味覺受體研究熱點分析
2022-09-30 07:49:50張一純陳艷紅李利君
食品科學 2022年17期
關鍵詞:研究
張一純,陳艷紅,2,3,李利君,2,3,倪 輝,2,3,*
(1.集美大學海洋食品與生物工程學院,福建 廈門 361021;2.福建省食品微生物與酶工程重點實驗室,福建 廈門 361021;3.廈門市食品生物工程技術研究中心,福建 廈門 361021)
味覺是人們重要的感知反應,基本的味覺反應包括甜、酸、苦、咸、鮮5 種。近年來,一些科學家還發現了脂肪味和“加強味”的存在。錯綜復雜的味覺反應不僅能帶給人們愉悅和美妙的感覺,還可以用來評估食物中化學物質的營養及安全特性。鮮味和甜味分別代表著食物富含蛋白質和能量;苦味和酸味可以幫助識別有害或腐敗的食物;咸味有利于機體在飲食中保持電解質平衡。除此之外,味覺反應還與身體健康狀況密切相關,很多疾病的發生都伴隨著味覺感知的改變。
人們對味覺的感知依賴于味覺受體,食物風味成分和味覺受體相互作用,向機體發出信號,從而產生對食物的味覺響應。1999年人們發現了第一個味覺受體(taste receptor family 1 member 1,T1R1)基因,2001年美國科學家在人體第4對染色體中鑒別出一個與甜味感知相關的基因,2006年又報道人類基因組共編碼了25 種苦味受體T2Rs。近年來,隨著分子生物學、細胞生物學和神經生物學等技術手段突飛猛進的發展,人們對味覺受體的認知快速增長,味覺感知途徑、呈味物質和受體的相互作用機制研究越來越深入,味覺受體領域正逐漸成為研究熱點。
在Web of Science核心數據庫中以味覺受體為主題詞進行檢索共得到1980—2020年間7 973 篇文獻。本綜述根據這些文獻,對味覺受體熱點關鍵詞、期刊、來源、研究手段、受體類型、研究人員等進行分析概述,為開展味覺受體相關研究提供文獻綜述參考。
1 熱點關鍵詞及學術期刊分析
如圖1A所示,味覺受體領域發表文獻記錄數(number of records,Recs)最多的前20 位常用詞匯分別是苦味(bitter)、甜味(sweet)、酸味(acid)和鮮味(umami)等。來源包括大鼠(rats/rat)、人類(human)、小鼠(mice/mouse)和果蠅()。熱點研究手段主要體現在細胞(cell)、表達(expression)、蛋白質(protein)、誘導(induced)、基因(gene)、信號(signaling)、神經元(neurons)、分子(molecular)等常用詞匯。如圖1B所示,受體感知味道的種類、來源及研究手段的Recs占比分別為29%、26%和41%。這說明用細胞學和蛋白質表達技術對鼠類和人類的苦味受體和甜味受體進行研究是當前的熱點。文獻本地引用次數(total local cite score,TLCS)表示在被處理的所有文獻中的被引次數,圖2顯示了味覺受體領域發表文獻Recs和TLCS排名均在前7 位的期刊,分別是、、、、、,和,其中包括、、和等著名期刊,均為中科院分區2區以上期刊,通過閱讀這些期刊可以獲取味覺受體領域的前沿信息。
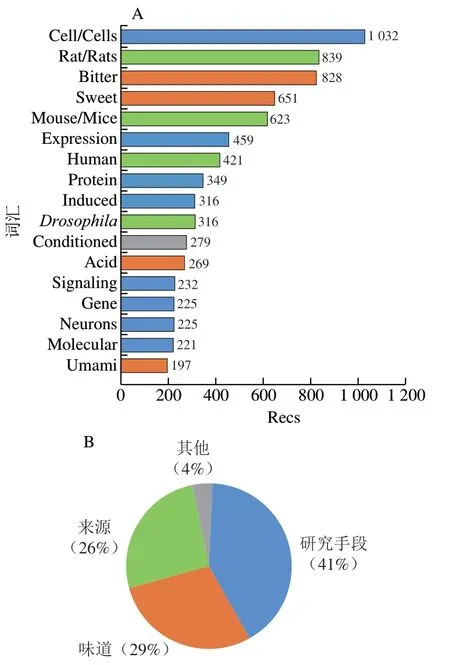
圖1 味覺受體領域文獻前20 位詞匯Recs(A)和歸類占比(B)Fig. 1 Top 20 lexical Recs (A) and their classification proportions (B)in the literature in the field of taste receptors
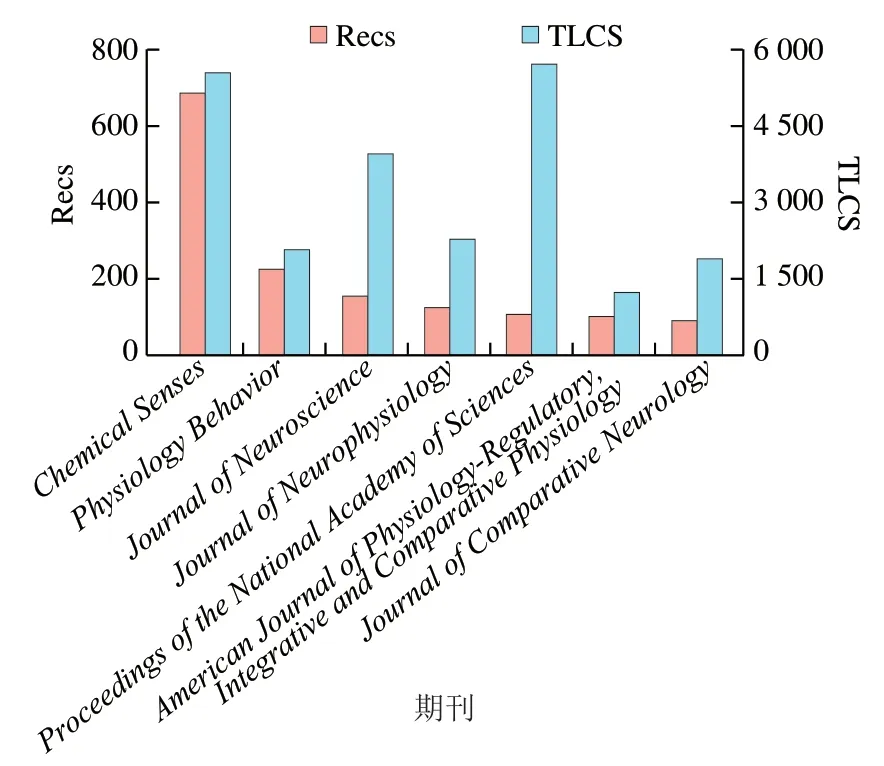
圖2 味覺受體領域的重要期刊Fig. 2 Important journals in the field of taste receptors
2 熱點來源分析
味覺受體來源的熱點詞匯分析表明,大鼠(rat/rats)和小鼠(mice/mouse)的味覺受體是科研工作中最常用的研究對象,其次是人類(human)和果蠅()。鼠類(包括大鼠和小鼠)和人類基因序列相似且操作簡單,是最常用的實驗材料;果蠅不僅和哺乳動物有相似的進食偏好,還可作為一個強大的遺傳模型,應用復雜的分子遺傳技術對其味覺的分子、生理和行為方面進行研究。科研人員利用鼠類味覺受體作為實驗對象模擬患者出現的生理癥狀,用于研發治療藥物。大鼠實驗證明,黃芪多糖可以改善高脂飲食損害的腸道甜味受體的信號傳導,通過調整2型糖尿病(diabetes mellitus type 2,T2DM)大鼠腸道中的葡萄糖轉運蛋白和甜味受體/胰高血糖素樣肽/胰高血糖素樣肽1受體途徑表達,從而減輕T2DM大鼠的癥狀。除此之外,通過鼠類實驗也驗證了味覺受體和其他激素之間也存在關聯性,例如唾液脂聯素可以影響胃中的味覺受體抑制饑餓素的分泌等。人類味覺受體的基因型常用于探索預防和治療肥胖等現代文明病的方法。例如在肥胖問題上,最新的一項研究表明肥胖與味蕾中基因表達量的改變有關,特別是肥胖病人中II型味覺細胞基因數減少;苦味受體T2R38 rs10246939與韓國人的飲食攝入量相關,具有TT基因型的女性會攝入更多的水果且肥胖風險也增加了1.75 倍。總的來說,味覺受體和肥胖具有關聯性,深入研究味覺受體對肥胖癥的預防和治療具有重要意義。最近的研究發現,果蠅的味覺器官分布在全身各處,并且除了味覺受體基因家族的成員,很多化學感受受體也都參與了味覺的感知,包括離子型受體(ionotropic receptor,Ir)、囊袋(pickpocket,ppk)和瞬時受體電位(transient receptor potential,Trp)基因家族編碼的受體,這些不同受體共同表達的模式可能是感知多種味道的基礎。近年來,科研人員對果蠅味覺系統與控制攝食、交配和產卵之間的聯系進行了研究,更加明確了味覺系統調節昆蟲行為的方式。總之,鼠類味覺受體易操作并且與人類受體相似,是大多數味覺受體研究的實驗對象。果蠅的味覺受體和哺乳動物的截然不同,通過分析果蠅的味覺受體可以為防治害蟲提供更好的策略。然而,不同來源的味覺受體之間仍然存在較大差異,例如人工甜味劑如阿斯巴甜、紐甜、甜蜜素、新橙皮苷二氫查耳酮和甜味蛋白等甜味物質均不會被鼠類動物感知。為了更準確探索人類味覺受體,還需要對人類本身的味覺受體進行深入的研究。
3 熱點研究手段分析
圖1顯示了8 個與味覺受體熱點研究手段相關的詞匯,其中表達(expression)、蛋白(protein)、誘導(induced)、基因(gene)和分子(molecular)屬于分子生物學和結構生物學的研究手段,細胞(cell/cells)屬于細胞生物學手段,除此之外,還包括了信號(signaling)和神經元(neurons)有關神經生物學研究手段。
目前,科研人員主要利用分子生物學手段使味覺受體表達并對其進行結構解析。哺乳動物表達系統可以使蛋白正確折疊并進行各種翻譯后修飾,保證味覺受體完整行使其功能。其中甜味受體和鮮味受體具有兩個亞基,要進行兩個亞基的共表達。此外,科研人員將味覺受體成功地在細菌表達系統中表達,大量生產重組蛋白。解析蛋白質等生物大分子及其配合物的三維結構信息的主流方法包括X射線衍射晶體學、核磁共振以及冷凍電子顯微鏡技術,這3 種方法各有優點,在高分子結構分析中具有重要意義。在這些技術中,X射線衍射晶體學仍然是最廣泛用于結構測定的技術。科研人員通過等溫滴定量熱法、熒光共振能量轉移和小角度X射線散射法分析了青鳉魚T1R2/T1R3,表明配體結合后異二聚體的構象發生轉化。接著通過捕蠅夾結構域(venus flytrap domain,VFTM)與抗原結合片段(fragment antigen binding,Fab)(由識別T1R2 VFTM的抗體制備)共結晶,又解析出分辨率為2.2~2.6 ?的青鳉魚T1R2/T1R3配體識別結構域,為理解味覺受體的化學感知提供了結構基礎。近年來,許多科學家通過對內分泌細胞構建模型進行體外研究,常用的方法包括熒光共聚焦顯微鏡、單細胞顯微切割捕獲系統、免疫印跡法等,發現了甜味受體激動劑可以分泌與食欲相關的激素,如胰高血糖素樣肽1、葡萄糖依賴性促胰島素激素和直腸黏膜酪酪肽,并且甜味受體也被認為與調節胰島素的分泌有關;我國上海交通大學近年來也發表了相關文獻闡述了葡萄糖代謝與甜味受體的表達有關;美國斯克利普斯研究所提出苦味受體的激活導致腸內分泌激素釋放,改善了代謝綜合征的癥狀,表明味覺受體與人類健康息息相關。
科研人員利用神經生物學主要對味覺受體的傳導過程及如何被感知進行了研究。當舌頭味蕾上的味覺受體接受到味覺刺激,信息通過顱神經VII、IX和X從舌傳遞到腦干。如圖3所示,依次通過孤束核(nucleus of solitary tract,NST)、腦橋臂旁核(parabrachial nucleus,Pbn)和腹后內側丘腦(parvicellular portion of the ventroposteromedial nucleus of the thalamus,VPMpc)的小細胞部分,然后傳遞給初級味覺皮層島皮質(insular cortex,IC)。研究表明,外周味覺系統中的味覺信息以標記線的方式編碼,因此每種味覺模式都有獨特的味覺受體和相應通路。其他研究者闡述了一種交叉纖維編碼的方式,即包括大量感覺傳入纖維的廣泛共激活,相同纖維的不同組合構成了不同味覺品質的編碼。電生理研究、雙光子鈣成像研究顯示,一些味覺神經元只負責一種味覺,而其他神經元對多種味覺刺激有廣泛的響應。東京大學重點研究了甜味信號的傳導過程,通過細胞消融、體內鈣成像和光磁實驗表征了Pbn中表達富含AT序列特異性結合蛋白2(special AT-rich sequencebinding protein 2,SATB2)的神經元在選擇性地將甜味信號傳遞到味覺丘腦中起重要作用;并對表達甜味受體突變體的所有細胞系進行了一系列細胞實驗,分析了與抑制劑的相互作用,基于這些結果,進行了對接模擬、能量最小化,闡明了人類甜味受體抑制劑的具體結構機制。Yoshida等通過熒光共聚焦顯微鏡鑒定味蕾細胞,再由高阻抗膜片鉗放大器記錄電信號,證明了苦味敏感的味覺細胞具有異質性,這種異質性可能有助于區分苦味化合物。Lu Bo等通過化學刺激和電生理信號記錄解釋了溫度對小鼠滋味感知的影響,其中溫度對甜味和鮮味刺激的響應伴有瞬時感受陽離子通道家族M成員5(transient receptor potential cation channel, subfamily M, member 5,TrpM5)的參與,而對鹽和酸的溫度調節響應并不是通過上皮細胞鈉離子通道和TrpM5的機制進行的,苦味刺激則受溫度的影響較小。在人類口腔中有5 000 個味蕾,它們都位于舌頭上表面、上顎和會厭,被分為I型(類膠質)細胞、II型(味覺受體)細胞、III型(突觸前)細胞、IV型(基底)細胞和V型(邊緣)細胞,I型、II型和III型細胞的協同作用是接受味覺的基礎。如圖4所示,I型細胞是味蕾中最豐富的細胞,由膠質細胞退化得到,頂端是鈉鉀離子泵,形成與鹽傳導有關的離子電流。對于II型細胞,鈣離子成像顯示其能夠響應甜味、苦味和鮮味刺激,能與多種有機分子結合,識別出甜、咸、苦等復雜分子造成的味覺。III型細胞能被水和氫離子激活,直接對酸味刺激和碳酸溶液作出響應。除此之外,味覺受體不僅在口腔表達,通過對其他組織細胞研究發現T1Rs和T2Rs廣泛表達于全身,分布于口腔外的受體和味蕾細胞中的受體信號通路相似,但無法感知味覺,而是具有營養傳感、氣道免疫防御和代謝過程調節等作用。嗅覺受體也同樣分布于全身,最近的一項實驗表明哺乳動物的味覺細胞上存在功能性嗅覺受體的表達。
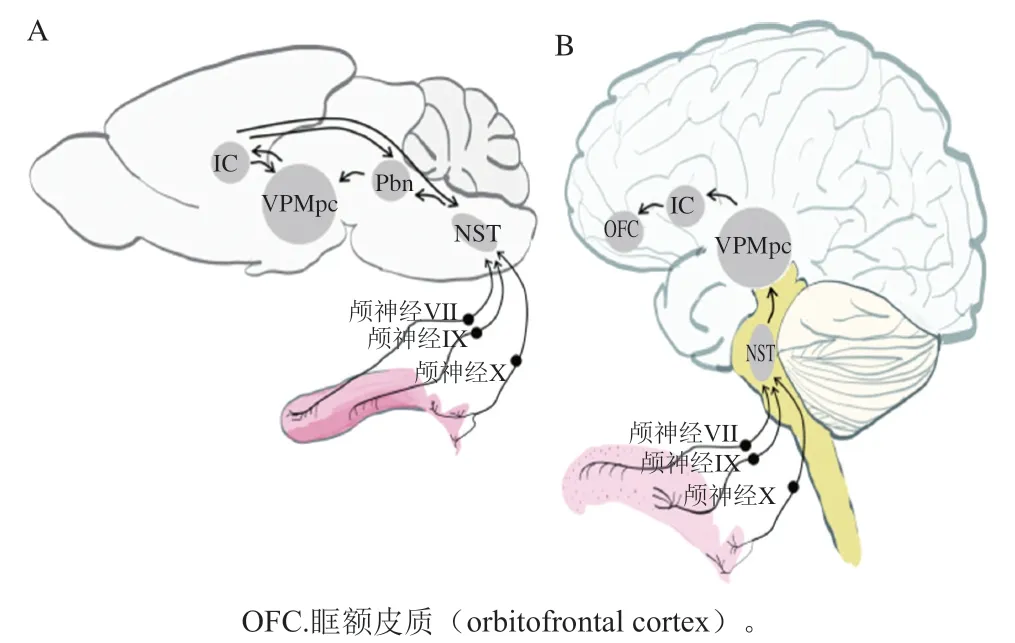
圖3 嚙齒動物(A)和哺乳動物(B)大腦感知甜味的味覺通路[32]Fig. 3 Taste pathways for sensing sweetness in rodent (A) and mammalian (B) brains[32]
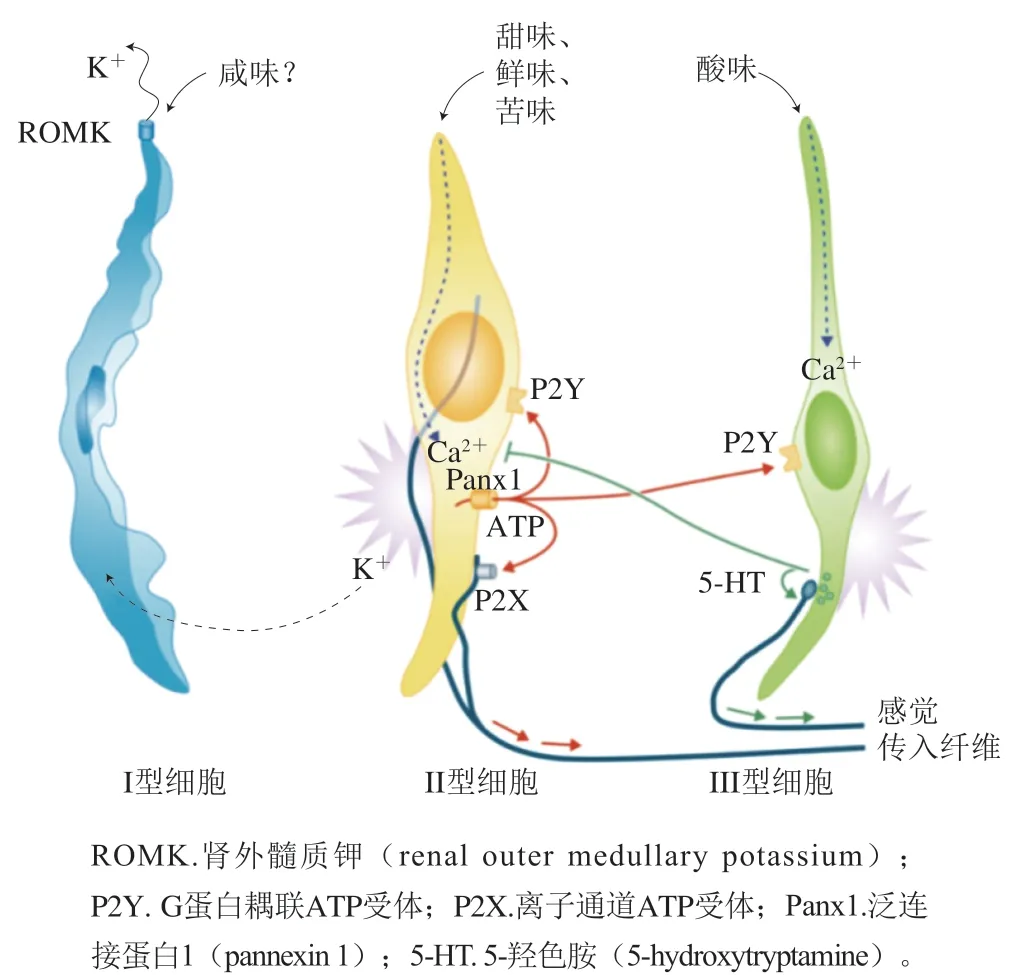
圖4 三大類味覺細胞[44]Fig. 4 Three major classes of taste cells[44]
近年來,科學家們基于味覺受體的表達及神經傳導領域的前沿技術,已經開始研發味覺受體方面的生物傳感器。中國科學院側重于研發體外生物電子舌來檢測苦味或鮮味。莫奈爾化學感官中心和我國浙江大學生物傳感器國家重點實驗室共同提出將人苦味受體T2R16在HEK-293細胞中作為生物傳感器的傳感元件,可對水楊酸進行特異性檢測。獲取高純度的蛋白質被認為是開發重復性強、性能穩定生物傳感器的關鍵;同時,氧化銦錫(indium tin oxide,ITO)作為苦味傳感器具有制備簡單、廉價和靈敏的優點。將基于大腸桿菌表達系統的味覺受體與ITO構建的電化學傳感器相結合,研發出的生物電子舌用于檢測特定苦味物質,該方法具有簡單和穩定的特點。此外,有些科研機構基于石墨烯效應晶體管構建雙鏈生物電子舌,通過將人的鮮味和甜味受體在HEK-293細胞中表達,利用細胞松弛素B將其制成納米囊泡,固定在微圖案化的石墨烯表面,可用于同時檢測鮮味和甜味。
總地來說,對味覺受體的探索已經進入分子生物學和神經生物學水平,但大多數受體的精確結構還未解析出,也難以區分不同特點的味覺神經刺激,目前主要采用計算機模擬和分子生物學實驗結合等手段對受體結構進行探索。
4 熱點受體研究進展分析
科研人員重點聚焦在苦味受體、甜味受體、酸味受體和鮮味受體上,其中苦味受體、甜味受體和鮮味受體均為G蛋白偶聯受體(G protein coupled receptors,GPCRs),酸味則由Otopetrin-1(OTOP1)質子通道感知。如圖5所示,GPCRs有相同的信號轉導通路,配體與GPCRs結合以后,GPCRs構象發生變化激活異源三聚體細胞內G蛋白復合物,G蛋白的-和-亞基從活化的-亞基上釋放,接著G蛋白的-和-亞基激活磷脂酶C-β2(phospholipase C-β2,PLCβ2)介導的肌醇1,4,5-三羥甲基氨基甲烷磷酸鹽(inositol 1,4,5-trihydroxymethyl aminomethane phosphate,PIP2)生成甘油二酯(diacylglycerol,DAG)和1,4,5-三磷酸肌醇(inositol 1,4,5-triphosphate,IP3),IP3作用于IP3受體(IP3 receptor,IP3R),使內質網釋放細胞內儲存的Ca,激活TrpM4/5通道,引發Na內流和細胞去極化,隨后細胞膜去極化激活電壓門控鈉通道(voltage-gated sodium channels,VGSC),最后激活鈣穩態調節器1/3(calcium homeostasis modulator 1/3,CALHM1/3)通道釋放ATP,導致位于感覺纖維中P2X2/P2X3的激活,觸發動作電位,向味覺皮層發出信號以進行感知。
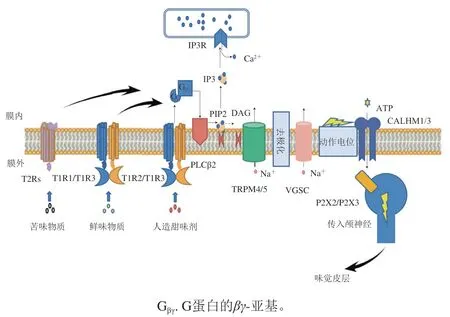
圖5 舌頭上的味覺受體細胞中甜味、鮮味、苦味的信號通路Fig. 5 Signaling pathways of sweet, umami, bitter in taste receptor cells in the tongue
苦味是5 種基本味道中最復雜的一種,苦味受體屬于GPCRs的A亞型,由味覺受體第2家族(T2Rs)所介導,以單體形式存在。與其他味覺GPCRs相比,TAS2R家族是最多樣化的并與多種激動劑結合。近年來,德國人類營養研究所發現了有助于感知二價錳離子或二價亞鐵離子鹽的苦味受體TAS2R7;根據同源建模和配體對接實驗對人T2R14進行點突變增強了對不同激動劑的選擇性。除此之外,該機構還提出人T2R14、T2R46在感知中藥的苦味上發揮了主導作用,并證明琥珀酸衍生化作用可使T2R14苦味受體對愈創木酚甘油醚的響應消失。現在普遍認為,苦味受體存在于呼吸道、腸道、生殖系統和泌尿道,不僅可以用于檢測有毒物質,而且可作為免疫“哨兵”防御致病性的侵略。賓夕法尼亞大學提出苦味受體T2R38功能的變化與慢性鼻竇炎的疾病狀態和疾病嚴重程度相關;T2R14作為上呼吸道感染的潛在治療靶點,其激動劑黃酮物質可能具有局部治療的臨床潛力。Ayabe等報道了啤酒中的異--酸和成熟啤酒花酸可能與腸道分泌細胞中的苦味受體結合,從而改善認知功能。此外,研究表明和基因可能與人類的長壽有關。對于苦味受體而言,有效的藥物開發很大程度上取決于對配體和受體結構-功能關系的研究,目前仍不清楚苦味受體的真實結構,主要通過同源建模、對接模擬、功能性實驗和突變體結合來獲得。除此之外,近年來我國的科學家也將目光投向了研究苦味受體和物種進化的關系上,中國科學院提出大熊貓和小熊貓由于飲食習慣造成了苦味基因的特異性進化,武漢大學則探究出功能性苦味受體的數量與禽類飲食中大量潛在毒素水平之間存在正相關。這些報道為研究鳥類和哺乳動物感覺進化的分子基礎以及對生態環境的適應過程提供了新思路。
甜味受體T1R2/T1R3和鮮味受體T1R1/T1R3屬于GPCRs的C亞型,味覺受體第1家族(T1Rs),以異二聚體形式存在,如圖6所示,T1R2/T1R3的結構包括VFTM、半胱氨酸富集區(cysteine rich domain,CRD)、跨膜區(transmembrane domain,TMD)和胞內結構域(intracellular domain,ID),T1R1/T1R3與之類似。甜味受體和鮮味受體兩者共享一個亞基T1R3,與T1R3結合的甜味劑——甜蜜素雖然不能激活鮮味受體,但能增強谷氨酸鹽誘導活性,是T1R1/R1R3的正變構調節劑,甜味抑制劑Lactisole則是T1R1/T1R3的負變構調節劑。除了人工甜味劑的傳導途徑外,甜味還涉及不同的信號傳導途徑(圖7)。蔗糖等糖類物質與T1R2/T1R3結合,使G蛋白的-亞基(G)活化,從而激活腺苷酸環化酶(adenylyl cyclase,AC),產生3’,5’-環腺苷酸(cyclic adenosine monophosphate,cAMP),導致胞內cAMP濃度上升,cAMP直接激活蛋白激酶A(protein kinase A,PKA),引起K通道的磷酸化,導致離子通道關閉,抑制K外流,引起膜去極化和神經遞質釋放。甜味受體已經被發現在大腦、胃腸道、腎臟和脂肪等組織中均有表達,受體不僅影響著代謝過程,如胰島素分泌、糖代謝和脂肪代謝,還與包括T2DM在內的慢性炎癥疾病有關。慢性代謝疾病患者具有較高的與飲食行為相關的等位基因頻率,他們對富含甜味的食物更加喜愛。隨著人們健康意識的提高和工作性質的轉換,低熱量、無熱量的甜味劑在食品領域得到廣泛使用。然而多數甜味劑甜味不純正,呈現速度和持續時間與蔗糖相比有差異,存在不良“異味”,導致產生不愉悅的口感。為了滿足消費者對減糖和口感的雙重需求,科研人員運用構建嵌合體、計算機模擬來探究甜味受體與配體相互作用從而設計新型甜味劑。鮮味受體方面的熱點聚焦在鮮味肽/鮮味增強肽的呈味規律和變構調節。除T1R1/T1R3外,目前報道的代謝型谷氨酸受體(metabotropic glutamate receptors,mGluRs)和細胞外鈣受體(extracellular calcium sensing receptor,CaSR)等也可以感知鮮味且均為GPCRs。不同的鮮味劑之間具有協同作用,例如在含谷氨酸的食物中添加游離核苷酸如5’-磷酸鳥苷后,鮮味會增加數倍。除此之外,鮮味劑還與其他味覺感知受體具有相互作用,例如鮮味肽通過人類苦味受體抑制苦味。2020年,Spaggiari等發表了一篇研究鮮味、甜味和苦味受體的計算機分子建模技術的完整概述,該方法已用于味覺感受器建模、模擬促味劑-受體結合方式以及篩選潛在促味劑等方面。近年來研究人員提出了幾種機器學習模型,用定量構效模型來預測分子的甜度。最新的一篇文獻報道了通過結構鑒定和分子對接從天然庫中篩選潛在的甜味劑。目前,主要通過建立可靠的突變實驗、構建嵌合體、數學建模、分子對接、電生理技術、動物味覺模型來深入研究有關受體分子的作用機制。然而甜味受體和鮮味受體的結構和微觀作用規律以及呈味規律還無法準確的闡明,因此,采用冷凍電子顯微鏡等技術解析甜味受體的精細結構是本領域的研究方向。
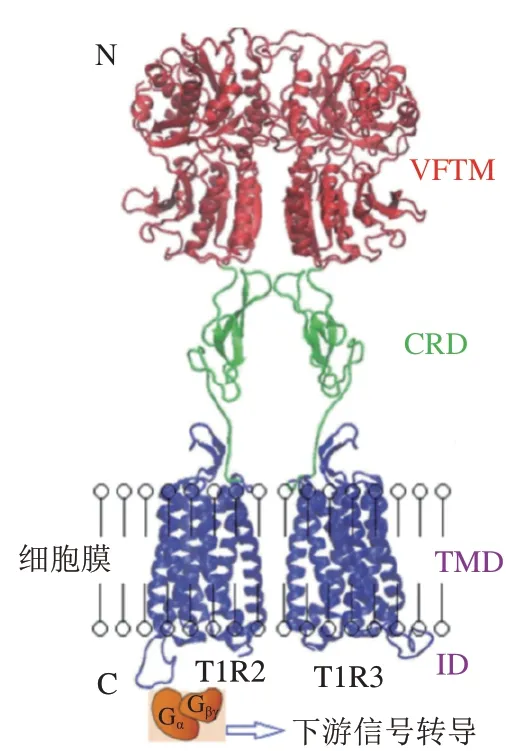
圖6 甜味受體T1R2/T1R3的結構示意圖[27]Fig. 6 Schematic diagram of sweet taste receptors[27]
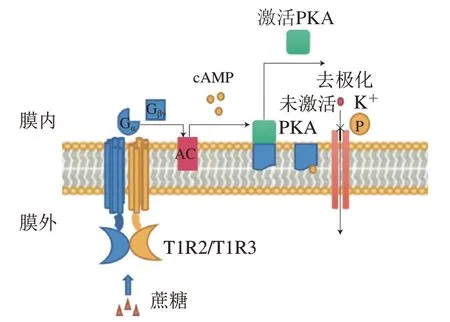
圖7 甜味受體感知蔗糖的信號傳導通路Fig. 7 Signaling pathways of sucrose sensing by sweet taste receptor
酸味一直被認為是5 種基本味覺之一,該味覺檢測途徑的發現歷經坎坷。早期,科學家提出了、、等候選基因可能與酸味感知有關,它們(特別是)在酸敏感離子通道或在酸敏感味覺受體細胞中富集表達。科學家一直致力于確認這種受體以及其他可能的酸味受體的作用機制和功能;并發現即使缺乏假定的酸味受體細胞,動物對酸仍然表現出強烈的厭惡,這為確定酸味受體的功能基因帶來了很多困難。因此,酸味受體的研究進展緩慢,直到2019年才發現了真正的酸味檢測通道OTOP1。近年來,科學家解析了斑馬魚OTOP1的結構,闡明了其轉導機制,發現強酸和弱酸的傳導機制完全不同,強酸的H通過對Zn敏感的H通道進入細胞,而弱酸以未解離的中性分子進入細胞再解離形成H。如圖8所示,H進入細胞引起細胞內的酸化,阻斷了K2.1 K通道,細胞膜去極化,電壓門控Na通道開放,引起一系列動作電位開放電壓門控Ca通道,神經遞質產生神經沖動引起酸味的感知。目前在破解傳入大腦途中處理信息的神經回路方面已經取得了一些進展,但其傳導通路中仍有許多環節有待研究。
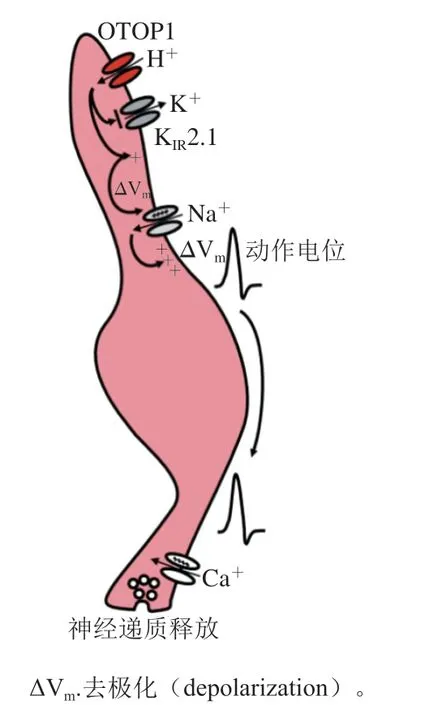
圖8 酸味受體的信號傳導通路[105]Fig. 8 Signaling pathways of sour taste receptors[105]
總而言之,味覺受體的結構特征和相關機制都尚未明確,該領域還需要更多的科學支持和技術支持。新型滋味物質的設計、味覺的感知機制、受體和健康的關系及個體感知的差異方面將不斷吸引著科學家的關注,是未來的探索方向。
5 熱點科研機構及科學家分析
如圖9所示,Recs排在前5 名的國家分別是美國、日本、德國、英國和中國;其中美國的Recs占全部Recs的44%,遙遙領先其他國家。TLCS排名前10的科研機構中有9 個來自于美國(圖10)。其中加州大學圣迭戈分校發表的文獻被引次數最高,該機構很早就致力于研究哺乳動物口腔感知鮮味、甜味和苦味的機制,發現了第一個味覺受體T1R1,并利用外源表達證明了味覺蛋白T1R2和T1R3結合感知甜味、苦味的編碼獨立于甜味和鮮味等基礎內容。莫奈爾化學感官中心提出苦味受體TS2R38的多態性編碼對包含N—C=S片段的苯硫脲和丙硫氧嘧啶化合物表現出不同的響應程度,這種多態性可能來源于進化壓力;苦味受體不僅可以防止有害食品的攝入,T2R38還在上呼吸道中表達并發揮“哨兵”的功能;該機構還提出甜味、苦味和鮮味傳入大腦的方式是通過CALHM1離子通道釋放ATP,最終激活傳入神經通路。美國國立衛生研究院牙科與頜面研究中心發表的文章對味覺受體研究有著深遠的影響,推動了味覺受體領域的發展,該中心的研究團隊很早就開始研究哺乳動物味覺受體的結構,相關文獻均發表在1999—2018年,2002年其發現了甜味和鮮味受體有一個共同的亞基;2004年發現了在甜味受體上存在多種配體結合位點,證明了T1R3和T1R2的不同功能。科羅拉多大學主要致力于研究味覺傳導機制,發現味覺傳導過程包括ATP信號傳導和鈣離子傳導,同時發現II型細胞是苦味、甜味和鮮味味覺轉導的主要轉導者,可能通過光滑內質網下池與神經系統通信,以取代傳統的突觸。邁阿密大學提出受體細胞在接受味覺刺激后通過Panx1分泌ATP,并進一步刺激其他味覺細胞釋放血清素,這一發現闡明了味蕾細胞間信號處理的方式,在味覺受體信號傳導方面作出了巨大貢獻;2015年該機構還提出脂肪組織產生的瘦素能夠特異性作用于甜味,對苦味誘發的遞質釋放沒有影響。
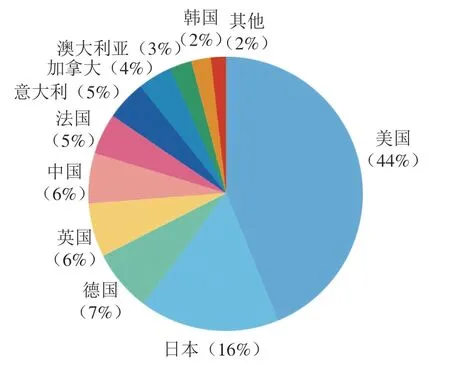
圖9 以發文量Recs為指標排名前10的國家/地區及Recs比例Fig. 9 Top 10 countries/regions by the number of publications and their percentages in total Recs
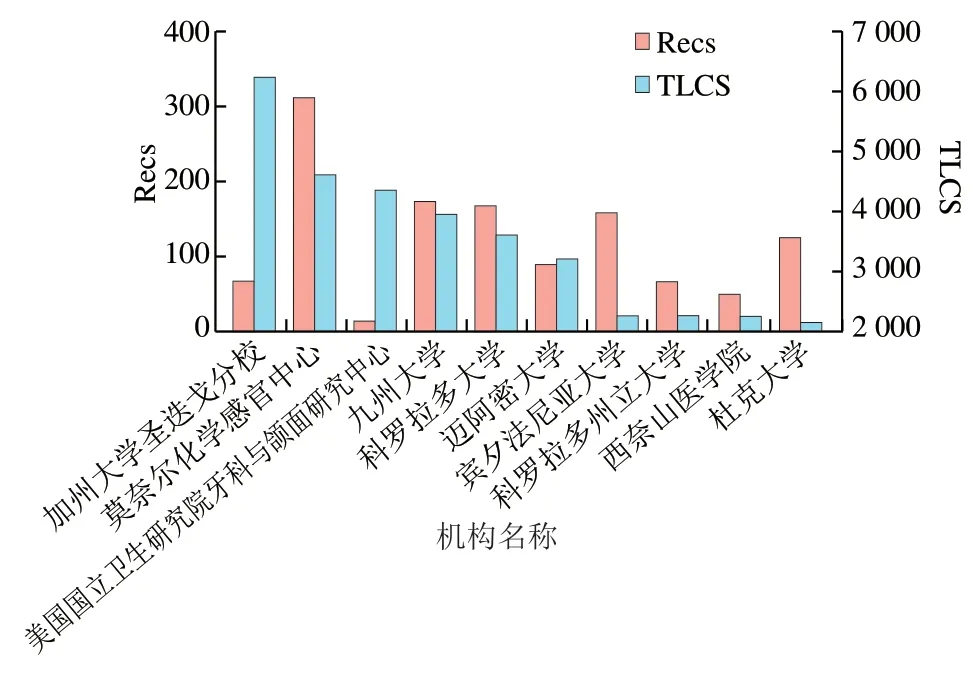
圖10 以文獻TLCS為指標排名前10的研究機構Fig. 10 Top 10 research institutions by literature TLCS
以TLCS進行排序,排名前5的高產作者也均來自于美國(表1),其中Ryba N. J. P.、Hoon M. A.和Chandrashekar J.來自加州大學圣迭戈分校,共同發表了Mammalian sweet taste receptors、T2Rs function as bitter taste receptors、A novel family of mammalian taste receptors等影響力較大的文獻,研究重點主要是哺乳動物甜味、鮮味和苦味的感知過程,排在第三名的霍華德·休斯醫學研究所的Zuker C. S.也參與了上述幾篇文獻的實驗研究。排名第一的作者Margolskee R. F.在闡述人類、鼠類的味覺感知機理以及診斷、治療相關疾病方面做出巨大貢獻,Margolskee R. F.研究了味覺蛋白T1R2、T1R3對甜味劑的響應和對胰島素分泌的調控機理,這對預防和治療吸收不良綜合征和包括糖尿病和肥胖癥在內的飲食相關疾病具有意義。總地來說,目前味覺受體領域的研究主要集中在美國的加州大學圣迭戈分校,Ryba N. J. P.、Hoon M. A.和Chandrashekar J.等科學家的研究處于領先位置。
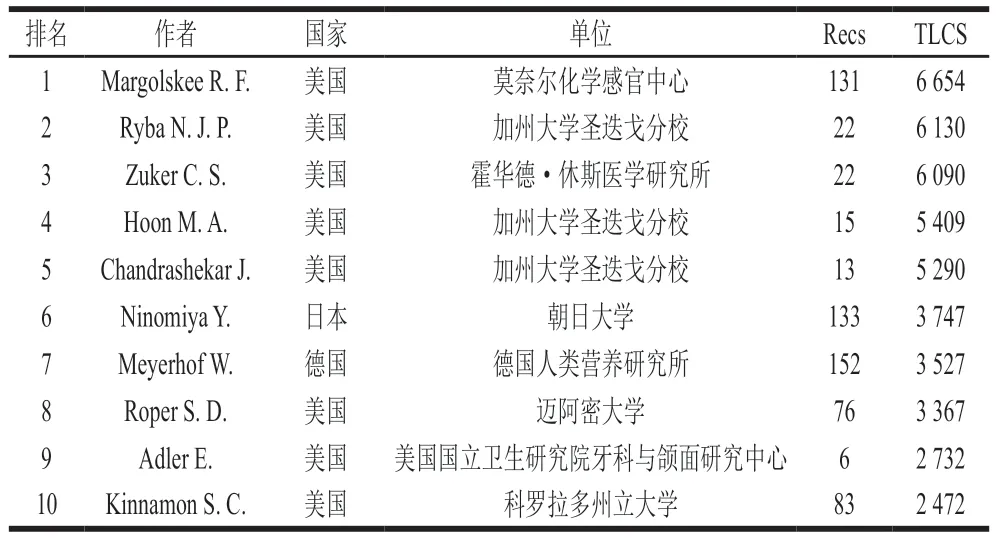
表1 Web of Science味覺受體領域文獻TLCS排序前10 名作者分布Table 1 Top 10 authors by TLCS of literature on taste receptors in Web of Science
6 結 語
味覺受體不僅存在于口腔,還遍布全身各處。口腔外的組織表達的味覺受體無法感知味覺,而是具有營養傳感、氣道免疫防御和代謝過程調節等作用,被認為是重要的藥物靶點,與呼吸道疾病、心腦血管疾病、糖尿病和肥胖等疾病相關,通過鼠類實驗模擬T2DM患者,發現黃芪多糖對T2DM癥狀有改善作用,黃酮則對治療上呼吸道感染疾病具有臨床潛力,因此開發用于治療疾病、減輕癥狀的小分子將成為一個非常活躍的研究領域。但由于獲得單晶困難,大多數受體的精確結構還未解析出,也難以區分不同特點的味覺神經刺激,目前主要采用計算機模擬(包括同源建模、分子對接等手段)構建受體模型,將現有模型與實驗數據進行比較,促進受體結構模型從低分辨率到高分辨率的過渡。雖然近年來我國在味覺受體尤其在生物傳感器和受體進化機制方面的研究取得很大的進展,但在味覺受體領域做出主要研究貢獻的國家還是美國,其中杰出的科學家包括加州大學圣迭戈分校的Ryba N. J. P.、Hoon M. A.和Chandrashekar J.等,我國需要與排在前列的國家、科研機構、科學家加大溝通交流。未來應深入加強味覺受體領域相互作用機制和信號傳導途徑方面的研究,攻克蛋白質表達純化和結晶方面的難題,探究進化和多態性的分子機制,同時通過將味覺受體和化學材料等多種學科結合增強傳感器穩定性。這些研究都有利于研制傳感器在模擬人類味覺、藥物篩選、研發新型增味劑和掩蔽劑、探索作為治療疾病的藥物靶點等方面的應用。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
