釀酒葡萄抗寒相關基因的研究進展
2022-10-04 05:04:02劉牧青孫雨航代玲敏
釀酒科技 2022年9期
關鍵詞:植物
馬 巖,劉牧青,孫雨航,鐘 軻,韓 寧,代玲敏
(齊魯工業大學(山東省科學院)生物工程學院,山東濟南 250353)
葡萄酒近年來在中國越來越被廣大人民接受,但葡萄酒的原料——歐洲葡萄品質雖好,但抗寒性較差。我國國土面積廣大,因氣候原因,北方地區冬季溫度過低導致葡萄植株受損而質量下滑。隨著氣候與土壤條件的不斷變化以及一些植物的跨地域栽培,植物對環境的適應變得極其重要。在一些極端條件下,植物為了適應環境不斷進化出一套防御體系。Jiang 等通過實驗發現,對低溫有一定抵抗能力的葡萄光合作用及與糖生物合成相關酶的活性比較強,這是植物開始適應低溫環境的一種本能反應;范高韜從歐亞種玫瑰香中分離得到一種基因并命名為,經低溫誘導及熒光定量PCR檢測后發現,該基因表達量在低溫誘導前期沒有顯著性變化,24 h 后顯著上調,這表明該基因可能被其他冷相關基因誘導表達。由此看出,基因的表達并不是獨立,而是一個甚至多個調控網絡。
隨著分子生物學等的不斷發展,利用分子生物學等方法探索在低溫條件下抗寒基因的表達途徑。第二信使分子在細胞內的信號傳遞過程中扮演著重要的角色,鈣離子與脫落酸(Abscisic Acid,ABA)比較常見。信號傳遞一般分為兩類,不依賴ABA途徑與依賴ABA途徑。
不依賴ABA 信號傳導途徑:在低溫條件下,植物細胞膜發生相變,這種刺激信號使Ca內流,細胞內的Ca受體蛋白激酶被磷酸化,CRLK 作為連接CRPK 與MAPK 級聯途徑的橋梁,低溫信號通過該途徑被傳遞并逐漸放大。Xu 等從山葡萄中克隆出兩種ICE 基因(、),當細胞感受到低溫信號后,在Ca信號作用下,基因編碼MYC 型轉錄因子并激活基因的表達,基因編碼bHLH 型轉錄因子以激活基因的表達,隨后轉錄因子與抗寒相關基因啟動子上的CRT/DRE 順式作用元件結合,啟動下游基因(、,基因相比于基因特異性調控基因)的表達。依賴ABA 信號傳導途徑:當細胞感受到低溫信號后,ABA 濃度升高,在ABA 受體PYL 上合成AREB 蛋白,該蛋白與CE 元件偶聯結合到啟動子片段的ABRE 順式作用元件上啟動下游基因的表達(見圖1)。
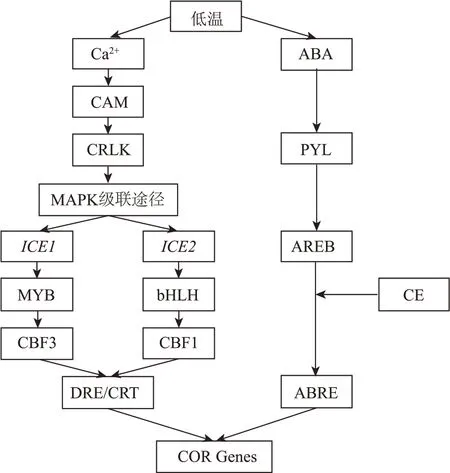
圖1 抗寒基因表達通路
低溫是限制植物生長發育及存活的逆境脅迫,不但會改變酶的生存條件,使酶活性降低甚至是喪失,還會改變細胞膜的狀態。當膜系統遭受冷凍傷害,為了適應低溫條件,細胞膜的不飽和脂肪酸比例升高,由于抗氧化酶活性受到影響,不飽和脂肪酸容易被氧化成脂氫過氧化物。脂氫過氧化物不僅會分解成丙二醛,還會均裂產生脂性自由基,這種物質既會不斷損傷細胞膜,又會氧化蛋白質從而使蛋白質發生聚集。低溫還會降低與光合作用相關的酶類活性:1,5-二磷酸核酮糖羧化酶/加氧酶(Rubisco)、磷酸核酮糖激酶(PRK)、3-磷酸甘油醛脫氫酶(GAPDH)等,減少糖類的合成。王依等[選取4種釀酒葡萄以篩選低溫適應性強的葡萄品種,低溫處理后發現過氧化氫酶活性以及可溶性脂肪含量升高,脯氨酸、可溶性糖及超氧化物歧化酶活性先升高后下降,過氧化物酶活性降低,電導率及丙二醛含量升高。2018年,賀蘭山地區發生低溫災害,導致90%一年生葡萄幼株、30%~40%兩年生葡萄幼株受凍死亡。低溫條件會觸發植物自身的抵抗機制來抵御脅迫,但仍會對其造成極大的損害。因此我們需要采取并對比一切可以提高葡萄抗寒性的合理措施以使葡萄安然越冬。
1 提高葡萄抗寒性的措施
提高葡萄越冬能力的方法多種多樣:(1)埋土。埋土作為廣泛采用的能使葡萄安然越冬的方法,從結果來看是極其有效的。葡萄在越冬期間,一年生枝干以及地表附近的根系容易受到損傷,土埋增加了根系周圍的土壤厚度,可有效提高根系周圍溫度,保護根系免受損傷,在來年可使其有效吸收水分及營養。但其存在的問題也比較明顯:首先,土埋比較費時費力,取土也存在一定風險,只從葡萄株周圍取而不回填可能會人為對根系造成損傷,取土會造成取土區土壤厚度下降,在冬季對其造成二次傷害;其次,在葡萄成熟期以及采摘期,如果操作不得當,還會使腐爛的葡萄顆粒留在土壤中使其感染病害。(2)選擇抗寒的砧木嫁接。不同砧木的抗寒性、抗旱性、抗鹽堿能力及抗病性存在差異,需要根據葡萄品種的特性選擇合適砧木。對5種抗寒性較好的砧木(5BB、5C、SO4、140R、kangzhen3)綜合抗寒力進行比較,發現5BB 抗寒性最高。(3)灌水和熏煙。該方法需要及時關注天氣情況,在霜凍來臨前5 h 灌水,或點燃碎木屑,利用燃燒產生的熱量增加株間溫度。(4)建設保溫棚。胡悅等發現保溫棚可以有效提高棚內地下20 cm 以內的溫度,空氣濕度、土壤濕度都能有效提高,但是大棚的建設成本比較高,費時費力,適合小規模使用。(5)建設防護林。在冬季風向的垂直方向建造防護林,使冷空氣沿樹體爬升。(6)噴施殼聚糖。殼聚糖分解后產生低聚合度的堿性寡糖可引起葡萄植株的應激反應,增加植株內滲透調節物的含量,降低冰點,提高抗寒性。(7)選擇合適的葡萄園,盡可能避開冷空氣聚集的低洼區。(8)通過基因工程的手段,定向提高葡萄的抗低溫能力。目前主要是通過轉入調控基因或者目的基因來改善葡萄的抗寒性。因為基因的插入位點隨機,需要篩選抗性優良的轉基因植株以穩定遺傳。目前發現低溫轉錄因子(、等)、脫水素家族、單鋅指結合蛋白家族和泛素/26S 蛋白酶體系的表達會顯著提高植物的抗寒性。外源基因在植物的有性生殖中具有遺傳穩定性,可避免每年投入大量的人力物力。從長遠角度考慮,利用分子生物學及遺傳學方法與技術定向提高葡萄的抗寒性具有很光明的前景。
2 抗寒相關基因
2.1 抗寒相關轉錄因子
CBF 轉錄因子屬于AP2/EREBP 型轉錄因子,是植物低溫脅迫中主要依賴的一條抗寒途徑。在擬南芥中發現的CBF1、CBF2、CBF3 和CBF4 基因,前三者位于四號染色上,后者位于五號染色體上且被發現是四類CBF 基因中唯一一個與ABA 調控途徑互作的基因。當植物遭受低溫脅迫,ICE(inducer of CBF expression)基因被誘導表達,該基因結合到CBF 基因的啟動子區域直接驅動CBF 基因的表達。CBF 蛋白隨后結合到COR 基因啟動子的CRT/DRE 順式作用元件上啟動下游基因的表達。從山葡萄中克隆得到VaCBF1 基因,經脅迫處理及熒光定量PCR 檢測后發現,該基因受低溫誘導表達,不受山梨醇和氯化鈉誘導。其他轉錄因子,如、、、及等都可以提高植物抗寒性。
2.2 抗寒功能基因
2.2.1 脫水素基因
脫水素(Dehydrin,DHN)是一種晚期胚胎發育豐富蛋白(late embryogenesis abundant protein,LEA),屬于LEA 蛋白的第二家族,分子量在9~200 KDa 之間,在種子發育后期和逆境條件下大量積累。Malik發現在非生物脅迫下,脫水素基因在轉錄和翻譯水平均上調且植物的抗逆性提高。脫水素由3 個保守片段:K 片段、S 片段和Y 片段組成,Mota還發現富含極性氨基酸的、不保守的φ片段參與形成SKn 型大豆脫水素。K 片段位于C末端,可形成雙親性的α-螺旋結構,K 片段與陽離子抗菌多肽性質極度相似,脫水素表達會抑制大腸桿菌的生長。S 片段由一系列絲氨酸組成,易發生磷酸化,與核定位有關的信號肽結合,進入細胞核,與脫水素定位到細胞核并幫助肽鏈初步折疊行使分子伴侶功能有關。還有一些S 片段包含LHR 序列(GXGGRRKK),能夠與K 片段連接,也可發生磷酸化。Y 片段((V/T)DEYGNP)位于N 末端,其氨基酸序列與植物和細菌的分子伴侶的核酸結合位點的氨基酸序列具有同源性。Y 片段富含甘氨酸,與組織特異性有關,亞細胞定位發現含Y 片段的脫水素在種子中大量表達,其他不含Y片段的脫水素沒有組織特異性。脫水素含有的3個保守片段數量和排列方式被分為五類,不同類型脫水素響應的脅迫也存在區別(見表1)。

表1 不同類型脫水素的性質與響應模式
其中,YSK型脫水素以其雙親性的特點,通過靜電作用與細胞膜的脂質分子結合,在細胞失水條件下可以防止膜聚集而保護膜結構。KS 型脫水素可以直接結合金屬離子或間接清除活性氧自由基,降低氧化損害。SK和K型脫水素在低溫脅迫下有明顯積累。脫水素最早在研究棉花子葉中的轉運蛋白時被發現,這類蛋白因在逆境脅迫下大量累積被廣泛關注。脫水素的功能主要體現在以下幾個方面:(1)K 片段賦予脫水素的雙親性結合膜結構,防止膜發生聚集;(2)結合細胞內的金屬離子維持細胞內抗氧化酶的活性,降低由金屬離子產生的活性氧自由基;(3)具有分子伴侶的功能。脫水素會在植物中選擇性表達避免浪費機體資源,如亞洲白樺樹為適應凍害在冬季表達大量脫水素蛋白但在夏季不會表達。第一次表明脫水素具有抗凍活性的研究是從桃中分離到的PAC60 基因,它既可以防止細胞內冰晶形成,又可以產生熱滯效應,使植物在一定程度上免受冰晶造成的物理損傷,提高植物的耐低溫性。過表達榴蓮脫水素基因的擬南芥經低溫處理,發現脫水素可以提高轉基因擬南芥的抗寒性。將梅花脫水素基因轉入煙草后發現轉基因煙草的葉片相對含水量、可溶性糖含量都明顯升高,抗寒性也隨之升高。Hincha認為脫水素對一些關鍵酶產生的保護作用與它本質上無規則的結構密切相關,脫水素維持酶的活性有兩個可能:(1)依賴其不規則的結構形成物理障礙,阻礙蛋白質的聚集變性;(2)脫水素可以防止蛋白質錯誤折疊從而維持酶活性。Sánchez 等從仙人掌中分離得到一個SK3 型脫水素基因,發現該基因的表達產物在植物體內可以自身相互作用,K 片段與富含組氨酸的基序在自身互作中起著關鍵性作用。脫水素分子量大小與其對乳酸脫氫酶的保護能力呈正相關。葡萄中的脫水素基因比較少,共分離出4種 并 命 名 為、、、,其 中受低溫、高溫、干旱和白粉病誘導表達,受高溫、低溫誘導表達,后兩者僅在胚胎發育后期的種子中表達。
2.2.2 DNA結合單鋅指蛋白基因
DNA 結合單鋅指(DNA binding with one zinc finger,Dof)蛋白是植物中特有的一類蛋白,單鋅指結構由4個半胱氨酸與一個鋅離子結合組成。
該蛋白一般由200~400個氨基酸組成,N末端是52 個氨基酸組成的具有保守性的功能結合結構域,可結合DNA 及蛋白質以相互作用;C 末端含有不保守的轉錄調控域,賦予Dof 蛋白多樣性。AAAG 基序是Dof 蛋白結合域的核心結合位點,Dof 蛋白還會與細胞內的可與DNA 結合的未磷酸化的HMG 蛋白互作,增加Dof 蛋白的結合能力。Dof 蛋白還參與植物中的種子萌發、碳氮代謝、開花調控及光調節等。Zhang 等從蘋果中分離出60 個Dof 蛋白基因,大多數響應低溫脅迫。Dof 蛋白是一種多功能轉錄因子,Su 等發現,GhDof 在低溫條件下可被誘導表達,并提高細胞內、、基因的表達及種子含油量;Ma等發現白菜基因在低溫條件下被上調;李成慧在葡萄當中鑒定出25 個基因,其中11 個基因參與響應生物和非生物脅迫;代偉娜發現基因響應激素誘導,對MeJA 響應最為明顯,MeJA 通常與植物抵御病蟲害等生物脅迫息息相關。
2.2.3 泛素/26S蛋白酶體基因
泛素是一類由76 個氨基酸組成的高度保守的小分子蛋白質。N 端的1 個甲硫氨酸及7 個賴氨酸可以與其他泛素分子結合形成泛素鏈以調節生物體不同的生命活動。泛素/26S 蛋白酶體系是真核生物體內降解蛋白的有效途徑,可降解生物體內80%~85%的蛋白質。該體系包含泛素、泛素激活酶(E1)、泛素結合酶(E2)、泛素連接酶(E3)、蛋白酶體。泛素在有ATP 供應下被E1 激活,隨即與E2 結合形成E2-泛素結合體,再與E3 連接并將泛素直接或間接轉移到蛋白上,使蛋白泛素化以被蛋白酶體識別并降解。泛素連接酶可被分為四類:U-box、HECT、RING 和F-box,前三者屬于單亞基類,后者屬于多亞基類。U-box 與RING 相似度較高,二者較明顯的區別在于U-box 類連接酶靠鹽橋和氫鍵維持結構的穩定性,RING 類連接酶靠半胱氨酸和鋅離子構成的鋅指結構維持穩定性。Ubox 擁有4 類結構域:ARM、Ser/Thr kinase、WD40和MIF4G。ARM 位于U-box 類型泛素連接酶的C末端,可與其他蛋白互作。Yao 等發現中國野生華東葡萄泛素連接酶基因經低溫誘導后在葡萄原生質體中大量表達,通過酵母雙雜技術發現,與互作,低溫條件下促進VpICE1 蛋白積累,抑制VpHOS1 蛋白對VpICE1 蛋白的降解作用。在過表達的擬南芥中,低溫處理后經RT-PCR 檢測發現不止該基因上調,與低溫脅迫有關的、、等基因也被上調。另一種U-box 類E3 泛素連接酶從左山一號被分離出并被命名為,該基因在低溫條件下在各個組織中大量表達,在擬南芥中過表達該基因后,發現與低溫相關的/、/及等冷相關基因被上調,但/基因被下調,該現象說明響應低溫脅迫的同時也會影響其他冷相關基因的表達。
3 總結與展望
提高歐洲葡萄抗寒性的方法多種多樣,具體可劃分為兩類:(1)改變葡萄生存的外部環境;(2)研究抗寒相關基因,并通過基因工程的手段改變葡萄自身以適應不利生存條件。本文闡述重要抗寒調控基因和抗寒功能基因的功能及其對植物抗寒性的影響,為研究葡萄抗寒基因并定向改良葡萄抗寒性提供參考。ICEs 類轉錄因子在遭受低溫脅迫時率先表達,作為一種“誘導劑”誘導CBFs 類轉錄因子的表達,最終該類型轉錄因子結合到抗寒相關基因啟動子區域的順式作用元件CRT/DRE 上啟動下游基因表達。脫水素基因表達的蛋白主要對細胞內的膜結構起到保護作用,起到分子伴侶的功能,保護細胞內關鍵酶的活性,保護能力與脫水素的大小呈正相關關系;單鋅指蛋白在植物遭受低溫脅迫時,通過調節光合作用合成生命活動所必需的物質;泛素/26S 蛋白酶體則通過降解蛋白以維持細胞內與抗寒相關蛋白的含量,提高植物抗寒性。三個類型的基因在低溫條件下都會引起其他大部分冷相關基因上調及小部分基因下調,說明基因的表達受到一系列小分子/大分子物質調控。目前國內外還存在抗寒基因及表達方式認知不足的問題,因此我們還需不斷挖掘抗寒基因,探索它們的表達方式以緩解低溫給植物生長造成的壓力。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
