGGTCGAGATCCGATTCTAATGACCGTGGAAGATAAACGACATGACG17865.266.4Capana12 g000825_ARF3CCCTTTGACAGAAGAAGAATGATCGAGCTACCAGCTCTTGTGAAGGCCTCTG24066.165.8
2.2 19C705果實發育調查
為明確19C705果實自何時開始發生旋轉,本研究對發育前期果實的旋轉情況進行了調查,結果發現花后8 d的果實已經有非常明顯的旋轉,并且在花后3和5 d都可以發現子房的不對稱生長(圖5(a)~(e)),說明旋轉出現的時期早于花后3 d。
2.3 轉錄組分析
對2種材料6個樣品進行測序共產生39.16 Gb的高質量數據,各樣品Q30均≥94.74%。在19C705與19C961間共鑒定出1 205個表達差異顯著基因,其中713個基因在19C705中被下調,492個基因被上調(圖6(a))。KOG分析發現上述差異顯著基因富集在從RNA處理與編輯(RNA processing and modification)到細胞周期調控-細胞分裂-染色體分割(Cell cycle control, cell division, chromosome partitioning)再到細胞壁-膜-包膜形成(Cell wall/membrane/envelope biogenesis)最后到細胞骨架(Cytoskeleton)的19個分類中(圖6(b))。另外,通過以往有關螺旋生長的研究,本研究在表達差異顯著基因中根據GO、COG、KOG、Pfam、Swiss prot、eggNOG和NR注釋,篩選出與細胞骨架、細胞周期與分類、細胞壁和激素相關的基因共45個(圖6(c)和(d))。其中,與細胞骨架相關基因中有3個驅動蛋白(Kinesin)基因(KIN
-UA
、KIN-14G
和KIN12A
)在19C705中被上調,1個Kinesin相關基因(KIN-14G
)被下調;1個驅動蛋白輕鏈相關蛋白(KLCR3
)基因在19C705中被下調;2個TPX2-like
基因被上調;1個成束阿拉伯半乳糖蛋白基因(FLA12
)與1個NPH3
基因被上調;另外,Flotillin-like3
(編碼筏蛋白)、Myosin-2-like
(編碼肌球蛋白)、OFP
8(編碼OVATE家族蛋白)與CML19
(編碼鈣調素相似蛋白)基因均在19C705中被下調,而ICR4
(編碼ROP持續激活抑制子)基因則被上調(圖6(c))。在與細胞周期和細胞分裂相關的基因中,僅有染色體結構維持蛋白基因(SMC1
)與Myb
基因在19C705中被上調,其他基因包括細胞周期蛋白基因CycA2
與2
個CycB1
、MEI2-like2
、細胞周期依賴激酶基因(CDK12/13
)、Myb113
基因、細胞分裂控制蛋白基因(CDC48C
)、絲氨酸/蘇氨酸激酶TOR
基因(mTOR
)和PIF
解旋酶基因(PIF1-like
helicase
),均在19C705中被下調(圖6(c))。在與細胞壁相關的基因中,植物細胞壁GP蛋白基因(GP1
)、WALL ARE THIN1(WAT1
)、擴展蛋白A1(ExpA1
)和β葡萄糖苷酶(BGLU42
)均在19C705中被上調,而果膠酯酶基因(PME53
)、多聚半乳糖醛酸酶基因(PG
)和另一個β
葡萄糖苷酶(BGLU42
)均在19C705中被下調(圖6(d))。在與植物激素相關的基因中生長素響應因子基因ARF3
、生長素受體基因TIR1
、吲哚-3-乙醛酸氧化酶基因AAO
、生長素響應蛋白SAUR32以及生長素極性運輸蛋白LAX3均在19C705中被下調,而生長素響應因子ARF18
和另一個生長素響應蛋白SAUR則在19C705中被上調;除生長素相關基因外,2個細胞分裂相關基因ZOG1
(玉米素-O-糖基轉移酶基因)均在19C705中被下調;赤霉素相關基因GA3ox2
、GA20ox1
與GA2ox8
的表達均在19C705中被上調,而赤霉素調節蛋白基因GASA6
則在19C705中被下調(圖6(d))。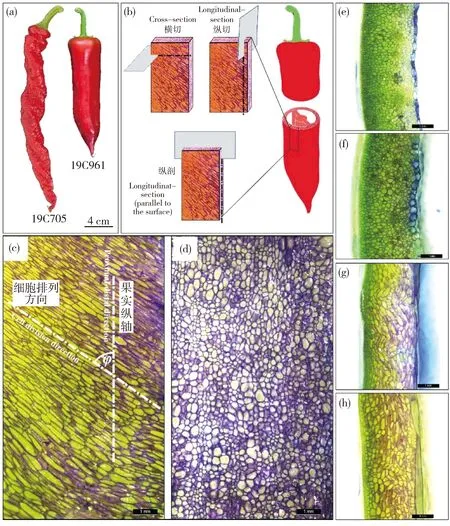
19C705和19C961分別代表試驗所用的螺絲椒與牛角椒。下同。(a)19C705與19C961成熟果實表型;(b)橫切、縱切與縱剖切示意圖;(c)和(d)分別為19C705和19C961縱剖切片圖;(e)和(f)分別為19C705和19C961橫切切片圖;(g)和(h)分別為19C705和19C961縱切切片圖。細胞排列方向與果實縱軸夾角為φ。圖(c)~(h)比例尺為1 mm。 19C705 and 19C961 respectively represent helical-shaped pepper and horn-shaped pepper used in the test. The same below. (a) 19C705 and 19C961 mature fruit phenotype; (b) Schematic diagram of cross cutting, longitudinal cutting and longitudinal cutting that parallel to the surface; (c) and (d) respectively represent 19C705 and 19C961 longitudinal cut slicediagram that parallel to the surface; (e) and (f) respectively represent 19C705 and 19C961 cross cut slice diagram; (g) and (h) respectively represent 19C705 and 19C961 longitudinal cut slice diagram. The angle between the cell arrangement direction and the longitudinal axis of the fruit is φ. The scale in Figs (c)-(h) is 1 mm.圖1 2種試材表型及徒手切片Fig.1 Phenotype and freehand section of two samples
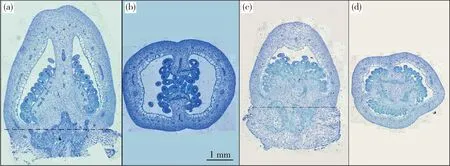
(a)19C705縱切石蠟切片圖;(b)19C705橫切石蠟切片圖;(c)19C961縱切石蠟切片圖;(d)19C961橫切石蠟切片圖。圖(a)和(c)中的虛線表示子房底端。
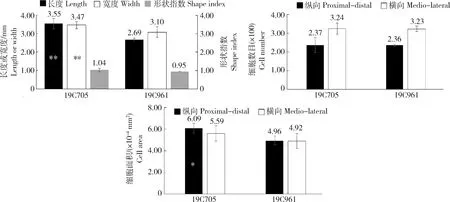
(a)花期子房長度、寬度與形狀指數;(b)花期子房縱切和橫切細胞總數;(c)花期子房縱切和橫切細胞面積。*表示0.05水平差異顯著;**表示0.01水平差異顯著。下同。

(a)果實長度、寬度與果形指數;(b)果實縱切與橫切面細胞數;(c)果實縱切、橫切與縱剖面細胞面積;(d)果實縱剖面細胞排列方向與縱軸夾角以及細胞在排列方向的細胞形狀指數。
2.4 基因表達的實時定量熒光PCR檢測
為了驗證轉錄組測序結果,本試驗挑選了8個基因進行了實時定量熒光PCR檢測(表2)。結果發現基因的表達模式與轉錄測序結果一致,其中OFP8
、CycB1
、PG
、ZOG1
和ARF3
分別在19C961花期子房中被上調了2.47、19.66、3.92、187.12和69.03倍;而ExpA1
和GA20ox1
分別在19C961被下調為19C705的0.38和0.51倍;對于KIN12A
基因而言,其在35個循環內沒有發生指數擴增,說明其表達量低于檢測范圍。3 討論與結論
3.1 2種試材果實形狀差異的組織學基礎

(a)~(e)為19C705試材花后0、3、5、8和12 d果實生長表型。
2種試材破色期果實在長度、果形、螺旋程度和表面平整度上均存在明顯差異。19C705的果實長度顯著大于19C961,而寬度顯著小于后者,這導致了其果形指數顯著高于19C961。在組織學層面,由于19C705的果實縱切面細胞大小顯著小于19C961,因此前者的果實長度主要受細胞數目的影響。而在果實寬度或周長方面,雖然19C705的橫向細胞數目顯著高于19C961,但增幅僅有9.74%,19C705果實的橫向細胞面積顯著小于19C961并且降幅高達57.45%。這種細胞數目與大小的相反變化趨勢最終導致了19C705果實寬度顯著小于19C961,同時也說明了橫向細胞面積是決定二者寬度的1個主要調控因子。綜合果實縱切、橫切、縱剖面細胞面積以及細胞形狀指數發現19C705的細胞較19C961更加細長。結合細胞排列方向與縱軸夾角φ
,可以通過S=S剖×(S縱×sinφ
)×1/2或S=S剖×(S橫×cosφ
)×1/2來粗略估算19C705的平均細胞大小,也可以通過S=(S剖×S橫×S縱)×1/2來粗略估算19C961的細胞平均大小。結果發現,19C705細胞的平均大小介于(10.41×10~11.83×10) mm之間,19C961細胞的平均大小約為11.88×10mm。這說明2種材料的整體果形與果實大小主要由細胞數目與細胞形狀共同決定。與上述情況不同的是,2種材料的花期子房在形狀指數上沒有明顯差異,說明二者果形指數的差異主要由果實發育階段差異決定。二者在子房大小與子房壁細胞大小上存在差異,可能暗示了開花前的發育過程影響了子房細胞的一些基本生長發育模式。在果實的螺旋生長方面,本試驗發現2種材料果實的差異主要體現在縱剖面細胞排列方向上,其中19C705的細胞基本呈直線排列,但排列方向與果實縱軸有明顯的夾角并呈右螺旋方向(果梗端到果頂端方向),而19C961細胞排列方向基本與果實縱軸平行。19C705這種細胞的排列方式與spr1
、spr2
、trn2
以及tor2
相似而與sun
和eb1b1
相反。由于19C705果實的螺旋并非由后期細胞分裂模式改變所導致,因此可以推測其果實螺旋生長的起始可能發生在子房發育較早階段。這種推測通過觀察果實連續發育結果得到了支持,因為在花后3 d即可觀察到果實的不對稱生長。另一方面,19C705果面的褶皺主要是由細胞在分裂方向上波浪狀排列所導致,然而在2種材料的花期子房切片中并沒有觀察到明顯的細胞排列變化,這種現象可能是由于子房細胞較小導致其排列變化幅度較小所導致,也可能是由螺旋生長的亞細胞基礎導致。前人發現,植物器官螺旋生長的原因之一是細胞壁纖維素微纖絲的傾斜螺旋排列,在這種情況下,植物細胞在膨大時會受到纖維素微纖絲長度與排布方式的約束,進而產生螺旋與細長化,并最終導致細胞形狀與分裂方向的改變使植物器官呈現螺旋生長。雖然這種纖維素微纖絲排布模式的改變可能發生在植物器官形成的早期,但是其組織學與形態學的明顯變化卻依賴于細胞的膨大,因此有可能在器官發育的早期階段觀察不到細胞形狀與排列方式的改變。3.2 2種試材果形差異的轉錄基礎

(a)所有差異表達基因熱圖;(b)差異表達基因的KOG富集;(c)細胞骨架、細胞周期以及與細胞分裂相關基因表達熱圖;(d)細胞壁與激素相關基因表達熱圖.(b)中方括號中的數字代表富集在對應功能的基因數目,百分數代表占比。
以往研究表明,植物器官的螺旋生長主要由細胞壁中纖維素微纖絲的排列模式調控,而纖維素微纖絲的排布模式又受到細胞中微管排布模式的調控。本研究在19C705和19C961花期子房轉錄組差異表達基因中篩選出與細胞骨架相關的14個基因,包括了驅動蛋白基因KIN
、驅動蛋白輕鏈相關蛋白基因KLCR3
基因、TPX2-like
家族基因、阿拉伯半乳糖蛋白基因FLA12
、NPH3
家族基因、筏蛋白基因Flotillin-like3
、肌球蛋白Myosin-2-like
、OVATE
家族蛋白基因OFP8
、鈣調素相似蛋白CML19與ROP持續激活抑制子基因ICR4
。其中KIN
、KLCR
、NPH3
和CML
基因家族成員的突變均被證明可以導致植物器官的螺旋生長。除上述基因外,ICR
4基因也被發現在2種材料子房中差異表達,該基因編碼1個ROP持續激活抑制子,而該抑制子被證明編碼植物特異Rho GTP酶并可以與IQ67蛋白結合。IQ67家族蛋白成員IQD12已被證明在高表達狀態下可以通過增加縱向細胞數和減少橫向細胞數使番茄果形伸長同時引起營養器官的螺旋生長。而IQ67蛋白也被證明可與KLCR
、CaM
以及CML
互作形成復合物并通過與微管的結合調控纖維素微纖絲的積累方向。因此,IQ67
相關通路可能在19C705中被激活并參與調控果實的螺旋生長與伸長。有趣的是,除IQ67
相關通路基因外,本研究還發現1個OFP家族成員基因OFP8
在19C705中被顯著下調。OFP蛋白家族成員已被證明在多種作物中調控器官的形態,例如在番茄中OVATE
基因的無義突變會通過改變果實基部細胞數目和伸長細胞形狀來使果實伸長并呈倒卵形。因此,19C705子房中OFP8
的表達下調可能也是導致該材料果實及果實中細胞伸長的原因之一。除細胞骨架相關基因外,一些與細胞壁相關的基因如GP1
、WAT1
和擴展蛋白ExpA1
均在19C705中被上調,而與細胞壁降解相關的果膠酯酶基因PME53
與多聚半乳糖醛酸酶基因PG
均在19C705中被下調,這也與子房細胞的分裂狀態相對應。除上述基因外,由于2種材料果實在細胞數目與形狀間也存在顯著差異,本研究也在差異表達基因中篩選出與細胞分裂及激素合成相關的基因。與細胞分裂相關的基因中除SMC1
與MYB
外,其余基因包括CYCLIN
、CDK
和CDC
等均在19C705中下調。這種情況與19C705細胞數目多于19C961的現象相反。出現這種現象的原因可能是19C705的細胞分裂高峰晚于19C961。植物激素尤其是生長素與赤霉素也被證明參與細胞數目與大小的調控。例如在番茄中改變ARF7
、ARF20
與miR160
的表達以及外施生長素均會導致果形的改變。然而在本試驗中除生長素響應基因SAUR
與ARF18
外,其他篩選出的基因包括與生長素合成、信號轉導與極性運輸相關基因均在19C705的子房中被下調。一方面說明SAUR
與ARF18
可能在19C705細胞分裂與伸長中發揮了重要作用,另一方面推測生長素在19C705與19C961果形的差異中并非發揮了最重要的作用。與之相反的是,2個細胞分裂素降解相關基因ZOG
均在19C705中被顯著下調,說明19C705子房中細胞分裂素的積累可能高于19C961,這可能是導致前者細胞數目多于后者的原因之一。同時,19C705子房中赤霉素合成相關基因GA20ox1
與赤霉素降解相關基因GA2ox8
與GA3ox2
的表達均被上調,說明19C705子房中赤霉素合成通路被激活。這種現象也可歸因于子房中細胞分裂素含量的提高,前人研究發現較高濃度的細胞分裂素可以誘導赤霉素的合成,植物體為了平衡體內提高的赤霉素含量又激活了赤霉素降解通路基因。表2 實時定量熒光PCR相對表達檢測結果
Table 2 Relative expression values of the interested genes in the real-time qPCR
基因IDGene ID19C705相對表達量19C705 relative expression level19C961相對表達量19C961 relative expression levelCapana09 g000310_OFP81.00±0.102.47±0.05Capana01 g002642_KIN12A1.00±0.03N.D.Capana11 g001424_CycB11.00±0.0819.66±3.04Capana03 g000798_ExpA11.00±0.040.38±0.08Capana08 g000965_PG1.00±0.063.92±0.07Capana03 g002435_GA20ox11.00±0.020.51±0.01Capana03 g003744_ZOG11.00±0.01187.12±40.11Capana12 g000825_ARF31.00±0.0569.03±1.43
注:N.D.表示在35個循環內沒有檢測到指數擴增。19C705和19C961分別代表試驗中所用的螺絲椒與牛角椒試材。
Note: N.D. indicates that exponential amplification is not detected within 35 cycles. 19C705 and 19C961 respectively represent helical-shaped pepper and horn-shaped pepper used in the test.
綜上所述,19C705果實可能通過改變KIN
、NPH3
以及IQ67
通路相關基因KLCR3
、CML
與ICR4
的表達來調控子房中細胞的初始分裂方向使果實呈現螺旋生長的表型,同時可能通過調控IQ67
通路相關基因、OFP8
以及細胞分裂相關基因的表達來增加縱向與橫向細胞數目并影響細胞形狀,最終使果實伸長。猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
小讀者(2021年2期)2021-03-29 05:03:48
華人時刊(2019年13期)2019-11-17 14:59:54
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
文苑(2018年22期)2018-11-19 02:54:14
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03